
Mediterranean Diet and Neurodegenerative Diseases: The Neglected Role of Nutrition in the Modulation of the Endocannabinoid System

 , and
, and
Abstract
:1. Endocannabinoid System: Physiology and Pathophysiology No
2. Mediterranean Diet
3. CB2R Ligands: β-Caryophyllene and 3,3′-Diindolylmethane
4. FAAH Inhibitors: Flavonoids
5. Dietary ω-3 and ω-6 Fatty Acids Balance and Endocannabinoid System Coupling
6. Between “Entourage” Effect and Noncanonical CBRs: N-Acylethanolamines (NAEs) and Resveratrol
7. EC Activity and Diseases
8. EC Induction of Obesity Counteracted by MedD-Related Compounds
9. ECs and Gut Microbiome
10. ECs and Inflammation
11. ECs and Cancer
12. Endocannabinoid Tone Modulation by Plant Flavonoids
13. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moreno, E.; Cavic, M.; Canela, E.I. Functional Fine-Tuning of Metabolic Pathways by the Endocannabinoid System—Implications for Health and Disease. Int. J. Mol. Sci. 2021, 22, 3661. [Google Scholar] [CrossRef] [PubMed]
- Simon, V.; Cota, D. Mechanisms in Endocrinology: Endocannabinoids and Metabolism: Past, Present and Future. Eur. J. Endocrinol. 2017, 176, R309–R324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J. Cannabis, Cannabinoid Receptors, and Endocannabinoid System: Yesterday, Today, and Tomorrow. Acta Pharm. Sin. 2019, 40, 297–299. [Google Scholar] [CrossRef] [PubMed]
- Gowran, A.; Noonan, J.; Campbell, V.A. The Multiplicity of Action of Cannabinoids: Implications for Treating Neurodegeneration. CNS Neurosci. Ther. 2011, 17, 637–644. [Google Scholar] [CrossRef]
- McPartland, J.M.; Guy, G.W.; Di Marzo, V. Care and Feeding of the Endocannabinoid System: A Systematic Review of Potential Clinical Interventions That Upregulate the Endocannabinoid System. PLoS ONE 2014, 9, e89566. [Google Scholar] [CrossRef]
- Pagotto, U.; Marsicano, G.; Cota, D.; Lutz, B.; Pasquali, R. The Emerging Role of the Endocannabinoid System in Endocrine Regulation and Energy Balance. Endocr. Rev. 2006, 27, 73–100. [Google Scholar] [CrossRef]
- Silvestri, C.; Di Marzo, V. The Endocannabinoid System in Energy Homeostasis and the Etiopathology of Metabolic Disorders. Cell Metab. 2013, 17, 475–490. [Google Scholar] [CrossRef] [Green Version]
- Harrold, J.A.; Williams, G. The Cannabinoid System: A Role in Both the Homeostatic and Hedonic Control of Eating? Br. J. Nutr. 2003, 90, 729–734. [Google Scholar] [CrossRef] [Green Version]
- Vendel, E.; de Lange, E.C.M. Functions of the CB1 and CB2 Receptors in Neuroprotection at the Level of the Blood–Brain Barrier. Neuromol. Med. 2014, 16, 620–642. [Google Scholar] [CrossRef]
- Svizenska, I.; Dubovy, P.; Sulcova, A. Cannabinoid Receptors 1 and 2 (CB1 and CB2), Their Distribution, Ligands and Functional Involvement in Nervous System Structures—A Short Review. Pharm. Biochem. Behav. 2008, 90, 501–511. [Google Scholar] [CrossRef]
- Berdyshev, E.V. Cannabinoid Receptors and the Regulation of Immune Response. Chem. Phys. Lipids 2000, 108, 169–190. [Google Scholar] [CrossRef]
- Sharkey, K.A.; Wiley, J.W. The Role of the Endocannabinoid System in the Brain–Gut Axis. Gastroenterology 2016, 151, 252–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagan, S.G.; Campbell, V.A. The Influence of Cannabinoids on Generic Traits of Neurodegeneration: Cannabinoids and Neurodegeneration. Br. J. Pharmcol. 2014, 171, 1347–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cristino, L.; Bisogno, T.; Di Marzo, V. Cannabinoids and the Expanded Endocannabinoid System in Neurological Disorders. Nat. Rev. Neurol. 2020, 16, 9–29. [Google Scholar] [CrossRef] [PubMed]
- Grygiel-Górniak, B. Peroxisome Proliferator-Activated Receptors and Their Ligands: Nutritional and Clinical Implications—A Review. Nutr. J. 2014, 13, 17. [Google Scholar] [CrossRef] [Green Version]
- Laleh, P.; Yaser, K.; Abolfazl, B.; Shahriar, A.; Mohammad, A.J.; Nazila, F.; Alireza, O. Oleoylethanolamide Increases the Expression of PPAR-A and Reduces Appetite and Body Weight in Obese People: A Clinical Trial. Appetite 2018, 128, 44–49. [Google Scholar] [CrossRef]
- Luchicchi, A.; Pistis, M. Anandamide and 2-Arachidonoylglycerol: Pharmacological Properties, Functional Features, and Emerging Specificities of the Two Major Endocannabinoids. Mol. Neurobiol. 2012, 46, 374–392. [Google Scholar] [CrossRef] [PubMed]
- Fezza, F.; Bari, M.; Florio, R.; Talamonti, E.; Feole, M.; Maccarrone, M. Endocannabinoids, Related Compounds and Their Metabolic Routes. Molecules 2014, 19, 17078–17106. [Google Scholar] [CrossRef]
- Fernández-Ruiz, J.; Romero, J.; Ramos, J.A. Endocannabinoids and Neurodegenerative Disorders: Parkinson’s Disease, Huntington’s Chorea, Alzheimer’s Disease, and Others. In Endocannabinoids; Pertwee, R.G., Ed.; Handbook of Experimental Pharmacology; Springer International Publishing: Cham, Switzerland, 2015; Volume 231, pp. 233–259. ISBN 978-3-319-20824-4. [Google Scholar]
- Basavarajappa, B.S.; Shivakumar, M.; Joshi, V.; Subbanna, S. Endocannabinoid System in Neurodegenerative Disorders. J. Neurochem. 2017, 142, 624–648. [Google Scholar] [CrossRef]
- Chitnis, T.; Weiner, H.L. CNS Inflammation and Neurodegeneration. J. Clin. Investig. 2017, 127, 3577–3587. [Google Scholar] [CrossRef] [Green Version]
- Scuderi, C.; Steardo, L. Neuroglial Roots of Neurodegenerative Diseases: Therapeutic Potential of Palmitoylethanolamide in Models of Alzheimer’s Disease. CNS Neurol. Disord. Drug Targets 2013, 12, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Köfalvi, A.; Lemos, C.; Martín-Moreno, A.M.; Pinheiro, B.S.; García-García, L.; Pozo, M.A.; Valério-Fernandes, Â.; Beleza, R.O.; Agostinho, P.; Rodrigues, R.J.; et al. Stimulation of Brain Glucose Uptake by Cannabinoid CB2 Receptors and Its Therapeutic Potential in Alzheimer’s Disease. Neuropharmacology 2016, 110, 519–529. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.; Zhang, J.; Wu, Y.; Wang, D.; Feng, G.; Tang, Y.-P.; Teng, Z.; Chen, C. Monoacylglycerol Lipase Is a Therapeutic Target for Alzheimer’s Disease. Cell Rep. 2012, 2, 1329–1339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, S.; Wang, Z.; Zhang, Y.; Chen, N. Potential Application of Endocannabinoid System Agents in Neuropsychiatric and Neurodegenerative Diseases—Focusing on FAAH/MAGL Inhibitors. Acta Pharm. Sin. 2020, 41, 1263–1271. [Google Scholar] [CrossRef] [PubMed]
- D’Addario, C.; Di Francesco, A.; Arosio, B.; Gussago, C.; Dell’Osso, B.; Bari, M.; Galimberti, D.; Scarpini, E.; Altamura, A.C.; Mari, D.; et al. Epigenetic Regulation of Fatty Acid Amide Hydrolase in Alzheimer Disease. PLoS ONE 2012, 7, e39186. [Google Scholar] [CrossRef] [Green Version]
- Aso, E.; Juvés, S.; Maldonado, R.; Ferrer, I. CB2 Cannabinoid Receptor Agonist Ameliorates Alzheimer-Like Phenotype in AβPP/PS1 Mice. J. Alzheimer’s Dis. 2013, 35, 847–858. [Google Scholar] [CrossRef] [Green Version]
- Koppel, J.; Vingtdeux, V.; Marambaud, P.; d’Abramo, C.; Jimenez, H.; Stauber, M.; Friedman, R.; Davies, P. CB2 Receptor Deficiency Increases Amyloid Pathology and Alters Tau Processing in a Transgenic Mouse Model of Alzheimer’s Disease. Mol. Med. 2013, 19, 29–36. [Google Scholar] [CrossRef]
- García-Arencibia, M.; González, S.; de Lago, E.; Ramos, J.A.; Mechoulam, R.; Fernández-Ruiz, J. Evaluation of the Neuroprotective Effect of Cannabinoids in a Rat Model of Parkinson’s Disease: Importance of Antioxidant and Cannabinoid Receptor-Independent Properties. Brain Res. 2007, 1134, 162–170. [Google Scholar] [CrossRef]
- Shi, J.; Cai, Q.; Zhang, J.; He, X.; Liu, Y.; Zhu, R.; Jin, L. AM1241 Alleviates MPTP-Induced Parkinson’s Disease and Promotes the Regeneration of DA Neurons in PD Mice. Oncotarget 2017, 8, 67837–67850. [Google Scholar] [CrossRef] [Green Version]
- Chung, Y.C.; Shin, W.-H.; Baek, J.Y.; Cho, E.J.; Baik, H.H.; Kim, S.R.; Won, S.-Y.; Jin, B.K. CB2 Receptor Activation Prevents Glial-Derived Neurotoxic Mediator Production, BBB Leakage and Peripheral Immune Cell Infiltration and Rescues Dopamine Neurons in the MPTP Model of Parkinson’s Disease. Exp. Mol. Med. 2016, 48, e205. [Google Scholar] [CrossRef]
- Gómez-Gálvez, Y.; Palomo-Garo, C.; Fernández-Ruiz, J.; García, C. Potential of the Cannabinoid CB2 Receptor as a Pharmacological Target against Inflammation in Parkinson’s Disease. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2016, 64, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Suárez, D.; Celorrio, M.; Riezu-Boj, J.I.; Ugarte, A.; Pacheco, R.; González, H.; Oyarzabal, J.; Hillard, C.J.; Franco, R.; Aymerich, M.S. The Monoacylglycerol Lipase Inhibitor JZL184 Is Neuroprotective and Alters Glial Cell Phenotype in the Chronic MPTP Mouse Model. Neurobiol. Aging 2014, 35, 2603–2616. [Google Scholar] [CrossRef] [PubMed]
- Celorrio, M.; Fernández-Suárez, D.; Rojo-Bustamante, E.; Echeverry-Alzate, V.; Ramírez, M.J.; Hillard, C.J.; López-Moreno, J.A.; Maldonado, R.; Oyarzábal, J.; Franco, R.; et al. Fatty Acid Amide Hydrolase Inhibition for the Symptomatic Relief of Parkinson’s Disease. Brain Behav. Immun. 2016, 57, 94–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viveros-Paredes, J.M.; Gonzalez-Castañeda, R.E.; Escalante-Castañeda, A.; Tejeda-Martínez, A.R.; Castañeda-Achutiguí, F.; Flores-Soto, M.E. Effect of inhibition of fatty acid amide hydrolase on MPTP-induced dopaminergic neuronal damage. Neurología 2019, 34, 143–152. [Google Scholar] [CrossRef]
- Di Marzo, V.; Hill, M.P.; Crossman, V.; Brotchie, M. Enhanced Levels of Endogenous Cannabinoids in the Globus Pallidus Are Associated with a Reduction in Movement in an Animal Model of Parkinson’s Disease. FASEB J. 2000, 14, 1432–1438. [Google Scholar] [CrossRef] [PubMed]
- Pisani, A.; Fezza, F.; Galati, S.; Battista, N.; Napolitano, S.; Finazzi-Agrò, A.; Bernardi, G.; Brusa, L.; Pierantozzi, M.; Stanzione, P.; et al. High Endogenous Cannabinoid Levels in the Cerebrospinal Fluid of Untreated Parkinson’s Disease Patients. Ann. Neurol. 2005, 57, 777–779. [Google Scholar] [CrossRef] [PubMed]
- Van Laere, K.; Casteels, C.; Lunskens, S.; Goffin, K.; Grachev, I.D.; Bormans, G.; Vandenberghe, W. Regional Changes in Type 1 Cannabinoid Receptor Availability in Parkinson’s Disease in Vivo. Neurobiol. Aging 2012, 33, 620.e1–620.e8. [Google Scholar] [CrossRef]
- Arevalo-Martin, A.; Molina-Holgado, E.; Guaza, C. A CB1/CB2 Receptor Agonist, WIN 55,212–2, Exerts Its Therapeutic Effect in a Viral Autoimmune Model of Multiple Sclerosis by Restoring Self-Tolerance to Myelin. Neuropharmacology 2012, 63, 385–393. [Google Scholar] [CrossRef] [Green Version]
- Baker, D.; Pryce, G.; Croxford, J.L.; Brown, P.; Pertwee, R.G.; Huffman, J.W.; Layward, L. Cannabinoids Control Spasticity and Tremor in a Multiple Sclerosis Model. Nature 2000, 404, 84–87. [Google Scholar] [CrossRef]
- Benito, C.; Romero, J.P.; Tolon, R.M.; Clemente, D.; Docagne, F.; Hillard, C.J.; Guaza, C.; Romero, J. Cannabinoid CB1 and CB2 Receptors and Fatty Acid Amide Hydrolase Are Specific Markers of Plaque Cell Subtypes in Human Multiple Sclerosis. J. Neurosci. 2007, 27, 2396–2402. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, S.E. Cannabinoids Go Nuclear: Evidence for Activation of Peroxisome Proliferator-Activated Receptors: Cannabinoids and PPARs. Br. J. Pharmcol. 2007, 152, 576–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malamas, M.S.; Farah, S.I.; Lamani, M.; Pelekoudas, D.N.; Perry, N.T.; Rajarshi, G.; Miyabe, C.Y.; Chandrashekhar, H.; West, J.; Pavlopoulos, S.; et al. Design and Synthesis of Cyanamides as Potent and Selective N-Acylethanolamine Acid Amidase Inhibitors. Bioorg. Med. Chem. 2020, 28, 115195. [Google Scholar] [CrossRef] [PubMed]
- Alhouayek, M.; Muccioli, G.G. Harnessing the Anti-Inflammatory Potential of Palmitoylethanolamide. Drug Discov. Today 2014, 19, 1632–1639. [Google Scholar] [CrossRef] [PubMed]
- Hansen, H.S.; Diep, T.A. N-Acylethanolamines, Anandamide and Food Intake. Biochem. Pharm. 2009, 78, 553–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrosino, S.; Di Marzo, V. The Pharmacology of Palmitoylethanolamide and First Data on the Therapeutic Efficacy of Some of Its New Formulations: Palmitoylethanolamide and Its New Formulations. Br. J. Pharmcol. 2017, 174, 1349–1365. [Google Scholar] [CrossRef]
- Casili, G.; Lanza, M.; Campolo, M.; Siracusa, R.; Paterniti, I.; Ardizzone, A.; Scuderi, S.A.; Cuzzocrea, S.; Esposito, E. Synergic Therapeutic Potential of PEA-Um Treatment and NAAA Enzyme Silencing In the Management of Neuroinflammation. Int. J. Mol. Sci. 2020, 21, 7486. [Google Scholar] [CrossRef] [PubMed]
- Gardener, H.; Caunca, M.R. Mediterranean Diet in Preventing Neurodegenerative Diseases. Curr. Nutr. Rep. 2018, 7, 10–20. [Google Scholar] [CrossRef]
- Gardener, H.; Wright, C.B.; Rundek, T.; Sacco, R.L. Brain Health and Shared Risk Factors for Dementia and Stroke. Nat. Rev. Neurol. 2015, 11, 651–657. [Google Scholar] [CrossRef]
- McGrattan, A.M.; McGuinness, B.; McKinley, M.C.; Kee, F.; Passmore, P.; Woodside, J.V.; McEvoy, C.T. Diet and Inflammation in Cognitive Ageing and Alzheimer’s Disease. Curr. Nutr. Rep. 2019, 8, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Ozawa, M.; Shipley, M.; Kivimaki, M.; Singh-Manoux, A.; Brunner, E.J. Dietary Pattern, Inflammation and Cognitive Decline: The Whitehall II Prospective Cohort Study. Clin. Nutr. 2017, 36, 506–512. [Google Scholar] [CrossRef] [Green Version]
- Tsivgoulis, G.; Judd, S.; Letter, A.J.; Alexandrov, A.V.; Howard, G.; Nahab, F.; Unverzagt, F.W.; Moy, C.; Howard, V.J.; Kissela, B.; et al. Adherence to a Mediterranean Diet and Risk of Incident Cognitive Impairment. Neurology 2013, 80, 1684–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarmeas, N.; Stern, Y.; Tang, M.-X.; Mayeux, R.; Luchsinger, J.A. Mediterranean Diet and Risk for Alzheimer’s Disease. Ann. Neurol. 2006, 59, 912–921. [Google Scholar] [CrossRef] [Green Version]
- Féart, C.; Samieri,, C.; Rondeau, V.; Portet, F.; Dartigues, J.-F.; Scarmeas, N.; Barberger-Gateau, P. Adherence to a Mediterranean Diet, Cognitive Decline, and Risk of Dementia. JAMA 2009, 302, 638–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tangney, C.C.; Kwasny, M.J.; Li, H.; Wilson, R.S.; Evans, D.A.; Morris, M.C. Adherence to a Mediterranean-Type Dietary Pattern and Cognitive Decline in a Community Population. Am. J. Clin. Nutr. 2011, 93, 601–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwingshackl, L.; Morze, J.; Hoffmann, G. Mediterranean Diet and Health Status: Active Ingredients and Pharmacological Mechanisms. Br. J. Pharmcol. 2020, 177, 1241–1257. [Google Scholar] [CrossRef] [Green Version]
- Davis, C.; Bryan, J.; Hodgson, J.; Murphy, K. Definition of the Mediterranean Diet; A Literature Review. Nutrients 2015, 7, 9139–9153. [Google Scholar] [CrossRef]
- Serra-Majem, L.; Tomaino, L.; Dernini, S.; Berry, E.M.; Lairon, D.; Ngo de la Cruz, J.; Bach-Faig, A.; Donini, L.M.; Medina, F.-X.; Belahsen, R.; et al. Updating the Mediterranean Diet Pyramid towards Sustainability: Focus on Environmental Concerns. Int. J. Env. Res. Public Health 2020, 17, 8758. [Google Scholar] [CrossRef]
- Abumweis, S.S.; Barake, R.; Jones, P. Plant Sterols/Stanols as Cholesterol Lowering Agents A Meta-Analysis of Randomized Controlled Trials. Food Nutr. Res. 2008, 52, 1811. [Google Scholar] [CrossRef]
- Salas-Salvadó, J.; Fernández-Ballart, J.; Ros, E.; Martínez-González, M.-A.; Fitó, M.; Estruch, R.; Corella, D.; Fiol, M.; Gómez-Gracia, E.; Arós, F.; et al. Effect of a Mediterranean Diet Supplemented With Nuts on Metabolic Syndrome Status: One-Year Results of the PREDIMED Randomized Trial. Arch. Intern. Med. 2008, 168, 2449. [Google Scholar] [CrossRef] [Green Version]
- Pusztai, A.; Grant, G. Assessment of Lectin Inactivation by Heat and Digestion. Methods Mol. Med. 1998, 9, 505–514. [Google Scholar] [CrossRef]
- De Punder, K.; Pruimboom, L. The Dietary Intake of Wheat and Other Cereal Grains and Their Role in Inflammation. Nutrients 2013, 5, 771–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petroski, W.; Minich, D.M. Is There Such a Thing as “Anti-Nutrients”? A Narrative Review of Perceived Problematic Plant Compounds. Nutrients 2020, 12, 2929. [Google Scholar] [CrossRef] [PubMed]
- Van Buul, V.J.; Brouns, F.J.P.H. Health Effects of Wheat Lectins: A Review. J. Cereal Sci. 2014, 59, 112–117. [Google Scholar] [CrossRef]
- Razquin, C.; Martinez-Gonzalez, M.A. A Traditional Mediterranean Diet Effectively Reduces Inflammation and Improves Cardiovascular Health. Nutrients 2019, 11, 1842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsigalou, C.; Konstantinidis, T.; Paraschaki, A.; Stavropoulou, E.; Voidarou, C.; Bezirtzoglou, E. Mediterranean Diet as a Tool to Combat Inflammation and Chronic Diseases. An Overview. Biomedicines 2020, 8, 201. [Google Scholar] [CrossRef] [PubMed]
- Bonaccio, M.; Pounis, G.; Cerletti, C.; Donati, M.B.; Iacoviello, L.; de Gaetano, G. MOLI-SANI Study Investigators Mediterranean Diet, Dietary Polyphenols and Low Grade Inflammation: Results from the MOLI-SANI Study. Br. J. Clin. Pharm. 2017, 83, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Schwingshackl, L.; Hoffmann, G. Mediterranean Dietary Pattern, Inflammation and Endothelial Function: A Systematic Review and Meta-Analysis of Intervention Trials. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 929–939. [Google Scholar] [CrossRef]
- Bailey, M.A.; Holscher, H.D. Microbiome-Mediated Effects of the Mediterranean Diet on Inflammation. Adv. Nutr. 2018, 9, 193–206. [Google Scholar] [CrossRef] [Green Version]
- Machowetz, A.; Poulsen, H.E.; Gruendel, S.; Weimann, A.; Fitó, M.; Marrugat, J.; Torre, R.; Salonen, J.T.; Nyyssönen, K.; Mursu, J.; et al. Effect of Olive Oils on Biomarkers of Oxidative DNA Stress in Northern and Southern Europeans. FASEB J. 2007, 21, 45–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annuzzi, G.; Bozzetto, L.; Costabile, G.; Giacco, R.; Mangione, A.; Anniballi, G.; Vitale, M.; Vetrani, C.; Cipriano, P.; Corte, G.D.; et al. Diets Naturally Rich in Polyphenols Improve Fasting and Postprandial Dyslipidemia and Reduce Oxidative Stress: A Randomized Controlled Trial. Am. J. Clin. Nutr. 2014, 99, 463–471. [Google Scholar] [CrossRef] [Green Version]
- Camargo, A.; Ruano, J.; Fernandez, J.M.; Parnell, L.D.; Jimenez, A.; Santos-Gonzalez, M.; Marin, C.; Perez-Martinez, P.; Uceda, M.; Lopez-Miranda, J.; et al. Gene Expression Changes in Mononuclear Cells in Patients with Metabolic Syndrome after Acute Intake of Phenol-Rich Virgin Olive Oil. BMC Genom. 2010, 11, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zern, T.L.; Fernandez, M.L. Cardioprotective Effects of Dietary Polyphenols. J. Nutr. 2005, 135, 2291–2294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malaguarnera Influence of Resveratrol on the Immune Response. Nutrients 2019, 11, 946. [CrossRef] [PubMed] [Green Version]
- Esposito, S.; Sparaco, M.; Maniscalco, G.T.; Signoriello, E.; Lanzillo, R.; Russo, C.; Carmisciano, L.; Cepparulo, S.; Lavorgna, L.; Gallo, A.; et al. Lifestyle and Mediterranean Diet Adherence in a Cohort of Southern Italian Patients with Multiple Sclerosis. Mult. Scler. Relat. Disord. 2021, 47, 102636. [Google Scholar] [CrossRef]
- Qosa, H.; Mohamed, L.A.; Batarseh, Y.S.; Alqahtani, S.; Ibrahim, B.; LeVine, H.; Keller, J.N.; Kaddoumi, A. Extra-Virgin Olive Oil Attenuates Amyloid-β and Tau Pathologies in the Brains of TgSwDI Mice. J. Nutr. Biochem. 2015, 26, 1479–1490. [Google Scholar] [CrossRef] [Green Version]
- Crespo, M.C.; Tomé-Carneiro, J.; Pintado, C.; Dávalos, A.; Visioli, F.; Burgos-Ramos, E. Hydroxytyrosol Restores Proper Insulin Signaling in an Astrocytic Model of Alzheimer’s Disease: Astrocytic Model of Alzheimer’s Disease. BioFactors 2017, 43, 540–548. [Google Scholar] [CrossRef]
- Feng, X.; Liang, N.; Zhu, D.; Gao, Q.; Peng, L.; Dong, H.; Yue, Q.; Liu, H.; Bao, L.; Zhang, J.; et al. Resveratrol Inhibits β-Amyloid-Induced Neuronal Apoptosis through Regulation of SIRT1-ROCK1 Signaling Pathway. PLoS ONE 2013, 8, e59888. [Google Scholar] [CrossRef] [Green Version]
- Baumgart, M.; Snyder, H.M.; Carrillo, M.C.; Fazio, S.; Kim, H.; Johns, H. Summary of the Evidence on Modifiable Risk Factors for Cognitive Decline and Dementia: A Population-Based Perspective. Alzheimer’s Dement. 2015, 11, 718–726. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Lapiscina, E.H.; Clavero, P.; Toledo, E.; Estruch, R.; Salas-Salvadó, J.; San Julián, B.; Sanchez-Tainta, A.; Ros, E.; Valls-Pedret, C.; Martinez-Gonzalez, M.Á. Mediterranean Diet Improves Cognition: The PREDIMED-NAVARRA Randomised Trial. J. Neurol. Neurosurg. Psychiatry 2013, 84, 1318–1325. [Google Scholar] [CrossRef] [Green Version]
- Valls-Pedret, C.; Sala-Vila, A.; Serra-Mir, M.; Corella, D.; de la Torre, R.; Martínez-González, M.Á.; Martínez-Lapiscina, E.H.; Fitó, M.; Pérez-Heras, A.; Salas-Salvadó, J.; et al. Mediterranean Diet and Age-Related Cognitive Decline: A Randomized Clinical Trial. JAMA Intern. Med. 2015, 175, 1094–1103. [Google Scholar] [CrossRef] [Green Version]
- Paknahad, Z.; Sheklabadi, E.; Derakhshan, Y.; Bagherniya, M.; Chitsaz, A. The Effect of the Mediterranean Diet on Cognitive Function in Patients with Parkinson’s Disease: A Randomized Clinical Controlled Trial. Complement. Ther. Med. 2020, 50, 102366. [Google Scholar] [CrossRef] [PubMed]
- Okubo, H.; Miyake, Y.; Sasaki, S.; Murakami, K.; Tanaka, K.; Fukushima, W.; Kiyohara, C.; Tsuboi, Y.; Yamada, T.; Oeda, T.; et al. Dietary Patterns and Risk of Parkinson’s Disease: A Case-Control Study in Japan: Dietary Patterns and Parkinson’s Disease. Eur. J. Neurol. 2012, 19, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Kihira, T.; Kobashi, G.; Washio, M.; Sasaki, S.; Yokoyama, T.; Miyake, Y.; Sakamoto, N.; Inaba, Y.; Nagai, M. Fruit and Vegetable Intake and Risk of Amyotrophic Lateral Sclerosis in Japan. Neuroepidemiology 2009, 32, 251–256. [Google Scholar] [CrossRef]
- Rivadeneyra, J.; Cubo, E.; Gil, C.; Calvo, S.; Mariscal, N.; Martínez, A. Factors Associated with Mediterranean Diet Adherence in Huntington’s Disease. Clin. Nutr. Espen 2016, 12, e7–e13. [Google Scholar] [CrossRef]
- Fidyt, K.; Fiedorowicz, A.; Strządała, L.; Szumny, A. β-Caryophyllene and β-Caryophyllene Oxide-Natural Compounds of Anticancer and Analgesic Properties. Cancer Med. 2016, 5, 3007–3017. [Google Scholar] [CrossRef]
- Russo, E.B. Taming THC: Potential Cannabis Synergy and Phytocannabinoid-Terpenoid Entourage Effects: Phytocannabinoid-Terpenoid Entourage Effects. Br. J. Pharmcol. 2011, 163, 1344–1364. [Google Scholar] [CrossRef] [PubMed]
- Machado, K.d.C.; Islam, M.T.; Ali, E.S.; Rouf, R.; Uddin, S.J.; Dev, S.; Shilpi, J.A.; Shill, M.C.; Reza, H.M.; Das, A.K.; et al. A Systematic Review on the Neuroprotective Perspectives of Beta-Caryophyllene: Neurobiological Effects of β-Caryophyllene. Phytother. Res. 2018, 32, 2376–2388. [Google Scholar] [CrossRef] [PubMed]
- Borge, G.I.A.; Sandberg, E.; Øyaas, J.; Abrahamsen, R.K. Variation of Terpenes in Milk and Cultured Cream from Norwegian Alpine Rangeland-Fed and in-Door Fed Cows. Food Chem. 2016, 199, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Chicca, A.; Caprioglio, D.; Minassi, A.; Petrucci, V.; Appendino, G.; Taglialatela-Scafati, O.; Gertsch, J. Functionalization of β-Caryophyllene Generates Novel Polypharmacology in the Endocannabinoid System. ACS Chem. Biol. 2014, 9, 1499–1507. [Google Scholar] [CrossRef]
- Gonçalves, E.C.D.; Baldasso, G.M.; Bicca, M.A.; Paes, R.S.; Capasso, R.; Dutra, R.C. Terpenoids, Cannabimimetic Ligands, beyond the Cannabis Plant. Molecules 2020, 25, 1567. [Google Scholar] [CrossRef] [Green Version]
- Gertsch, J.; Leonti, M.; Raduner, S.; Racz, I.; Chen, J.-Z.; Xie, X.-Q.; Altmann, K.-H.; Karsak, M.; Zimmer, A. Beta-Caryophyllene Is a Dietary Cannabinoid. Proc. Natl. Acad. Sci. USA 2008, 105, 9099–9104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, C.M.; Al Kaabi, J.M.; Nurulain, S.N.; Goyal, S.; Amjad Kamal, M.; Ojha, S. Polypharmacological Properties and Therapeutic Potential of β-Caryophyllene: A Dietary Phytocannabinoid of Pharmaceutical Promise. Curr. Pharm. Des. 2016, 22, 3237–3264. [Google Scholar] [CrossRef] [PubMed]
- Glass, C.K.; Saijo, K.; Winner, B.; Marchetto, M.C.; Gage, F.H. Mechanisms Underlying Inflammation in Neurodegeneration. Cell 2010, 140, 918–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Zeng, Z.; Wang, B.; Guo, S. Trans-Caryophyllene Inhibits Amyloid β (Aβ) Oligomer-Induced Neuroinflammation in BV-2 Microglial Cells. Int. Immunopharmacol. 2017, 51, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Dong, Z.; Liu, S. β-Caryophyllene Ameliorates the Alzheimer-like Phenotype in APP/PS1 Mice through CB2 Receptor Activation and the PPARγ Pathway. Pharmacology 2014, 94, 1–12. [Google Scholar] [CrossRef]
- Ojha, S.; Javed, H.; Azimullah, S.; Haque, M.E. β-Caryophyllene, a Phytocannabinoid Attenuates Oxidative Stress, Neuroinflammation, Glial Activation, and Salvages Dopaminergic Neurons in a Rat Model of Parkinson Disease. Mol. Cell Biochem. 2016, 418, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Alberti, T.; Barbosa, W.; Vieira, J.; Raposo, N.; Dutra, R. (−)-β-Caryophyllene, a CB2 Receptor-Selective Phytocannabinoid, Suppresses Motor Paralysis and Neuroinflammation in a Murine Model of Multiple Sclerosis. Int. J. Mol. Sci. 2017, 18, 691. [Google Scholar] [CrossRef]
- Gertsch, J. Antiinflammatory Cannabinoids in Diet—Towards a Better Understanding of CB2 Receptor Action?: Towards a Better Understanding of CB2 Receptor Action? Commun. Integr. Biol. 2008, 1, 26–28. [Google Scholar] [CrossRef]
- Navarro, G.; Morales, P.; Rodríguez-Cueto, C.; Fernández-Ruiz, J.; Jagerovic, N.; Franco, R. Targeting Cannabinoid CB2 Receptors in the Central Nervous System. Medicinal Chemistry Approaches with Focus on Neurodegenerative Disorders. Front. Neurosci. 2016, 10, 406. [Google Scholar] [CrossRef] [Green Version]
- Saha, R.N.; Pahan, K. Regulation of Inducible Nitric Oxide Synthase Gene in Glial Cells. Antioxid. Redox Signal. 2006, 8, 929–947. [Google Scholar] [CrossRef]
- Kim, H.-Y.; Hwang, K.W.; Park, S.-Y. Extracts of Actinidia Arguta Stems Inhibited LPS-Induced Inflammatory Responses through Nuclear Factor–ΚB Pathway in Raw 264.7 Cells. Nutr. Res. 2014, 34, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Segat, G.C.; Manjavachi, M.N.; Matias, D.O.; Passos, G.F.; Freitas, C.S.; Costa, R.; Calixto, J.B. Antiallodynic Effect of β-Caryophyllene on Paclitaxel-Induced Peripheral Neuropathy in Mice. Neuropharmacology 2017, 125, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Wu, X.; Yan, S.; Xie, X.; Fan, Y.; Zhang, J.; Peng, C.; You, Z. The Antidepressant-like Effects of Pioglitazone in a Chronic Mild Stress Mouse Model Are Associated with PPARγ-Mediated Alteration of Microglial Activation Phenotypes. J. Neuroinflamm. 2016, 13, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Sullivan, S.E.; Kendall, D.A.; Randall, M.D. Time-Dependent Vascular Effects of Endocannabinoids Mediated by Peroxisome Proliferator-Activated Receptor Gamma (PPARγ). PPAR Res. 2009, 2009, 425289. [Google Scholar] [CrossRef] [Green Version]
- Geldmacher, D.S.; Fritsch, T.; McClendon, M.J.; Landreth, G. A Randomized Pilot Clinical Trial of the Safety of Pioglitazone in Treatment of Patients With Alzheimer Disease. Arch. Neurol. 2011, 68, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Xu, H.E. Couple Dynamics: PPARγ and Its Ligand Partners. Structure 2012, 20, 2–4. [Google Scholar] [CrossRef] [Green Version]
- Pinto, A.; Bonucci, A.; Maggi, E.; Corsi, M.; Businaro, R. Anti-Oxidant and Anti-Inflammatory Activity of Ketogenic Diet: New Perspectives for Neuroprotection in Alzheimer’s Disease. Antioxidants 2018, 7, 63. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Sun, T.; Wang, X. Activation of Type 2 Cannabinoid Receptors (CB2R) Promotes Fatty Acid Oxidation through the SIRT1/PGC-1α Pathway. Biochem. Biophys. Res. Commun. 2013, 436, 377–381. [Google Scholar] [CrossRef]
- Niedzielska, E.; Smaga, I.; Gawlik, M.; Moniczewski, A.; Stankowicz, P.; Pera, J.; Filip, M. Oxidative Stress in Neurodegenerative Diseases. Mol. Neurobiol. 2016, 53, 4094–4125. [Google Scholar] [CrossRef] [Green Version]
- Assis, L.C.; Straliotto, M.R.; Engel, D.; Hort, M.A.; Dutra, R.C.; de Bem, A.F. β-Caryophyllene Protects the C6 Glioma Cells against Glutamate-Induced Excitotoxicity through the Nrf2 Pathway. Neuroscience 2014, 279, 220–231. [Google Scholar] [CrossRef]
- Fontes, L.B.A.; Dias, D.d.S.; Aarestrup, B.J.V.; Aarestrup, F.M.; Da Silva Filho, A.A.; Corrêa, J.O.d.A. β-Caryophyllene Ameliorates the Development of Experimental Autoimmune Encephalomyelitis in C57BL/6 Mice. Biomed. Pharm. 2017, 91, 257–264. [Google Scholar] [CrossRef]
- Javed, H.; Azimullah, S.; Haque, M.E.; Ojha, S.K. Cannabinoid Type 2 (CB2) Receptors Activation Protects against Oxidative Stress and Neuroinflammation Associated Dopaminergic Neurodegeneration in Rotenone Model of Parkinson’s Disease. Front. Neurosci. 2016, 10, 321. [Google Scholar] [CrossRef] [Green Version]
- Calleja, M.A.; Vieites, J.M.; Montero-Meterdez, T.; Torres, M.I.; Faus, M.J.; Gil, A.; Suárez, A. The Antioxidant Effect of β-Caryophyllene Protects Rat Liver from Carbon Tetrachloride-Induced Fibrosis by Inhibiting Hepatic Stellate Cell Activation. Br. J. Nutr. 2013, 109, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Baird, L.; Swift, S.; Llères, D.; Dinkova-Kostova, A.T. Monitoring Keap1–Nrf2 Interactions in Single Live Cells. Biotechnol. Adv. 2014, 32, 1133–1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tebay, L.E.; Robertson, H.; Durant, S.T.; Vitale, S.R.; Penning, T.M.; Dinkova-Kostova, A.T.; Hayes, J.D. Mechanisms of Activation of the Transcription Factor Nrf2 by Redox Stressors, Nutrient Cues, and Energy Status and the Pathways through Which It Attenuates Degenerative Disease. Free Radic. Biol. Med. 2015, 88, 108–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Cabrerizo, R.; García-Fuster, M.J. Opposite Regulation of Cannabinoid CB1 and CB2 Receptors in the Prefrontal Cortex of Rats Treated with Cocaine during Adolescence. Neurosci. Lett. 2016, 615, 60–65. [Google Scholar] [CrossRef]
- Sierra, S.; Luquin, N.; Rico, A.J.; Gómez-Bautista, V.; Roda, E.; Dopeso-Reyes, I.G.; Vázquez, A.; Martínez-Pinilla, E.; Labandeira-García, J.L.; Franco, R.; et al. Detection of Cannabinoid Receptors CB1 and CB2 within Basal Ganglia Output Neurons in Macaques: Changes Following Experimental Parkinsonism. Brain Struct. Funct. 2015, 220, 2721–2738. [Google Scholar] [CrossRef] [Green Version]
- Callén, L.; Moreno, E.; Barroso-Chinea, P.; Moreno-Delgado, D.; Cortés, A.; Mallol, J.; Casadó, V.; Lanciego, J.L.; Franco, R.; Lluis, C.; et al. Cannabinoid Receptors CB1 and CB2 Form Functional Heteromers in Brain. J. Biol. Chem. 2012, 287, 20851–20865. [Google Scholar] [CrossRef] [Green Version]
- Kawai, Y.; Garduño, L.; Theodore, M.; Yang, J.; Arinze, I.J. Acetylation-Deacetylation of the Transcription Factor Nrf2 (Nuclear Factor Erythroid 2-Related Factor 2) Regulates Its Transcriptional Activity and Nucleocytoplasmic Localization. J. Biol. Chem. 2011, 286, 7629–7640. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.-J.; Zhao, X.-H.; Chen, W.; Bo, N.; Wang, X.-J.; Chi, Z.-F.; Wu, W. Sirtuin 1 Activation Enhances the PGC-1α/Mitochondrial Antioxidant System Pathway in Status Epilepticus. Mol. Med. Rep. 2015, 11, 521–526. [Google Scholar] [CrossRef] [Green Version]
- Puigserver, P.; Spiegelman, B.M. Peroxisome Proliferator-Activated Receptor-γ Coactivator 1α (PGC-1α): Transcriptional Coactivator and Metabolic Regulator. Endocr. Rev. 2003, 24, 78–90. [Google Scholar] [CrossRef] [Green Version]
- Olmos, Y.; Sánchez-Gómez, F.J.; Wild, B.; García-Quintans, N.; Cabezudo, S.; Lamas, S.; Monsalve, M. SirT1 Regulation of Antioxidant Genes Is Dependent on the Formation of a FoxO3a/PGC-1α Complex. Antioxid. Redox Signal. 2013, 19, 1507–1521. [Google Scholar] [CrossRef] [PubMed]
- Freitas, A.E.; Egea, J.; Buendia, I.; Gómez-Rangel, V.; Parada, E.; Navarro, E.; Casas, A.I.; Wojnicz, A.; Ortiz, J.A.; Cuadrado, A.; et al. Agmatine, by Improving Neuroplasticity Markers and Inducing Nrf2, Prevents Corticosterone-Induced Depressive-Like Behavior in Mice. Mol. Neurobiol. 2016, 53, 3030–3045. [Google Scholar] [CrossRef] [PubMed]
- Galán-Ganga, M.; del Río, R.; Jiménez-Moreno, N.; Díaz-Guerra, M.; Lastres-Becker, I. Cannabinoid CB2 Receptor Modulation by the Transcription Factor NRF2 Is Specific in Microglial Cells. Cell Mol. Neurobiol. 2020, 40, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Kagoya, Y.; Yoshimi, A.; Kataoka, K.; Nakagawa, M.; Kumano, K.; Arai, S.; Kobayashi, H.; Saito, T.; Iwakura, Y.; Kurokawa, M. Positive Feedback between NF-ΚB and TNF-α Promotes Leukemia-Initiating Cell Capacity. J. Clin. Investig. 2014, 124, 528–542. [Google Scholar] [CrossRef]
- Choi, I.-Y.; Ju, C.; Anthony Jalin, A.M.A.; Lee, D.I.; Prather, P.L.; Kim, W.-K. Activation of Cannabinoid CB2 Receptor–Mediated AMPK/CREB Pathway Reduces Cerebral Ischemic Injury. Am. J. Pathol. 2013, 182, 928–939. [Google Scholar] [CrossRef] [PubMed]
- Wrann, C.D.; White, J.P.; Salogiannnis, J.; Laznik-Bogoslavski, D.; Wu, J.; Ma, D.; Lin, J.D.; Greenberg, M.E.; Spiegelman, B.M. Exercise Induces Hippocampal BDNF through a PGC-1α/FNDC5 Pathway. Cell Metab. 2013, 18, 649–659. [Google Scholar] [CrossRef] [Green Version]
- Eyileten, C.; Kaplon-Cieslicka, A.; Mirowska-Guzel, D.; Malek, L.; Postula, M. Antidiabetic Effect of Brain-Derived Neurotrophic Factor and Its Association with Inflammation in Type 2 Diabetes Mellitus. J. Diabetes Res. 2017, 2017, 2823671. [Google Scholar] [CrossRef]
- Giampà, C.; Montagna, E.; Dato, C.; Melone, M.A.B.; Bernardi, G.; Fusco, F.R. Systemic Delivery of Recombinant Brain Derived Neurotrophic Factor (BDNF) in the R6/2 Mouse Model of Huntington’s Disease. PLoS ONE 2013, 8, e64037. [Google Scholar] [CrossRef]
- Avraham, H.K.; Jiang, S.; Fu, Y.; Rockenstein, E.; Makriyannis, A.; Zvonok, A.; Masliah, E.; Avraham, S. The Cannabinoid CB2 Receptor Agonist AM1241 Enhances Neurogenesis in GFAP/Gp120 Transgenic Mice Displaying Deficits in Neurogenesis: Effects of CB2 Agonist on Neurogenesis. Br. J. Pharmcol. 2014, 171, 468–479. [Google Scholar] [CrossRef] [Green Version]
- Palazuelos, J.; Aguado, T.; Egia, A.; Mechoulam, R.; Guzmán, M.; Galve-Roperh, I.; Palazuelos, J.; Aguado, T.; Egia, A.; Mechoulam, R.; et al. Non-psychoactive CB2 Cannabinoid Agonists Stimulate Neural Progenitor Proliferation. FASEB J. 2006, 20, 2405–2407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, M.; Sherchan, P.; Krafft, P.R.; Rolland, W.B.; Soejima, Y.; Zhang, J.H. Cannabinoid Type 2 Receptor Stimulation Attenuates Brain Edema by Reducing Cerebral Leukocyte Infiltration Following Subarachnoid Hemorrhage in Rats. J. Neurol. Sci. 2014, 342, 101–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, X.; Peng, J.; Zhong, J.; Yang, M.; Pang, J.; Lou, J.; Li, M.; An, R.; Zhang, Q.; Xu, L.; et al. β-Caryophyllene Protects in Vitro Neurovascular Unit against Oxygen-Glucose Deprivation and Re-Oxygenation-Induced Injury. J. Neurochem. 2016, 139, 757–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, J.; Cao, G.; Li, R.; Liu, J.; Dong, Z.; Xu, L. β-Caryophyllene Attenuates Focal Cerebral Ischemia-Reperfusion Injury by Nrf2/HO-1 Pathway in Rats. Neurochem. Res. 2016, 41, 1291–1304. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, N.; Allart-Simon, I.; De Nicola, G.R.; Iori, R.; Renault, J.-H.; Rollin, P.; Nuzillard, J.-M. Advanced NMR-Based Structural Investigation of Glucosinolates and Desulfoglucosinolates. J. Nat. Prod. 2018, 81, 323–334. [Google Scholar] [CrossRef]
- Fuentes, F.; Paredes-Gonzalez, X.; Kong, A.-N.T. Dietary Glucosinolates Sulforaphane, Phenethyl Isothiocyanate, Indole-3-Carbinol/3,3′-Diindolylmethane: Antioxidative Stress/Inflammation, Nrf2, Epigenetics/Epigenomics and In Vivo Cancer Chemopreventive Efficacy. Curr. Pharm. Rep. 2015, 1, 179–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNaughton, S.A.; Marks, G.C. Development of a Food Composition Database for the Estimation of Dietary Intakes of Glucosinolates, the Biologically Active Constituents of Cruciferous Vegetables. Br. J. Nutr. 2003, 90, 687–697. [Google Scholar] [CrossRef] [Green Version]
- Hayes, J.D.; Kelleher, M.O.; Eggleston, I.M. The Cancer Chemopreventive Actions of Phytochemicals Derived from Glucosinolates. Eur. J. Nutr. 2008, 47, 73–88. [Google Scholar] [CrossRef]
- Link, L.B.; Potter, J.D. Raw versus Cooked Vegetables and Cancer Risk. Cancer Epidemiol. Biomark. Prev. 2004, 13, 1422–1435. [Google Scholar]
- Calabrese, V.; Cornelius, C.; Trovato-Salinaro, A.; Cambria, M.; Locascio, M.; Rienzo, L.; Condorelli, D.; Mancuso, C.; De Lorenzo, A.; Calabrese, E. The Hormetic Role of Dietary Antioxidants in Free Radical-Related Diseases. Curr. Pharm. Des. 2010, 16, 877–883. [Google Scholar] [CrossRef]
- Zhang, W.W.; Feng, Z.; Narod, S.A. Multiple Therapeutic and Preventive Effects of 3,39-Diindolylmethane on Cancers Including Prostate Cancer and High Grade Prostatic Intraepithelial Neoplasia. J. Biomed. Res. 2014, 339–348. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Cheng, L.; Liu, Y.; Wang, J.; Jiang, W. Indole-3-Carbinol (I3C) and Its Major Derivatives: Their Pharmacokinetics and Important Roles in Hepatic Protection. Curr. Drug Metab. 2016, 17, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kong, D.; Ahmad, A.; Bao, B.; Sarkar, F.H. Antioxidant Function of Isoflavone and 3,3′-Diindolylmethane: Are They Important for Cancer Prevention and Therapy? Antioxid. Redox Signal. 2013, 19, 139–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rzemieniec, J.; Bratek, E.; Wnuk, A.; Przepiórska, K.; Salińska, E.; Kajta, M. Neuroprotective Effect of 3,3’-Diindolylmethane against Perinatal Asphyxia Involves Inhibition of the AhR and NMDA Signaling and Hypermethylation of Specific Genes. Apoptosis 2020, 25, 747–762. [Google Scholar] [CrossRef] [PubMed]
- Anderton, M.J.; Manson, M.M.; Verschoyle, R.; Gescher, A.; Steward, W.P.; Williams, M.L.; Mager, D.E. Physiological Modeling of Formulated and Crystalline 3,3’-Diindolylmethane Pharmacokinetics Following Oral Administration in Mice. Drug Metab. Dispos. 2004, 32, 632–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, G.A.; Sunega, J.M.; Sullivan, D.K.; Gray, J.C.; Mayo, M.S.; Crowell, J.A.; Hurwitz, A. Single-Dose Pharmacokinetics and Tolerability of Absorption-Enhanced 3,3’-Diindolylmethane in Healthy Subjects. Cancer Epidemiol. Biomark. Prev. 2008, 17, 2619–2624. [Google Scholar] [CrossRef] [Green Version]
- Dalessandri, K.M.; Firestone, G.L.; Fitch, M.D.; Bradlow, H.L.; Bjeldanes, L.F. Pilot Study: Effect of 3,3′-Diindolylmethane Supplements on Urinary Hormone Metabolites in Postmenopausal Women With a History of Early-Stage Breast Cancer. Nutr. Cancer 2004, 50, 161–167. [Google Scholar] [CrossRef] [Green Version]
- Nair, A.; Jacob, S. A Simple Practice Guide for Dose Conversion between Animals and Human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.D.; Yoo, J.-M.; Baek, S.Y.; Li, F.Y.; Sok, D.-E.; Kim, M.R. 3,3′-Diindolylmethane Promotes BDNF and Antioxidant Enzyme Formation via TrkB/Akt Pathway Activation for Neuroprotection against Oxidative Stress-Induced Apoptosis in Hippocampal Neuronal Cells. Antioxidants 2019, 9, 3. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.J.; Seon, M.R.; Lee, Y.M.; Kim, J.; Kim, J.-K.; Kim, S.G.; Park, J.H.Y. 3,3′-Diindolylmethane Suppresses the Inflammatory Response to Lipopolysaccharide in Murine Macrophages. J. Nutr. 2008, 138, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.W.; Kim, J.; Kim, J.; Lee, S.; Choi, B.-R.; Han, J.-S.; Lee, K.W.; Lee, H.J. 3,3’-Diindolylmethane Inhibits Lipopolysaccharide-Induced Microglial Hyperactivation and Attenuates Brain Inflammation. Toxicol. Sci. 2014, 137, 158–167. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.K.; Choi, E.-J. Pathological Roles of MAPK Signaling Pathways in Human Diseases. Biochim. Biophys. Acta Mol. Basis Dis. 2010, 1802, 396–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rzemieniec, J.; Wnuk, A.; Lasoń, W.; Bilecki, W.; Kajta, M. The Neuroprotective Action of 3,3′-Diindolylmethane against Ischemia Involves an Inhibition of Apoptosis and Autophagy That Depends on HDAC and AhR/CYP1A1 but Not ERα/CYP19A1 Signaling. Apoptosis 2019, 24, 435–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, K.; Eguchi, Y.; Imagawa, Y.; Akai, S.; Mochizuki, H.; Tsujimoto, Y. MPP+ Induces Necrostatin-1- and Ferrostatin-1-Sensitive Necrotic Death of Neuronal SH-SY5Y Cells. Cell Death Discov. 2017, 3, 17013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Miranda, B.R.; Miller, J.A.; Hansen, R.J.; Lunghofer, P.J.; Safe, S.; Gustafson, D.L.; Colagiovanni, D.; Tjalkens, R.B. Neuroprotective Efficacy and Pharmacokinetic Behavior of Novel Anti-Inflammatory Para-Phenyl Substituted Diindolylmethanes in a Mouse Model of Parkinson’s Disease. J. Pharmacol. Exp. Ther. 2013, 345, 125–138. [Google Scholar] [CrossRef]
- Qin, C.; Morrow, D.; Stewart, J.; Spencer, K.; Porter, W.; Smith, R.; Phillips, T.; Abdelrahim, M.; Samudio, I.; Safe, S. A New Class of Peroxisome Proliferator-Activated Receptor Gamma (PPARgamma) Agonists That Inhibit Growth of Breast Cancer Cells: 1,1-Bis(3’-Indolyl)-1-(p-Substituted Phenyl)Methanes. Mol. Cancer Ther. 2004, 3, 247–260. [Google Scholar]
- Almeida, R.D.; Manadas, B.J.; Melo, C.V.; Gomes, J.R.; Mendes, C.S.; Grãos, M.M.; Carvalho, R.F.; Carvalho, A.P.; Duarte, C.B. Neuroprotection by BDNF against Glutamate-Induced Apoptotic Cell Death Is Mediated by ERK and PI3-Kinase Pathways. Cell Death Differ. 2005, 12, 1329–1343. [Google Scholar] [CrossRef] [Green Version]
- Saw, C.L.-L.; Cintrón, M.; Wu, T.-Y.; Guo, Y.; Huang, Y.; Jeong, W.-S.; Kong, A.-N.T. Pharmacodynamics of Dietary Phytochemical Indoles I3C and DIM: Induction of Nrf2-Mediated Phase II Drug Metabolizing and Antioxidant Genes and Synergism with Isothiocyanates. Biopharm. Drug Dispos. 2011, 32, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Gertsch, J. Cannabimimetic Phytochemicals in the Diet—An Evolutionary Link to Food Selection and Metabolic Stress Adaptation?: Cannabimimetic Phytochemicals in the Diet. Br. J. Pharmcol. 2017, 174, 1464–1483. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.; Chu, A.; Li, W.; Wang, B.; Shelton, F.; Otero, F.; Nguyen, D.G.; Caldwell, J.S.; Chen, Y.A. Lipid G Protein-Coupled Receptor Ligand Identification Using β-Arrestin PathHunterTM Assay. J. Biol. Chem. 2009, 284, 12328–12338. [Google Scholar] [CrossRef] [Green Version]
- Brunetti, L.; Loiodice, F.; Piemontese, L.; Tortorella, P.; Laghezza, A. New Approaches to Cancer Therapy: Combining Fatty Acid Amide Hydrolase (FAAH) Inhibition with Peroxisome Proliferator-Activated Receptors (PPARs) Activation: Miniperspective. J. Med. Chem. 2019, 62, 10995–11003. [Google Scholar] [CrossRef] [PubMed]
- Toczek, M.; Malinowska, B. Enhanced Endocannabinoid Tone as a Potential Target of Pharmacotherapy. Life Sci. 2018, 204, 20–45. [Google Scholar] [CrossRef] [PubMed]
- Fowler, C.J.; Stenström, A.; Tiger, G. Ibuprofen Inhibits the Metabolism of the Endogenous Cannabimimetic Agent Anandamide. Pharm. Toxicol. 1997, 80, 103–107. [Google Scholar] [CrossRef]
- Lehmann, J.M.; Lenhard, J.M.; Oliver, B.B.; Ringold, G.M.; Kliewer, S.A. Peroxisome Proliferator-Activated Receptors α and γ Are Activated by Indomethacin and Other Non-Steroidal Anti-Inflammatory Drugs. J. Biol. Chem. 1997, 272, 3406–3410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thors, L.; Belghiti, M.; Fowler, C.J. Inhibition of Fatty Acid Amide Hydrolase by Kaempferol and Related Naturally Occurring Flavonoids: Inhibition of FAAH by Flavonoids. Br. J. Pharmcol. 2008, 155, 244–252. [Google Scholar] [CrossRef] [Green Version]
- Lenman, A.; Fowler, C.J. Interaction of Ligands for the Peroxisome Proliferator-Activated Receptor γ with the Endocannabinoid System: Ciglitazone Inhibits FAAH Activity. Br. J. Pharmcol. 2007, 151, 1343–1351. [Google Scholar] [CrossRef] [Green Version]
- Winkel-Shirley, B. Flavonoid Biosynthesis. A Colorful Model for Genetics, Biochemistry, Cell Biology, and Biotechnology. Plant. Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef] [Green Version]
- Graf, B.A.; Milbury, P.E.; Blumberg, J.B. Flavonols, Flavones, Flavanones, and Human Health: Epidemiological Evidence. J. Med. Food 2005, 8, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Wu, J.H.Y. Flavonoids, Dairy Foods, and Cardiovascular and Metabolic Health: A Review of Emerging Biologic Pathways. Circ. Res. 2018, 122, 369–384. [Google Scholar] [CrossRef]
- Cassidy, A.; Minihane, A.-M. The Role of Metabolism (and the Microbiome) in Defining the Clinical Efficacy of Dietary Flavonoids. Am. J. Clin. Nutr. 2017, 105, 10–22. [Google Scholar] [CrossRef] [Green Version]
- Reiter, E.; Beck, V.; Medjakovic, S.; Mueller, M.; Jungbauer, A. Comparison of Hormonal Activity of Isoflavone-Containing Supplements Used to Treat Menopausal Complaints. Menopause 2009, 16, 1049–1060. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarczyk-Sedlak, I.; Zych, M.; Wojnar, W.; Ozimina-Kamińska, E.; Dudek, S.; Chadała, N.; Kachel, A. Biochanin a Shows No Effect on Skeletal System in Ovariectomized Rats, When Administered in Moderate Dose. Acta Pol. Pharm. 2015, 72, 587–596. [Google Scholar]
- Lv, Q.; Yang, Y.; Zhao, Y.; Gu, D.; He, D.; Yili, A.; Ma, Q.; Cheng, Z.; Gao, Y.; Aisa, H.A.; et al. Comparative Study on Separation and Purification of Isoflavones from the Seeds and Sprouts of Chickpea by High-Speed Countercurrent Chromatography. J. Liq. Chromatogr. Relat. Technol. 2009, 32, 2879–2892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarfraz, A.; Javeed, M.; Shah, M.A.; Hussain, G.; Shafiq, N.; Sarfraz, I.; Riaz, A.; Sadiqa, A.; Zara, R.; Zafar, S.; et al. Biochanin A: A Novel Bioactive Multifunctional Compound from Nature. Sci. Total Environ. 2020, 722, 137907. [Google Scholar] [CrossRef] [PubMed]
- Thors, L.; Burston, J.; Alter, B.; McKinney, M.; Cravatt, B.; Ross, R.; Pertwee, R.; Gereau, R.; Wiley, J.; Fowler, C. Biochanin A, a Naturally Occurring Inhibitor of Fatty Acid Amide Hydrolase: Inhibition of FAAH by Isoflavones. Br. J. Pharmcol. 2010, 160, 549–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howes, J.; Waring, M.; Huang, L.; Howes, L.G. Long-Term Pharmacokinetics of an Extract of Isoflavones from Red Clover (Trifolium Pratense). J. Altern. Complement. Med. 2002, 8, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Moon, Y.J.; Sagawa, K.; Frederick, K.; Zhang, S.; Morris, M.E. Pharmacokinetics and Bioavailability of the Isoflavone Biochanin A in Rats. AAPS J. 2006, 8, E433–E442. [Google Scholar] [CrossRef] [Green Version]
- Thors, L.; Alajakku, K.; Fowler, C.J. The ‘Specific’ Tyrosine Kinase Inhibitor Genistein Inhibits the Enzymic Hydrolysis of Anandamide: Implications for Anandamide Uptake: Genistein Inhibits FAAH Activity. Br. J. Pharmcol. 2007, 150, 951–960. [Google Scholar] [CrossRef] [Green Version]
- Miadoková, E. Isoflavonoids—An Overview of Their Biological Activities and Potential Health Benefits. Interdiscip. Toxicol. 2009, 2, 211–218. [Google Scholar] [CrossRef]
- Dang, Z.-C.; Audinot, V.; Papapoulos, S.E.; Boutin, J.A.; Löwik, C.W.G.M. Peroxisome Proliferator-Activated Receptor γ (PPARγ) as a Molecular Target for the Soy Phytoestrogen Genistein. J. Biol. Chem. 2003, 278, 962–967. [Google Scholar] [CrossRef] [Green Version]
- Thors, L.; Eriksson, J.; Fowler, C.J. Inhibition of the Cellular Uptake of Anandamide by Genistein and Its Analogue Daidzein in Cells with Different Levels of Fatty Acid Amide Hydrolase-Driven Uptake: Genistein, Daidzein and Anandamide Uptake. Br. J. Pharmcol. 2007, 152, 744–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chacko, B.K.; Chandler, R.T.; D’Alessandro, T.L.; Mundhekar, A.; Khoo, N.K.H.; Botting, N.; Barnes, S.; Patel, R.P. Anti-Inflammatory Effects of Isoflavones Are Dependent on Flow and Human Endothelial Cell PPARγ. J. Nutr. 2007, 137, 351–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasilopoulou, E.; Georga, K.; Joergensen, M.; Naska, A.; Trichopoulou, A. The Antioxidant Properties of Greek Foods and the Flavonoid Content of the Mediterranean Menu. CMCIEMA 2005, 5, 33–45. [Google Scholar] [CrossRef]
- Sureda, A.; Sanches Silva, A.; Sánchez-Machado, D.I.; López-Cervantes, J.; Daglia, M.; Nabavi, S.F.; Nabavi, S.M. Hypotensive Effects of Genistein: From Chemistry to Medicine. Chem. Biol. Interact. 2017, 268, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Subedi, L.; Ji, E.; Shin, D.; Jin, J.; Yeo, J.; Kim, S. Equol, a Dietary Daidzein Gut Metabolite Attenuates Microglial Activation and Potentiates Neuroprotection In Vitro. Nutrients 2017, 9, 207. [Google Scholar] [CrossRef] [Green Version]
- López, P.; Sánchez, M.; Perez-Cruz, C.; Velázquez-Villegas, L.A.; Syeda, T.; Aguilar-López, M.; Rocha-Viggiano, A.K.; del Carmen Silva-Lucero, M.; Torre-Villalvazo, I.; Noriega, L.G.; et al. Long-Term Genistein Consumption Modifies Gut Microbiota, Improving Glucose Metabolism, Metabolic Endotoxemia, and Cognitive Function in Mice Fed a High-Fat Diet. Mol. Nutr. Food Res. 2018, 62, 1800313. [Google Scholar] [CrossRef]
- Morton, M.S.; Arisaka, O.; Miyake, N.; Morgan, L.D.; Evans, B.A.J. Phytoestrogen Concentrations in Serum from Japanese Men and Women over Forty Years of Age. J. Nutr. 2002, 132, 3168–3171. [Google Scholar] [CrossRef] [PubMed]
- Woo, H.W.; Kim, M.K.; Lee, Y.-H.; Shin, D.H.; Shin, M.-H.; Choi, B.Y. Sex-Specific Associations of Habitual Intake of Soy Protein and Isoflavones with Risk of Type 2 Diabetes. Clin. Nutr. 2021, 40, 127–136. [Google Scholar] [CrossRef]
- Plumb, J.; Pigat, S.; Bompola, F.; Cushen, M.; Pinchen, H.; Nørby, E.; Astley, S.; Lyons, J.; Kiely, M.; Finglas, P. EBASIS (Bioactive Substances in Food Information Systems) and Bioactive Intakes: Major Updates of the Bioactive Compound Composition and Beneficial Bioeffects Database and the Development of a Probabilistic Model to Assess Intakes in Europe. Nutrients 2017, 9, 320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salehi, B.; Venditti, A.; Sharifi-Rad, M.; Kręgiel, D.; Sharifi-Rad, J.; Durazzo, A.; Lucarini, M.; Santini, A.; Souto, E.; Novellino, E.; et al. The Therapeutic Potential of Apigenin. Int. J. Mol. Sci. 2019, 20, 1305. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.-C.; Tsai, S.-H.; Tsai, D.-C.; Lin-Shiau, S.-Y.; Lin, J.-K. Suppression of Inducible Cyclooxygenase and Nitric Oxide Synthase through Activation of Peroxisome Proliferator-Activated Receptor-γ by Flavonoids in Mouse Macrophages. Febs Lett. 2001, 496, 12–18. [Google Scholar] [CrossRef] [Green Version]
- Venigalla, M.; Gyengesi, E.; Münch, G. Curcumin and Apigenin—Novel and Promising Therapeutics against Chronic Neuroinflammation in Alzheimer′s Disease. Neural Regen. Res. 2015, 10, 1181. [Google Scholar] [CrossRef] [PubMed]
- Hostetler, G.L.; Ralston, R.A.; Schwartz, S.J. Flavones: Food Sources, Bioavailability, Metabolism, and Bioactivity. Adv. Nutr. 2017, 8, 423–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tauchen, J.; Huml, L.; Rimpelova, S.; Jurášek, M. Flavonoids and Related Members of the Aromatic Polyketide Group in Human Health and Disease: Do They Really Work? Molecules 2020, 25, 3846. [Google Scholar] [CrossRef] [PubMed]
- Russo, E.B. Beyond Cannabis: Plants and the Endocannabinoid System. Trends Pharm. Sci. 2016, 37, 594–605. [Google Scholar] [CrossRef]
- Fang, X.-K.; Gao, J.; Zhu, D.-N. Kaempferol and Quercetin Isolated from Euonymus Alatus Improve Glucose Uptake of 3T3-L1 Cells without Adipogenesis Activity. Life Sci. 2008, 82, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Rauf, A.; Shah, Z.A.; Saeed, F.; Imran, A.; Arshad, M.U.; Ahmad, B.; Bawazeer, S.; Atif, M.; Peters, D.G.; et al. Chemo-Preventive and Therapeutic Effect of the Dietary Flavonoid Kaempferol: A Comprehensive Review: Chemo-Preventive and Therapeutic Effect of Kaempferol: A Comprehensive. Phytother. Res. 2019, 33, 263–275. [Google Scholar] [CrossRef]
- Yang, Y.-L.; Cheng, X.; Li, W.-H.; Liu, M.; Wang, Y.-H.; Du, G.-H. Kaempferol Attenuates LPS-Induced Striatum Injury in Mice Involving Anti-Neuroinflammation, Maintaining BBB Integrity, and Down-Regulating the HMGB1/TLR4 Pathway. Int. J. Mol. Sci. 2019, 20, 491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DuPont, M.S.; Day, A.J.; Bennett, R.N.; Mellon, F.A.; Kroon, P.A. Absorption of Kaempferol from Endive, a Source of Kaempferol-3-Glucuronide, in Humans. Eur. J. Clin. Nutr. 2004, 58, 947–954. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, H.; Rauf, K.; Zada, W.; McCarthy, M.; Abbas, G.; Anwar, F.; Shah, A.J. Kaempferol Facilitated Extinction Learning in Contextual Fear Conditioned Rats via Inhibition of Fatty-Acid Amide Hydrolase. Molecules 2020, 25, 4683. [Google Scholar] [CrossRef]
- Catalá, A. Five Decades with Polyunsaturated Fatty Acids: Chemical Synthesis, Enzymatic Formation, Lipid Peroxidation and Its Biological Effects. J. Lipids 2013, 2013, 710290. [Google Scholar] [CrossRef] [Green Version]
- Dyall, S.C. Long-Chain Omega-3 Fatty Acids and the Brain: A Review of the Independent and Shared Effects of EPA, DPA and DHA. Front. Aging Neurosci. 2015, 7, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domenichiello, A.F.; Kitson, A.P.; Bazinet, R.P. Is Docosahexaenoic Acid Synthesis from α-Linolenic Acid Sufficient to Supply the Adult Brain? Prog. Lipid Res. 2015, 59, 54–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyall, S.C. Interplay Between N-3 and n-6 Long-Chain Polyunsaturated Fatty Acids and the Endocannabinoid System in Brain Protection and Repair. Lipids 2017, 52, 885–900. [Google Scholar] [CrossRef] [PubMed]
- Bisogno, T.; Maccarrone, M. Endocannabinoid Signaling and Its Regulation by Nutrients: Endocannabinoid Signaling and Nutrients. BioFactors 2014, 40, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Li, Y.; Watkins, B.A. Endocannabinoid Signaling and Energy Metabolism: A Target for Dietary Intervention. Nutrition 2011, 27, 624–632. [Google Scholar] [CrossRef]
- Artmann, A.; Petersen, G.; Hellgren, L.I.; Boberg, J.; Skonberg, C.; Nellemann, C.; Hansen, S.H.; Hansen, H.S. Influence of Dietary Fatty Acids on Endocannabinoid and N-Acylethanolamine Levels in Rat Brain, Liver and Small Intestine. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2008, 1781, 200–212. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The Importance of the Ratio of Omega-6/Omega-3 Essential Fatty Acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- Casas, R.; Sacanella, E.; Estruch, R. The Immune Protective Effect of the Mediterranean Diet against Chronic Low-Grade Inflammatory Diseases. Endocr. Metab. Immune Disord. Drug Targets 2014, 14, 245–254. [Google Scholar] [CrossRef] [Green Version]
- Bosma-den Boer, M.M.; van Wetten, M.-L.; Pruimboom, L. Chronic Inflammatory Diseases Are Stimulated by Current Lifestyle: How Diet, Stress Levels and Medication Prevent Our Body from Recovering. Nutr. Metab. 2012, 9, 32. [Google Scholar] [CrossRef] [Green Version]
- Bigford, G.E.; Del Rossi, G. Supplemental Substances Derived from Foods as Adjunctive Therapeutic Agents for Treatment of Neurodegenerative Diseases and Disorders. Adv. Nutr. 2014, 5, 394–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panagiotakos, D.B.; Kastorini, C.-M.; Pitsavos, C.; Stefanadis, C. The Current Greek Diet and the Omega-6/Omega-3 Balance: The Mediterranean Diet Score Is Inversely Associated with the Omega-6/Omega-3 Ratio. In World Review of Nutrition and Dietetics; Simopoulos, A.P., Ed.; KARGER: Basel, Switzerland, 2011; Volume 102, pp. 53–56. ISBN 978-3-8055-9780-7. [Google Scholar]
- Scoditti, E.; Capurso, C.; Capurso, A.; Massaro, M. Vascular Effects of the Mediterranean Diet—Part II: Role of Omega-3 Fatty Acids and Olive Oil Polyphenols. Vasc. Pharmacol. 2014, 63, 127–134. [Google Scholar] [CrossRef]
- Massaro, M.; Scoditti, E.; Carluccio, M.A.; De Caterina, R. Basic Mechanisms behind the Effects of N-3 Fatty Acids on Cardiovascular Disease. Prostaglandins Leukot. Essent. Fat. Acids 2008, 79, 109–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, J.T.; Williams, J.S.; Pandarinathan, L.; Janero, D.R.; Lammi-Keefe, C.J.; Makriyannis, A. Dietary Docosahexaenoic Acid Supplementation Alters Select Physiological Endocannabinoid-System Metabolites in Brain and Plasma. J. Lipid Res. 2010, 51, 1416–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutchins-Wiese, H.L.; Li, Y.; Hannon, K.; Watkins, B.A. Hind Limb Suspension and Long-Chain Omega-3 PUFA Increase MRNA Endocannabinoid System Levels in Skeletal Muscle. J. Nutr. Biochem. 2012, 23, 986–993. [Google Scholar] [CrossRef]
- Campos, A.C.; Fogaça, M.V.; Sonego, A.B.; Guimarães, F.S. Cannabidiol, Neuroprotection and Neuropsychiatric Disorders. Pharmacol. Res. 2016, 112, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Larrieu, T.; Madore, C.; Joffre, C.; Layé, S. Nutritional N-3 Polyunsaturated Fatty Acids Deficiency Alters Cannabinoid Receptor Signaling Pathway in the Brain and Associated Anxiety-like Behavior in Mice. J. Physiol. Biochem. 2012, 68, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Lafourcade, M.; Larrieu, T.; Mato, S.; Duffaud, A.; Sepers, M.; Matias, I.; De Smedt-Peyrusse, V.; Labrousse, V.F.; Bretillon, L.; Matute, C.; et al. Nutritional Omega-3 Deficiency Abolishes Endocannabinoid-Mediated Neuronal Functions. Nat. Neurosci. 2011, 14, 345–350. [Google Scholar] [CrossRef] [Green Version]
- Thomazeau, A.; Bosch-Bouju, C.; Manzoni, O.; Layé, S. Nutritional N-3 PUFA Deficiency Abolishes Endocannabinoid Gating of Hippocampal Long-Term Potentiation. Cereb. Cortex 2016, 27, bhw052. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.-P.; Zhang, H.-Q.; Wang, W.; Guo, Y.-F.; Xiao, N.; Cao, X.-H.; Liu, L.-J. Some Subtypes of Endocannabinoid/Endovanilloid Receptors Mediate Docosahexaenoic Acid-Induced Enhanced Spatial Memory in Rats. Brain Res. 2011, 1412, 18–27. [Google Scholar] [CrossRef]
- Yurko-Mauro, K. Cognitive and Cardiovascular Benefits of Docosahexaenoic Acid in Aging and Cognitive Decline. Curr. Alzheimer Res. 2010, 7, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, V.; Stella, N.; Zimmer, A. Endocannabinoid Signalling and the Deteriorating Brain. Nat. Rev. Neurosci. 2015, 16, 30–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyall, S.C.; Michael, G.J.; Michael-Titus, A.T. Omega-3 Fatty Acids Reverse Age-Related Decreases in Nuclear Receptors and Increase Neurogenesis in Old Rats. J. Neurosci. Res. 2010, 88, 2091–2102. [Google Scholar] [CrossRef]
- Dyall, S.C.; Mandhair, H.K.; Fincham, R.E.A.; Kerr, D.M.; Roche, M.; Molina-Holgado, F. Distinctive Effects of Eicosapentaenoic and Docosahexaenoic Acids in Regulating Neural Stem Cell Fate Are Mediated via Endocannabinoid Signalling Pathways. Neuropharmacology 2016, 107, 387–395. [Google Scholar] [CrossRef]
- Yang, R.; Fredman, G.; Krishnamoorthy, S.; Agrawal, N.; Irimia, D.; Piomelli, D.; Serhan, C.N. Decoding Functional Metabolomics with Docosahexaenoyl Ethanolamide (DHEA) Identifies Novel Bioactive Signals. J. Biol. Chem. 2011, 286, 31532–31541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rovito, D.; Giordano, C.; Vizza, D.; Plastina, P.; Barone, I.; Casaburi, I.; Lanzino, M.; De Amicis, F.; Sisci, D.; Mauro, L.; et al. Omega-3 PUFA Ethanolamides DHEA and EPEA Induce Autophagy through PPARγ Activation in MCF-7 Breast Cancer Cells. J. Cell. Physiol. 2013, 228, 1314–1322. [Google Scholar] [CrossRef]
- De Bus, I.; Witkamp, R.; Zuilhof, H.; Albada, B.; Balvers, M. The Role of N-3 PUFA-Derived Fatty Acid Derivatives and Their Oxygenated Metabolites in the Modulation of Inflammation. Prostaglandins Lipid Mediat. 2019, 144, 106351. [Google Scholar] [CrossRef]
- Park, T.; Chen, H.; Kevala, K.; Lee, J.-W.; Kim, H.-Y. N-Docosahexaenoylethanolamine Ameliorates LPS-Induced Neuroinflammation via CAMP/PKA-Dependent Signaling. J. Neuroinflamm. 2016, 13, 284. [Google Scholar] [CrossRef] [Green Version]
- Hansen, H.S.; Moesgaard, B.; Hansen, H.H.; Petersen, G. N-Acylethanolamines and Precursor Phospholipids—Relation to Cell Injury. Chem. Phys. Lipids 2000, 108, 135–150. [Google Scholar] [CrossRef]
- Iannotti, F.A.; Di Marzo, V.; Petrosino, S. Endocannabinoids and Endocannabinoid-Related Mediators: Targets, Metabolism and Role in Neurological Disorders. Prog. Lipid Res. 2016, 62, 107–128. [Google Scholar] [CrossRef]
- Wang, J.; Ueda, N. Biology of Endocannabinoid Synthesis System. Prostaglandins Lipid Mediat. 2009, 89, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.Q.; Mikkelsen, T.S.; McKinney, M.K.; Lander, E.S.; Cravatt, B.F. A Second Fatty Acid Amide Hydrolase with Variable Distribution among Placental Mammals. J. Biol. Chem. 2006, 281, 36569–36578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cravatt, B.F.; Demarest, K.; Patricelli, M.P.; Bracey, M.H.; Giang, D.K.; Martin, B.R.; Lichtman, A.H. Supersensitivity to Anandamide and Enhanced Endogenous Cannabinoid Signaling in Mice Lacking Fatty Acid Amide Hydrolase. Proc. Natl. Acad. Sci. USA 2001, 98, 9371–9376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuboi, K.; Takezaki, N.; Ueda, N. TheN-Acylethanolamine-Hydrolyzing Acid Amidase (NAAA). Chem. Biodivers. 2007, 4, 1914–1925. [Google Scholar] [CrossRef] [PubMed]
- Ueda, N.; Tsuboi, K.; Uyama, T. N-Acylethanolamine Metabolism with Special Reference to N-Acylethanolamine-Hydrolyzing Acid Amidase (NAAA). Prog. Lipid Res. 2010, 49, 299–315. [Google Scholar] [CrossRef] [PubMed]
- Costa, B.; Comelli, F.; Bettoni, I.; Colleoni, M.; Giagnoni, G. The Endogenous Fatty Acid Amide, Palmitoylethanolamide, Has Anti-Allodynic and Anti-Hyperalgesic Effects in a Murine Model of Neuropathic Pain: Involvement of CB1, TRPV1 and PPARγ Receptors and Neurotrophic Factors. Pain 2008, 139, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Godlewski, G.; Offertáler, L.; Wagner, J.A.; Kunos, G. Receptors for Acylethanolamides—GPR55 and GPR119. Prostaglandins Lipid Mediat. 2009, 89, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Tsuboi, K.; Uyama, T.; Okamoto, Y.; Ueda, N. Endocannabinoids and Related N-Acylethanolamines: Biological Activities and Metabolism. Inflamm. Regen. 2018, 38, 28. [Google Scholar] [CrossRef]
- Hansen, H.S.; Moesgaard, B.; Petersen, G.; Hansen, H.H. Putative Neuroprotective Actions of N-Acyl-Ethanolamines. Pharmacol. Ther. 2002, 95, 119–126. [Google Scholar] [CrossRef]
- Di Marzo, V.; Melck, D.; Orlando, P.; Bisogno, T.; Zagoory, O.; Bifulco, M.; Vogel, Z.; de Petrocellis, L. Palmitoylethanolamide Inhibits the Expression of Fatty Acid Amide Hydrolase and Enhances the Anti-Proliferative Effect of Anandamide in Human Breast Cancer Cells. Biochem. J. 2001, 358, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, K.O.; Vandevoorde, S.; Lambert, D.M.; Tiger, G.; Fowler, C.J. Effects of Homologues and Analogues of Palmitoylethanolamide upon the Inactivation of the Endocannabinoid Anandamide. Br. J. Pharmcol. 2001, 133, 1263–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertwee, R.G. GPR55: A New Member of the Cannabinoid Receptor Clan?: Commentary. Br. J. Pharmcol. 2007, 152, 984–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, W.-L.; Peng, Y.-Y.; Peng, B.-W. Modulation of Neuroinflammation: Role and Therapeutic Potential of TRPV1 in the Neuro-Immune Axis. Brain Behav. Immun. 2017, 64, 354–366. [Google Scholar] [CrossRef] [PubMed]
- Lo Verme, J.; Fu, J.; Astarita, G.; La Rana, G.; Russo, R.; Calignano, A.; Piomelli, D. The Nuclear Receptor Peroxisome Proliferator-Activated Receptor-α Mediates the Anti-Inflammatory Actions of Palmitoylethanolamide. Mol. Pharm. 2005, 67, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Bento-Abreu, A.; Tabernero, A.; Medina, J.M. Peroxisome Proliferator-Activated Receptor-Alpha Is Required for the Neurotrophic Effect of Oleic Acid in Neurons. J. Neurochem. 2007, 103, 871–881. [Google Scholar] [CrossRef] [PubMed]
- Wasilewski, M.; Więckowski, M.R.; Dymkowska, D.; Wojtczak, L. Effects of N-Acylethanolamines on Mitochondrial Energetics and Permeability Transition. Biochim. Biophys. Acta Bioenerg. 2004, 1657, 151–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zolese, G.; Bacchetti, T.; Masciangelo, S.; Ragni, L.; Ambrosi, S.; Ambrosini, A.; Marini, M.; Ferretti, G. Effect of Acylethanolamides on Lipid Peroxidation and Paraoxonase Activity. Biofactors 2008, 33, 201–209. [Google Scholar] [CrossRef]
- Franklin, A.; Parmentier-Batteur, S.; Walter, L.; Greenberg, D.A.; Stella, N. Palmitoylethanolamide Increases after Focal Cerebral Ischemia and Potentiates Microglial Cell Motility. J. Neurosci. 2003, 23, 7767–7775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galan-Rodriguez, B.; Suarez, J.; Gonzalez-Aparicio, R.; Bermudez-Silva, F.J.; Maldonado, R.; Robledo, P.; Rodriguez de Fonseca, F.; Fernandez-Espejo, E. Oleoylethanolamide Exerts Partial and Dose-Dependent Neuroprotection of Substantia Nigra Dopamine Neurons. Neuropharmacology 2009, 56, 653–664. [Google Scholar] [CrossRef]
- Sayd, A.; Anton, M.; Alen, F.; Caso, J.R.; Pavon, J.; Leza, J.C.; Rodriguez de Fonseca, F.; Garcia-Bueno, B.; Orio, L. Systemic Administration of Oleoylethanolamide Protects from Neuroinflammation and Anhedonia Induced by LPS in Rats. Int. J. Neuropsychopharmacol. 2015, 18, pyu111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCoy, K.L. Interaction between Cannabinoid System and Toll-Like Receptors Controls Inflammation. Mediat. Inflamm. 2016, 2016, 5831315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fidaleo, M.; Fanelli, F.; Ceru, M.; Moreno, S. Neuroprotective Properties of Peroxisome Proliferator-Activated Receptor Alpha (PPARα) and Its Lipid Ligands. Curr. Med. Chem. 2014, 21, 2803–2821. [Google Scholar] [CrossRef] [PubMed]
- Skaper, S.D.; Facci, L.; Barbierato, M.; Zusso, M.; Bruschetta, G.; Impellizzeri, D.; Cuzzocrea, S.; Giusti, P. N-Palmitoylethanolamine and Neuroinflammation: A Novel Therapeutic Strategy of Resolution. Mol. Neurobiol. 2015, 52, 1034–1042. [Google Scholar] [CrossRef]
- Rahimi, A.; Faizi, M.; Talebi, F.; Noorbakhsh, F.; Kahrizi, F.; Naderi, N. Interaction between the Protective Effects of Cannabidiol and Palmitoylethanolamide in Experimental Model of Multiple Sclerosis in C57BL/6 Mice. Neuroscience 2015, 290, 279–287. [Google Scholar] [CrossRef] [PubMed]
- D’Agostino, G.; Russo, R.; Avagliano, C.; Cristiano, C.; Meli, R.; Calignano, A. Palmitoylethanolamide Protects Against the Amyloid-Β25-35-Induced Learning and Memory Impairment in Mice, an Experimental Model of Alzheimer Disease. Neuropsychopharmacology 2012, 37, 1784–1792. [Google Scholar] [CrossRef] [Green Version]
- Esposito, E.; Impellizzeri, D.; Mazzon, E.; Paterniti, I.; Cuzzocrea, S. Neuroprotective Activities of Palmitoylethanolamide in an Animal Model of Parkinson’s Disease. PLoS ONE 2012, 7, e41880. [Google Scholar] [CrossRef] [Green Version]
- Joshi, U.; Evans, J.E.; Joseph, R.; Emmerich, T.; Saltiel, N.; Lungmus, C.; Oberlin, S.; Langlois, H.; Ojo, J.; Mouzon, B.; et al. Oleoylethanolamide Treatment Reduces Neurobehavioral Deficits and Brain Pathology in a Mouse Model of Gulf War Illness. Sci. Rep. 2018, 8, 12921. [Google Scholar] [CrossRef] [Green Version]
- Orio, L.; Alen, F.; Pavón, F.J.; Serrano, A.; García-Bueno, B. Oleoylethanolamide, Neuroinflammation, and Alcohol Abuse. Front. Mol. Neurosci. 2019, 11, 490. [Google Scholar] [CrossRef]
- Diep, T.A.; Madsen, A.N.; Holst, B.; Kristiansen, M.M.; Wellner, N.; Hansen, S.H.; Hansen, H.S. Dietary Fat Decreases Intestinal Levels of the Anorectic Lipids through a Fat Sensor. FASEB J. 2011, 25, 765–774. [Google Scholar] [CrossRef]
- Hansen, S.L.; Nielsen, A.H.; Knudsen, K.E.; Artmann, A.; Petersen, G.; Kristiansen, U.; Hansen, S.H.; Hansen, H.S. Ketogenic Diet Is Antiepileptogenic in Pentylenetetrazole Kindled Mice and Decrease Levels of N-Acylethanolamines in Hippocampus. Neurochem. Int. 2009, 54, 199–204. [Google Scholar] [CrossRef]
- Castonguay-Paradis, S.; Lacroix, S.; Rochefort, G.; Parent, L.; Perron, J.; Martin, C.; Lamarche, B.; Raymond, F.; Flamand, N.; Di Marzo, V.; et al. Dietary Fatty Acid Intake and Gut Microbiota Determine Circulating Endocannabinoidome Signaling beyond the Effect of Body Fat. Sci. Rep. 2020, 10, 15975. [Google Scholar] [CrossRef] [PubMed]
- De Luca, L.; Ferracane, R.; Vitaglione, P. Food Database of N-Acyl-Phosphatidylethanolamines, N-Acylethanolamines and Endocannabinoids and Daily Intake from a Western, a Mediterranean and a Vegetarian Diet. Food Chem. 2019, 300, 125218. [Google Scholar] [CrossRef] [PubMed]
- Silva, P.; Sureda, A.; Tur, J.A.; Andreoletti, P.; Cherkaoui-Malki, M.; Latruffe, N. How Efficient Is Resveratrol as an Antioxidant of the Mediterranean Diet, towards Alterations during the Aging Process? Free Radic. Res. 2019, 53, 1101–1112. [Google Scholar] [CrossRef] [PubMed]
- Walle, T.; Hsieh, F.; DeLegge, M.H.; Oatis, J.E.; Walle, U.K. High Absorption but Very Low Bioavailability of Oral Resveratrol in Humans. Drug Metab. Dispos. 2004, 32, 1377–1382. [Google Scholar] [CrossRef] [Green Version]
- Ditano-Vázquez, P.; Torres-Peña, J.D.; Galeano-Valle, F.; Pérez-Caballero, A.I.; Demelo-Rodríguez, P.; Lopez-Miranda, J.; Katsiki, N.; Delgado-Lista, J.; Alvarez-Sala-Walther, L.A. The Fluid Aspect of the Mediterranean Diet in the Prevention and Management of Cardiovascular Disease and Diabetes: The Role of Polyphenol Content in Moderate Consumption of Wine and Olive Oil. Nutrients 2019, 11, 2833. [Google Scholar] [CrossRef] [Green Version]
- Sun, A.Y.; Wang, Q.; Simonyi, A.; Sun, G.Y. Resveratrol as a Therapeutic Agent for Neurodegenerative Diseases. Mol. Neurobiol. 2010, 41, 375–383. [Google Scholar] [CrossRef] [Green Version]
- Gea, A.; Bes-Rastrollo, M.; Toledo, E.; Garcia-Lopez, M.; Beunza, J.J.; Estruch, R.; Martinez-Gonzalez, M.A. Mediterranean Alcohol-Drinking Pattern and Mortality in the SUN (Seguimiento Universidad de Navarra) Project: A Prospective Cohort Study. Br. J. Nutr. 2014, 111, 1871–1880. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Xu, J.; Rottinghaus, G.E.; Simonyi, A.; Lubahn, D.; Sun, G.Y.; Sun, A.Y. Resveratrol Protects against Global Cerebral Ischemic Injury in Gerbils. Brain Res. 2002, 958, 439–447. [Google Scholar] [CrossRef]
- Milne, J.C.; Denu, J.M. The Sirtuin Family: Therapeutic Targets to Treat Diseases of Aging. Curr. Opin. Chem. Biol. 2008, 12, 11–17. [Google Scholar] [CrossRef]
- Lindsay, J.; Laurin, D.; Verreault, R.; Hébert, R.; Helliwell, B.; Hill, G.B.; McDowell, I. Risk Factors for Alzheimer’s Disease: A Prospective Analysis from the Canadian Study of Health and Aging. Am. J. Epidemiol. 2002, 156, 445–453. [Google Scholar] [CrossRef] [Green Version]
- Albani, D.; Polito, L.; Signorini, A.; Forloni, G. Neuroprotective Properties of Resveratrol in Different Neurodegenerative Disorders. BioFactors 2010, 36, 370–376. [Google Scholar] [CrossRef]
- Brents, L.K.; Medina-Bolivar, F.; Seely, K.A.; Nair, V.; Bratton, S.M.; Ñopo-Olazabal, L.; Patel, R.Y.; Liu, H.; Doerksen, R.J.; Prather, P.L.; et al. Natural Prenylated Resveratrol Analogs Arachidin-1 and -3 Demonstrate Improved Glucuronidation Profiles and Have Affinity for Cannabinoid Receptors. Xenobiotica 2012, 42, 139–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassanzadeh, P.; Arbabi, E.; Atyabi, F.; Dinarvand, R. The Endocannabinoid System and NGF Are Involved in the Mechanism of Action of Resveratrol: A Multi-Target Nutraceutical with Therapeutic Potential in Neuropsychiatric Disorders. Psychopharmacology 2016, 233, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Allen, S.J.; Watson, J.J.; Shoemark, D.K.; Barua, N.U.; Patel, N.K. GDNF, NGF and BDNF as Therapeutic Options for Neurodegeneration. Pharmacol. Ther. 2013, 138, 155–175. [Google Scholar] [CrossRef] [PubMed]
- Conner, J.M.; Franks, K.M.; Titterness, A.K.; Russell, K.; Merrill, D.A.; Christie, B.R.; Sejnowski, T.J.; Tuszynski, M.H. NGF Is Essential for Hippocampal Plasticity and Learning. J. Neurosci. 2009, 29, 10883–10889. [Google Scholar] [CrossRef]
- Faustino, C.; Rijo, P.; Reis, C.P. Nanotechnological Strategies for Nerve Growth Factor Delivery: Therapeutic Implications in Alzheimer’s Disease. Pharm. Res. 2017, 120, 68–87. [Google Scholar] [CrossRef]
- Oliveira, C.D.C.; e Castor, M.G.M.; e Castor, C.G.M.; Costa, Á.D.F.; Ferreira, R.C.M.; da Silva, J.F.; Pelaez, J.M.N.; Capettini, L.D.S.A.; Lemos, V.S.; Duarte, I.D.G.; et al. Evidence for the Involvement of Opioid and Cannabinoid Systems in the Peripheral Antinociception Mediated by Resveratrol. Toxicol. Appl. Pharmacol. 2019, 369, 30–38. [Google Scholar] [CrossRef]
- Carta, G.; Poddighe, L.; Serra, M.; Boi, M.; Melis, T.; Lisai, S.; Murru, E.; Muredda, L.; Collu, M.; Banni, S.; et al. Preventive Effects of Resveratrol on Endocannabinoid System and Synaptic Protein Modifications in Rat Cerebral Cortex Challenged by Bilateral Common Carotid Artery Occlusion and Reperfusion. Int. J. Mol. Sci. 2018, 19, 426. [Google Scholar] [CrossRef] [Green Version]
- Sundaresan, A.; Radhiga, T.; Deivasigamani, B. Biological Activity of Biochanin A: A Review. Asian J. Pharm. Pharm. 2018, 4, 1–5. [Google Scholar] [CrossRef]
- McDougle, D.R.; Watson, J.E.; Abdeen, A.A.; Adili, R.; Caputo, M.P.; Krapf, J.E.; Johnson, R.W.; Kilian, K.A.; Holinstat, M.; Das, A. Anti-Inflammatory ω-3 Endocannabinoid Epoxides. Proc. Natl. Acad. Sci. USA 2017, 114, E6034–E6043. [Google Scholar] [CrossRef] [Green Version]
- Yurko-Mauro, K.; Alexander, D.D.; Van Elswyk, M.E. Docosahexaenoic Acid and Adult Memory: A Systematic Review and Meta-Analysis. PLoS ONE 2015, 10, e0120391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Wurff, I.S.M.; Meyer, B.J.; de Groot, R.H.M. Effect of Omega-3 Long Chain Polyunsaturated Fatty Acids (n-3 LCPUFA) Supplementation on Cognition in Children and Adolescents: A Systematic Literature Review with a Focus on n-3 LCPUFA Blood Values and Dose of DHA and EPA. Nutrients 2020, 12, 3115. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.P.; Behm, B.; Mehta, Z.; Fernandez, C. The Potential Benefits of Palmitoylethanolamide in Palliation: A Qualitative Systematic Review. Am. J. Hosp. Palliat. Care 2019, 36, 1134–1154. [Google Scholar] [CrossRef] [PubMed]
- Payahoo, L.; Kahjebishak, Y.; Asghari Jafarabadi, M.; Ostadrahimi, A. Oleoylethanolamide Supplementation Reduces Inflammation and Oxidative Stress in Obese People: A Clinical Trial. Adv. Pharm. Bull. 2018, 8, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Pagotto, U.; Vicennati, V.; Pasquali, R. Endocannabinoid system and energy metabolism: Physiology and pathophysiology. G Ital. Cardiol. 2008, 9, 74S–82S. [Google Scholar]
- He, M.; Shi, B. Gut Microbiota as a Potential Target of Metabolic Syndrome: The Role of Probiotics and Prebiotics. Cell Biosci. 2017, 7, 54. [Google Scholar] [CrossRef] [Green Version]
- Bensaid, M.; Gary-Bobo, M.; Esclangon, A.; Maffrand, J.P.; Le Fur, G.; Oury-Donat, F.; Soubrié, P. The Cannabinoid CB1 Receptor Antagonist SR141716 Increases Acrp30 MRNA Expression in Adipose Tissue of Obese Fa/Fa Rats and in Cultured Adipocyte Cells. Mol. Pharm. 2003, 63, 908–914. [Google Scholar] [CrossRef] [Green Version]
- Ravinet Trillou, C.; Arnone, M.; Delgorge, C.; Gonalons, N.; Keane, P.; Maffrand, J.-P.; Soubrié, P. Anti-Obesity Effect of SR141716, a CB1 Receptor Antagonist, in Diet-Induced Obese Mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 284, R345–R353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteleone, P.; Matias, I.; Martiadis, V.; De Petrocellis, L.; Maj, M.; Di Marzo, V. Blood Levels of the Endocannabinoid Anandamide Are Increased in Anorexia Nervosa and in Binge-Eating Disorder, but Not in Bulimia Nervosa. Neuropsychopharmacology 2005, 30, 1216–1221. [Google Scholar] [CrossRef] [Green Version]
- Ströhle, A.; Hahn, A. Diets of Modern Hunter-Gatherers Vary Substantially in Their Carbohydrate Content Depending on Ecoenvironments: Results from an Ethnographic Analysis. Nutr. Res. 2011, 31, 429–435. [Google Scholar] [CrossRef]
- Mazier, W.; Saucisse, N.; Gatta-Cherifi, B.; Cota, D. The Endocannabinoid System: Pivotal Orchestrator of Obesity and Metabolic Disease. Trends Endocrinol. Metab. 2015, 26, 524–537. [Google Scholar] [CrossRef]
- Di Marzo, V.; Goparaju, S.K.; Wang, L.; Liu, J.; Bátkai, S.; Járai, Z.; Fezza, F.; Miura, G.I.; Palmiter, R.D.; Sugiura, T.; et al. Leptin-Regulated Endocannabinoids Are Involved in Maintaining Food Intake. Nature 2001, 410, 822–825. [Google Scholar] [CrossRef] [PubMed]
- Burhans, M.S.; Hagman, D.K.; Kuzma, J.N.; Schmidt, K.A.; Kratz, M. Contribution of Adipose Tissue Inflammation to the Development of Type 2 Diabetes Mellitus. Compr. Physiol. 2018, 9, 1–58. [Google Scholar] [CrossRef] [PubMed]
- Knani, I.; Earley, B.J.; Udi, S.; Nemirovski, A.; Hadar, R.; Gammal, A.; Cinar, R.; Hirsch, H.J.; Pollak, Y.; Gross, I.; et al. Targeting the Endocannabinoid/CB1 Receptor System for Treating Obesity in Prader–Willi Syndrome. Mol. Metab. 2016, 5, 1187–1199. [Google Scholar] [CrossRef] [PubMed]
- Alvheim, A.R.; Malde, M.K.; Osei-Hyiaman, D.; Hong, Y.H.; Pawlosky, R.J.; Madsen, L.; Kristiansen, K.; Frøyland, L.; Hibbeln, J.R. Dietary Linoleic Acid Elevates Endogenous 2-AG and Anandamide and Induces Obesity. Obesity 2012, 20, 1984–1994. [Google Scholar] [CrossRef] [Green Version]
- Rousseaux, C.; Thuru, X.; Gelot, A.; Barnich, N.; Neut, C.; Dubuquoy, L.; Dubuquoy, C.; Merour, E.; Geboes, K.; Chamaillard, M.; et al. Lactobacillus Acidophilus Modulates Intestinal Pain and Induces Opioid and Cannabinoid Receptors. Nat. Med. 2007, 13, 35–37. [Google Scholar] [CrossRef]
- Muccioli, G.G.; Naslain, D.; Bäckhed, F.; Reigstad, C.S.; Lambert, D.M.; Delzenne, N.M.; Cani, P.D. The Endocannabinoid System Links Gut Microbiota to Adipogenesis. Mol. Syst. Biol. 2010, 6, 392. [Google Scholar] [CrossRef]
- Di Marzo, V.; Silvestri, C. Lifestyle and Metabolic Syndrome: Contribution of the Endocannabinoidome. Nutrients 2019, 11, 1956. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Bátkai, S.; Pacher, P.; Harvey-White, J.; Wagner, J.A.; Cravatt, B.F.; Gao, B.; Kunos, G. Lipopolysaccharide Induces Anandamide Synthesis in Macrophages via CD14/MAPK/Phosphoinositide 3-Kinase/NF-ΚB Independently of Platelet-Activating Factor. J. Biol. Chem. 2003, 278, 45034–45039. [Google Scholar] [CrossRef] [Green Version]
- Forte, N.; Fernández-Rilo, A.C.; Palomba, L.; Di Marzo, V.; Cristino, L. Obesity Affects the Microbiota-Gut-Brain Axis and the Regulation Thereof by Endocannabinoids and Related Mediators. Int. J. Mol. Sci. 2020, 21, 1554. [Google Scholar] [CrossRef] [Green Version]
- Iannotti, F.A.; Di Marzo, V. The Gut Microbiome, Endocannabinoids and Metabolic Disorders. J. Endocrinol. 2021, 248, R83–R97. [Google Scholar] [CrossRef]
- Mehrpouya-Bahrami, P.; Chitrala, K.N.; Ganewatta, M.S.; Tang, C.; Murphy, E.A.; Enos, R.T.; Velazquez, K.T.; McCellan, J.; Nagarkatti, M.; Nagarkatti, P. Blockade of CB1 Cannabinoid Receptor Alters Gut Microbiota and Attenuates Inflammation and Diet-Induced Obesity. Sci. Rep. 2017, 7, 15645. [Google Scholar] [CrossRef]
- Leuti, A.; Fazio, D.; Fava, M.; Piccoli, A.; Oddi, S.; Maccarrone, M. Bioactive Lipids, Inflammation and Chronic Diseases. Adv. Drug Deliv. Rev. 2020, 159, 133–169. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Bátkai, S.; Kunos, G. The Endocannabinoid System as an Emerging Target of Pharmacotherapy. Pharm. Rev. 2006, 58, 389–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinz, B.; Ramer, R. Anti-Tumour Actions of Cannabinoids. Br. J. Pharmcol. 2019, 176, 1384–1394. [Google Scholar] [CrossRef] [PubMed]
- Chaplin, A.; Carpéné, C.; Mercader, J. Resveratrol, Metabolic Syndrome, and Gut Microbiota. Nutrients 2018, 10, 1651. [Google Scholar] [CrossRef] [Green Version]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, H.A.; Lerin, C.; Kalra, A.; Prabhu, V.V.; Allard, J.S.; Lopez-Lluch, G.; Lewis, K.; et al. Resveratrol Improves Health and Survival of Mice on a High-Calorie Diet. Nature 2006, 444, 337–342. [Google Scholar] [CrossRef]
- Macarulla, M.T.; Alberdi, G.; Gómez, S.; Tueros, I.; Bald, C.; Rodríguez, V.M.; Martínez, J.A.; Portillo, M.P. Effects of Different Doses of Resveratrol on Body Fat and Serum Parameters in Rats Fed a Hypercaloric Diet. J. Physiol. Biochem. 2009, 65, 369–376. [Google Scholar] [CrossRef]
- Kim, S.; Jin, Y.; Choi, Y.; Park, T. Resveratrol Exerts Anti-Obesity Effects via Mechanisms Involving down-Regulation of Adipogenic and Inflammatory Processes in Mice. Biochem. Pharmacol. 2011, 81, 1343–1351. [Google Scholar] [CrossRef]
- Picard, F.; Kurtev, M.; Chung, N.; Topark-Ngarm, A.; Senawong, T.; Machado de Oliveira, R.; Leid, M.; McBurney, M.W.; Guarente, L. Sirt1 Promotes Fat Mobilization in White Adipocytes by Repressing PPAR-γ. Nature 2004, 429, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.; Hong, H.J.; Guan, J.; Kim, D.G.; Yang, E.-J.; Koh, G.; Park, D.; Han, C.H.; Lee, Y.-J.; Lee, D.-H. Resveratrol Improves Insulin Signaling in a Tissue-Specific Manner under Insulin-Resistant Conditions Only: In Vitro and in Vivo Experiments in Rodents. Metabolism 2012, 61, 424–433. [Google Scholar] [CrossRef]
- Springer, M.; Moco, S. Resveratrol and Its Human Metabolites—Effects on Metabolic Health and Obesity. Nutrients 2019, 11, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dvorakova, M.; Landa, P. Anti-Inflammatory Activity of Natural Stilbenoids: A Review. Pharm. Res. 2017, 124, 126–145. [Google Scholar] [CrossRef] [PubMed]
- Martín, A.R.; Villegas, I.; La Casa, C.; de la Lastra, C.A. Resveratrol, a Polyphenol Found in Grapes, Suppresses Oxidative Damage and Stimulates Apoptosis during Early Colonic Inflammation in Rats. Biochem. Pharm. 2004, 67, 1399–1410. [Google Scholar] [CrossRef]
- Ølholm, J.; Paulsen, S.K.; Cullberg, K.B.; Richelsen, B.; Pedersen, S.B. Anti-Inflammatory Effect of Resveratrol on Adipokine Expression and Secretion in Human Adipose Tissue Explants. Int. J. Obes. 2010, 34, 1546–1553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yen, G.-C.; Chen, Y.-C.; Chang, W.-T.; Hsu, C.-L. Effects of Polyphenolic Compounds on Tumor Necrosis Factor-α (TNF-α)-Induced Changes of Adipokines and Oxidative Stress in 3T3-L1 Adipocytes. J. Agric. Food Chem. 2011, 59, 546–551. [Google Scholar] [CrossRef] [PubMed]
- Ghanim, H.; Sia, C.L.; Korzeniewski, K.; Lohano, T.; Abuaysheh, S.; Marumganti, A.; Chaudhuri, A.; Dandona, P. A Resveratrol and Polyphenol Preparation Suppresses Oxidative and Inflammatory Stress Response to a High-Fat, High-Carbohydrate Meal. J. Clin. Endocrinol. Metab. 2011, 96, 1409–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomé-Carneiro, J.; Gonzálvez, M.; Larrosa, M.; Yáñez-Gascón, M.J.; García-Almagro, F.J.; Ruiz-Ros, J.A.; García-Conesa, M.T.; Tomás-Barberán, F.A.; Espín, J.C. One-Year Consumption of a Grape Nutraceutical Containing Resveratrol Improves the Inflammatory and Fibrinolytic Status of Patients in Primary Prevention of Cardiovascular Disease. Am. J. Cardiol. 2012, 110, 356–363. [Google Scholar] [CrossRef]
- Chen, M.; Hou, P.; Zhou, M.; Ren, Q.; Wang, X.; Huang, L.; Hui, S.; Yi, L.; Mi, M. Resveratrol Attenuates High-Fat Diet-Induced Non-Alcoholic Steatohepatitis by Maintaining Gut Barrier Integrity and Inhibiting Gut Inflammation through Regulation of the Endocannabinoid System. Clin. Nutr. 2020, 39, 1264–1275. [Google Scholar] [CrossRef]
- Sikand, G.; Kris-Etherton, P.; Boulos, N.M. Impact of Functional Foods on Prevention of Cardiovascular Disease and Diabetes. Curr. Cardiol. Rep. 2015, 17, 39. [Google Scholar] [CrossRef]
- Medina-Gomez, G.; Gray, S.; Vidal-Puig, A. Adipogenesis and Lipotoxicity: Role of Peroxisome Proliferator-Activated Receptor γ (PPARγ) and PPARγcoactivator-1 (PGC1). Public Health Nutr. 2007, 10, 1132–1137. [Google Scholar] [CrossRef]
- Wang, L.; Waltenberger, B.; Pferschy-Wenzig, E.-M.; Blunder, M.; Liu, X.; Malainer, C.; Blazevic, T.; Schwaiger, S.; Rollinger, J.M.; Heiss, E.H.; et al. Natural Product Agonists of Peroxisome Proliferator-Activated Receptor Gamma (PPARγ): A Review. Biochem. Pharm. 2014, 92, 73–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bento, A.F.; Marcon, R.; Dutra, R.C.; Claudino, R.F.; Cola, M.; Pereira Leite, D.F.; Calixto, J.B. β-Caryophyllene Inhibits Dextran Sulfate Sodium-Induced Colitis in Mice through CB2 Receptor Activation and PPARγ Pathway. Am. J. Pathol. 2011, 178, 1153–1166. [Google Scholar] [CrossRef]
- Basha, R.H.; Sankaranarayanan, C. β-Caryophyllene, a Natural Sesquiterpene Lactone Attenuates Hyperglycemia Mediated Oxidative and Inflammatory Stress in Experimental Diabetic Rats. Chem. Biol. Interact. 2016, 245, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Basha, R.H.; Sankaranarayanan, C. β-Caryophyllene, a Natural Sesquiterpene, Modulates Carbohydrate Metabolism in Streptozotocin-Induced Diabetic Rats. Acta Histochem. 2014, 116, 1469–1479. [Google Scholar] [CrossRef] [PubMed]
- Leonti, M. The Co-Evolutionary Perspective of the Food-Medicine Continuum and Wild Gathered and Cultivated Vegetables. Genet. Resour. Crop. Evol. 2012, 59, 1295–1302. [Google Scholar] [CrossRef]
- Klauke, A.-L.; Racz, I.; Pradier, B.; Markert, A.; Zimmer, A.M.; Gertsch, J.; Zimmer, A. The Cannabinoid CB2 Receptor-Selective Phytocannabinoid Beta-Caryophyllene Exerts Analgesic Effects in Mouse Models of Inflammatory and Neuropathic Pain. Eur. Neuropsychopharmacology 2014, 24, 608–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Notarnicola, M.; Tutino, V.; Tafaro, A.; Bianco, G.; Guglielmi, E.; Caruso, M.G. Dietary Olive Oil Induces Cannabinoid CB2 Receptor Expression in Adipose Tissue of ApcMin/+ Transgenic Mice. Nutr. Health Aging 2016, 4, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Simopoulos, A. An Increase in the Omega-6/Omega-3 Fatty Acid Ratio Increases the Risk for Obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef] [Green Version]
- Caracciolo, B.; Xu, W.; Collins, S.; Fratiglioni, L. Cognitive Decline, Dietary Factors and Gut–Brain Interactions. Mech. Ageing Dev. 2014, 136–137, 59–69. [Google Scholar] [CrossRef]
- Di Marzo, V. ‘Endocannabinoids’ and Other Fatty Acid Derivatives with Cannabimimetic Properties: Biochemistry and Possible Physiopathological Relevance. Biochim. Biophys. Acta Lipids Lipid. Metab. 1998, 1392, 153–175. [Google Scholar] [CrossRef]
- Russo, E.B. Clinical Endocannabinoid Deficiency Reconsidered: Current Research Supports the Theory in Migraine, Fibromyalgia, Irritable Bowel, and Other Treatment-Resistant Syndromes. Cannabis Cannabinoid Res. 2016, 1, 154–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McPartland, J.M.; Glass, M.; Pertwee, R.G. Meta-Analysis of Cannabinoid Ligand Binding Affinity and Receptor Distribution: Interspecies Differences: Ligand Affinity and Receptor Distribution of CB1. Br. J. Pharmcol. 2007, 152, 583–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
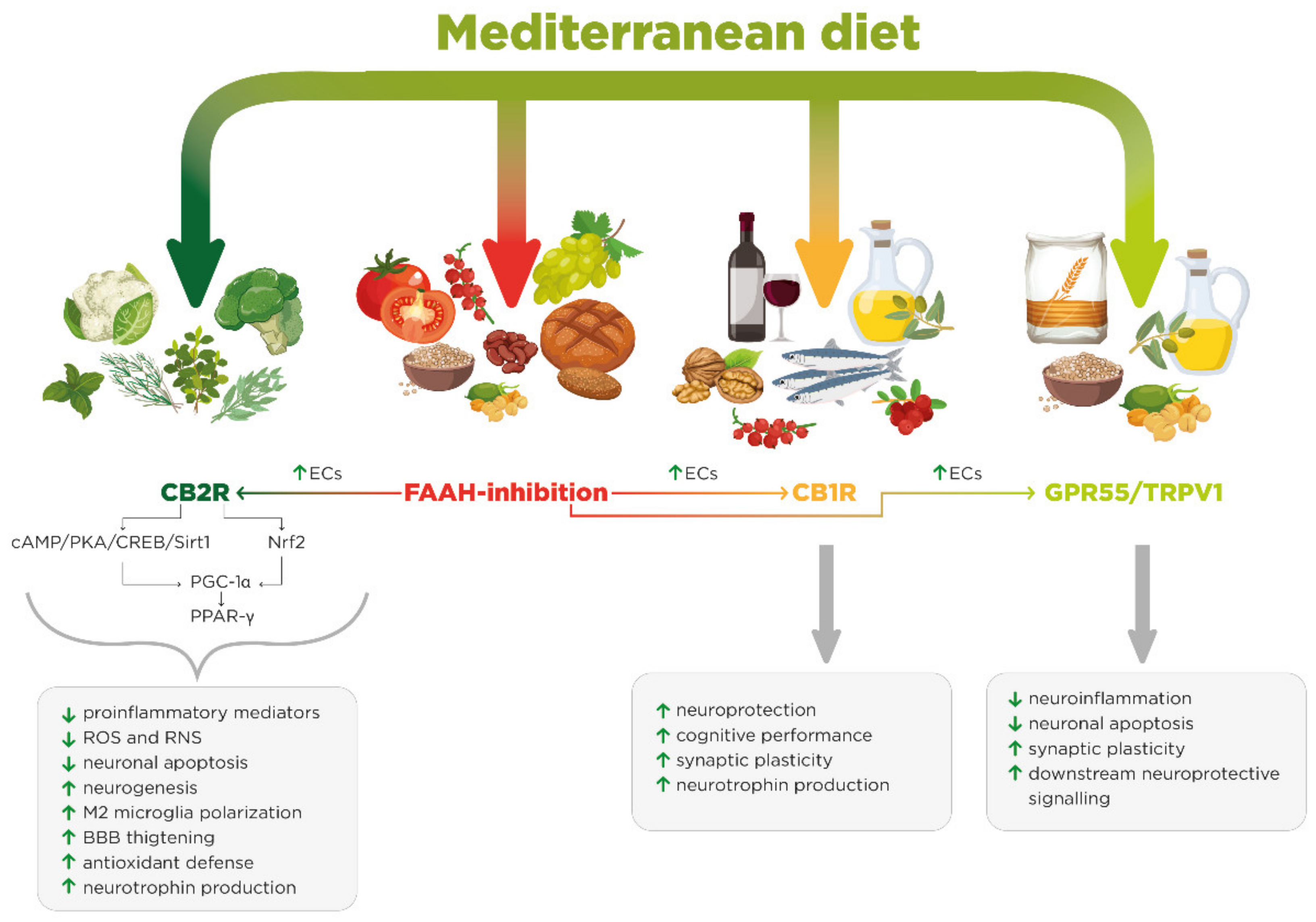{kind=link}
{kind=link}
| Class of Molecules | Compound | Dietary Origin | Endocannabinoid Target/Effects | Potency | Proposed Oral Dose in Humans |
|---|---|---|---|---|---|
| Bicyclic sesquiterpene | β-caryophyllene (BCP) | Black pepper, hops, oregano, cloves, valerian, wild sage, rosemary, basil and cow’s milk | CB2R full agonist | 100 nM [196] | 4 mg/kg/day [196] |
| Glucosinolates (GLs) | 3,3′-diindolylmethane (DIM) | Broccoli, cabbage, cauliflower and horseradish | CB2R partial agonist | 1 μM (binding affinity) [161] | 0.8–1.6 mg/K [149,150] |
| Flavonoids | Biochanin A (isoflavone) | Peanuts and chickpeas | FAAH inhibitor | 2.4 μM (IC50) [176] | 50 mg/kg (rats) [281] |
| Genistein (isoflavone) | Whole grain bread, beans, lentils, chickpeas, split chicklings and fava beans | FAAH competitive inhibitor/PPAR-γ agonist | 2.8 μM (Ki value) [182] | 9.0 mg/day [189]( based on an Asian population) | |
| Daidzein (isoflavone) | FAAH competitive inhibitor/PPAR-γ agonist | 1.7 μM (Ki value) [182] | 7.4 mg/day [189] (based on an Asian population) | ||
| Apigenin (flavons) | Parsley, onions, celery, chamomile, oregano, thyme, basil, oranges, tea, beer and wine | FAAH competitive inhibitor/PPAR-γ agonist/COX-2 inhibitor | 15 μM (IC50) [166] | 0.7 mg/kg [191] | |
| Kaempferol (flavonol) | Leafy vegetables, apples, onions, broccoli, berries, tea, cabbage, broccoli, endive, kale, beans, tomato, strawberries, leek and grapes | Competitive FAAH inhibitor/PPAR-γ agonist | 5 μM (Ki value) [166] | 1.4–5.7 mg/kg/day [195] | |
| ω-3 PUFAs | DHA/EPA | Walnuts, purslane, legumes, green leafy vegetables, olive oil, salmon, trout, mackerel, herring and sardines | ↑ CBRs NAPE-PLD, DAGLα and DAGβ expression (DHA/EPA); CBRs/PPAR-γ partial agonists (DHEA/EPEA); GPR110 agonist (DHEA) | DHEA → CB1 1044 nM (EC50), CB2 305 nM (EC50); EPEA → CB1 0.1 nM (EC50), CB2 2.1 nM (EC50) [282] | >1 g/day DHA/EPA [283,284] |
| N-acylethanolamines (NAEs) | Palmitoylethanolamine (PEA) | Refined wheat flour, beans, lentils, chickpeas and extra-virgin olive oil | Competitive FAAH/NAAA inhibitor, TRPV1/PPAR- α/GPR55 agonist | / | 600–1200 mg/day [285] |
| Oleoylethanolamide (OEA) | Competitive FAAH/NAAA inhibitor, TRPV1/PPAR-α agonist | / | 125 mg/day [286] | ||
| Polyphenols | Resveratrol (stilbene) | Red grapes, red wine, peanuts, blackberries, blackcurrants, blueberries and cranberries | ↑ DHA and PEA levels; ↑ CBRs and PPAR-α expression; CB1R-mediated neuroprotection | / | 1.4–4.2 mg/kg/day [195] |
| Disorders | Disease Models | Compound | Effects | |
|---|---|---|---|---|
| AD | Transgenic APP/PS1 mice | BCP | ↑ Cognitive performance ↓ Aβ accumulation ↓ Astrogliosis ↓Pro-inflammatory mediators | [96] |
| AD | ICR mice (scopolamine) | DIM | ↑ BDNF ↑ Antioxidant enzyme expression ↑ Cognitive performance ↓ Neuroapoptosis | [150] |
| AD | Transgenic APP/PS1 mice | Apigenin | ↑ Neuroprotection | [191] |
| AD | Tg 2576 mice | RSV | ↑ Cognitive performance ↓ Amyloid brain pathology | [273] |
| AD | Mouse model p25 | RSV | ↓ Hippocampal neurodegeneration ↑ Cognitive performance, ↑ SIRT1 overexpression | [273] |
| AD | TgSwDI mice | EVOO (OEA) | ↓ Aβ and neurofibrillary tangles ↑ Cognitive performance BBB Aβ clearance | [76] |
| AD | C57BL6 mice (Aβ 25–35 intracerebroventricular injection) | PEA | ↓ Neuroapoptosis ↓ Neuroinflammation ↓ Memory dysfunction ↓ OxS | [257] |
| PD | Wistar rats (rotenone-induced toxicity) | BCP | ↓ Neuroapoptosis ↓ iNOS), COX-2, TNFα, IL-6 and IL- 1β expression ↑ Antioxidant defenses | [97] |
| PD | C57BL/6J mice (MPTP) | DIM | ↓ Neuroapoptosis ↑ Cytoprotection | [156] |
| PD | Wistar rats (6-OHDA) | OEA | ↑ Neuroprotection | [251] |
| PD | PPAR-αKO and PPAR-αWT mice (MPTP) | PEA | ↓ Microglial activation ↓ Microtube alteration ↓ Neuroapoptosis ↑ Motor behavior | [258] |
| MS | EAE mouse model | BCP | ↓ Axonal demyelination ↓ Microglial cells, CD4+ and CD8+ activity ↑ Th1/Treg immune balance | [98] |
| MS | EAE mouse | PEA | ↓ Demyelination ↓ Neuronal damage ↑ Clinical phenotype | [256] |
| MS | EAE mouse model | BCP | ↓ NO and H2O2 production | [112] |
| ALS | Mouse model of ALS | RSV | ↑ Neuroprotection | [273] |
| Neuropsychiatric diseases | C57BL6/J mice (ω-3 deficiency) | ω-3 | ↑ Anxiolytic effect | [219] |
| Obesity | ω-3 deficiency | ω-3 | ↑ CBR-Gi/o coupling↑ Mood modulation | [220] |
| Obesity | Obese mice | ω-3 | ↑ AEA/2-AG balance | [5] |
| Memory impairment | P18–P24 CD1 mice (ω-3 deficiency) | ω-3 | ↑ Hippocampal LTP ↑ Synaptic plasticity ↓ Hippocampal LTP | [221] |
| Brain aging | Old Wistar rats
(EPA/DHA supplementation) | EPA/DHA | ↓ Neuroapoptosis ↑ Neuroprotection ↑ PPAR-γ expression | [225] |
| LPS-induced neuroinflammation | C57BL/6J mice | DHEA | ↑ cAMP/PKA signaling ↑ Inhibition of NF-κB activation ↓ Pro-inflammatory mediators | [230] |
| LPS-induced neuroinflammation | Wistar Hannover rats | OEA/PEA | ↓ Oxs ↑ NF-κB inhibition ↓ Pro-inflammatory mediators | [252] |
| LPS-induced neuroinflammation | C57BL/6 mice | DIM | ↓ iNOS and COX-2 expression ↑ NF-κB inhibition | [152] |
| Neuropathic pain | C57BL/6 mice (formalin) | BCA | ↓ ERK phosphorylation ↓ Pain | [176] |
| Neuropathic pain | Swiss mice | RSV | ↓ Pain ↑ Neuroprotection | [279] |
| Brain atrophy | Wistar rats | RSV | ↑ NGF | [275] |
| Occlusion-reperfusion stress | Wistar rats | RSV | ↓ OxS ↓ Neuroapoptosis ↑ DHA and PEA levels ↑ CBR and PPAR-α expression ↑ Synaptic plasticity | [280] |
| Stress-related disorders | Wistar rats (contextual fear conditioning) | Kmp | ↑ Anxiolytic effect | [201] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Armeli, F.; Bonucci, A.; Maggi, E.; Pinto, A.; Businaro, R. Mediterranean Diet and Neurodegenerative Diseases: The Neglected Role of Nutrition in the Modulation of the Endocannabinoid System. Biomolecules 2021, 11, 790. https://doi.org/10.3390/biom11060790
Armeli F, Bonucci A, Maggi E, Pinto A, Businaro R. Mediterranean Diet and Neurodegenerative Diseases: The Neglected Role of Nutrition in the Modulation of the Endocannabinoid System. Biomolecules. 2021; 11(6):790. https://doi.org/10.3390/biom11060790
Chicago/Turabian StyleArmeli, Federica, Alessio Bonucci, Elisa Maggi, Alessandro Pinto, and Rita Businaro. 2021. "Mediterranean Diet and Neurodegenerative Diseases: The Neglected Role of Nutrition in the Modulation of the Endocannabinoid System" Biomolecules 11, no. 6: 790. https://doi.org/10.3390/biom11060790