
Cardamonin Attenuates Experimental Colitis and Associated Colorectal Cancer

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Mice
2.3. Cell Lines and Cell Culture
2.4. Cytotoxicity by MTT Assay
2.5. NO measurement by Griess Reagent
2.6. Real-Time PCR
2.7. Collection of Condition Media after Cardamonin Treatment
2.8. Immunofluorescence
2.9. In Vivo Colitis Model and Cardamonin Treatment
2.10. In Vivo Colitis-Associated Colorectal Cancer Model (CAC) and Cardamonin Treatment
2.11. RNA Isolation from Tumor Samples
2.12. MicroRNA Microarray of Tumor Samples and Analysis
2.13. Computational Analysis for Target Prediction
2.14. Statistical Analysis
3. Results
3.1. Cardamonin Did Not Affect the Cell Viability but Inhibited Nitric Oxide (NO) Release in Lipopolysaccharide (LPS) Stimulated RAW 264.7 Cells
3.2. Cardamonin Inhibited the Inflammatory Gene Expression
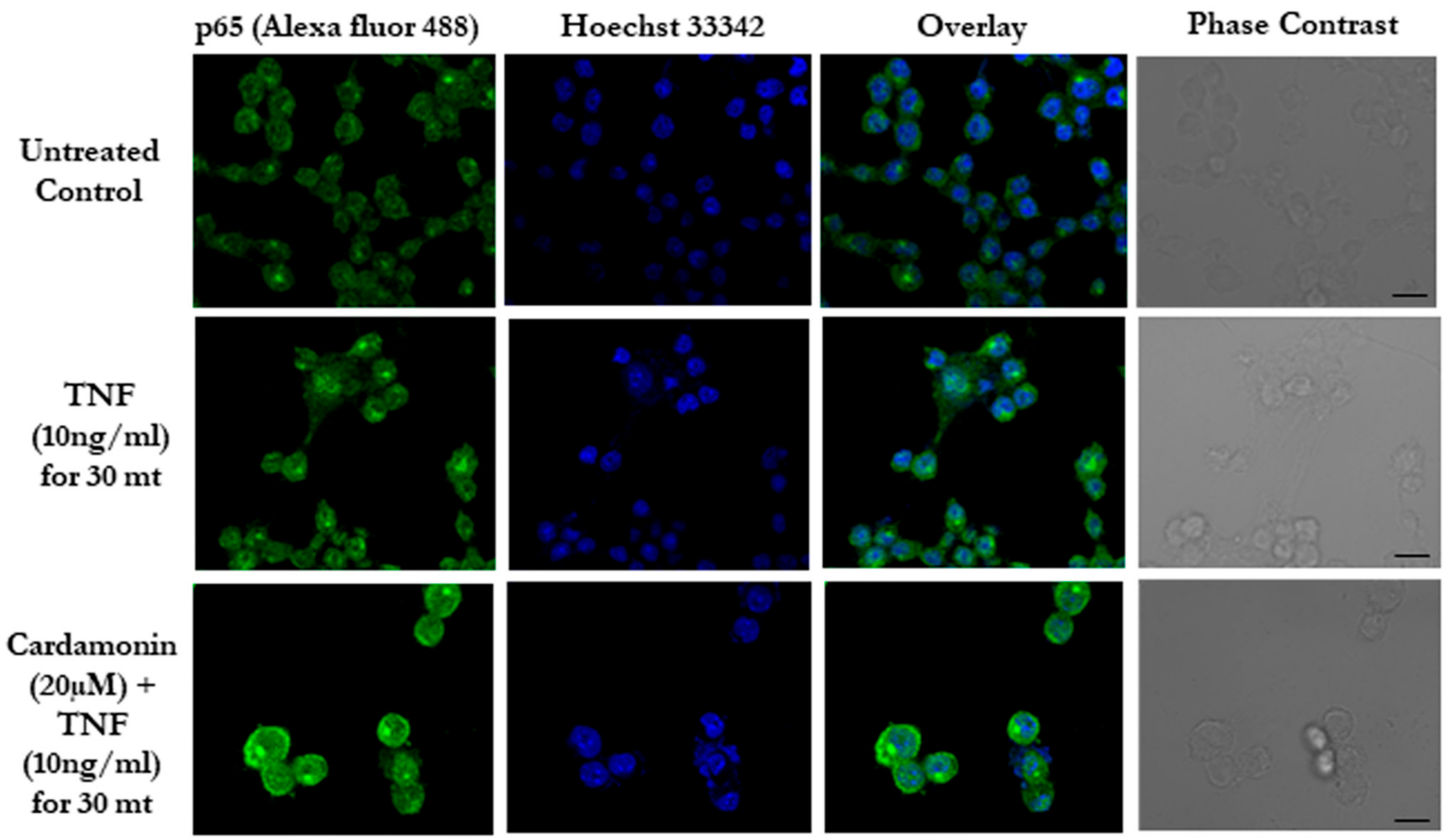
3.3. Cardamonin Blocks the NF-κB Nuclear Translocation
3.4. Cardamonin Attenuated the Colitis Elicited by DSS
3.5. Cardamonin Reduced the Tumor Burden in the Colitis-Associated Colon Cancer
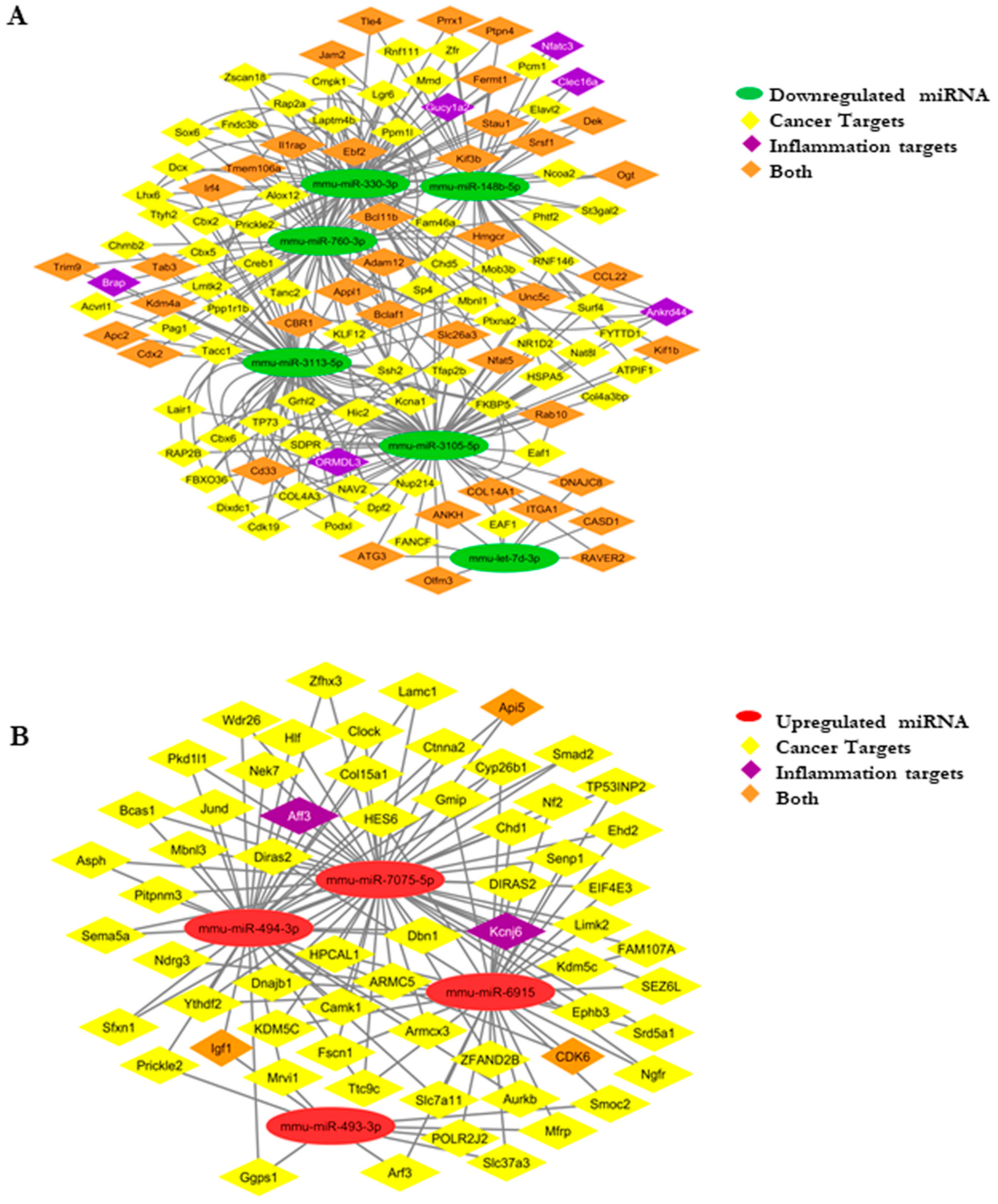
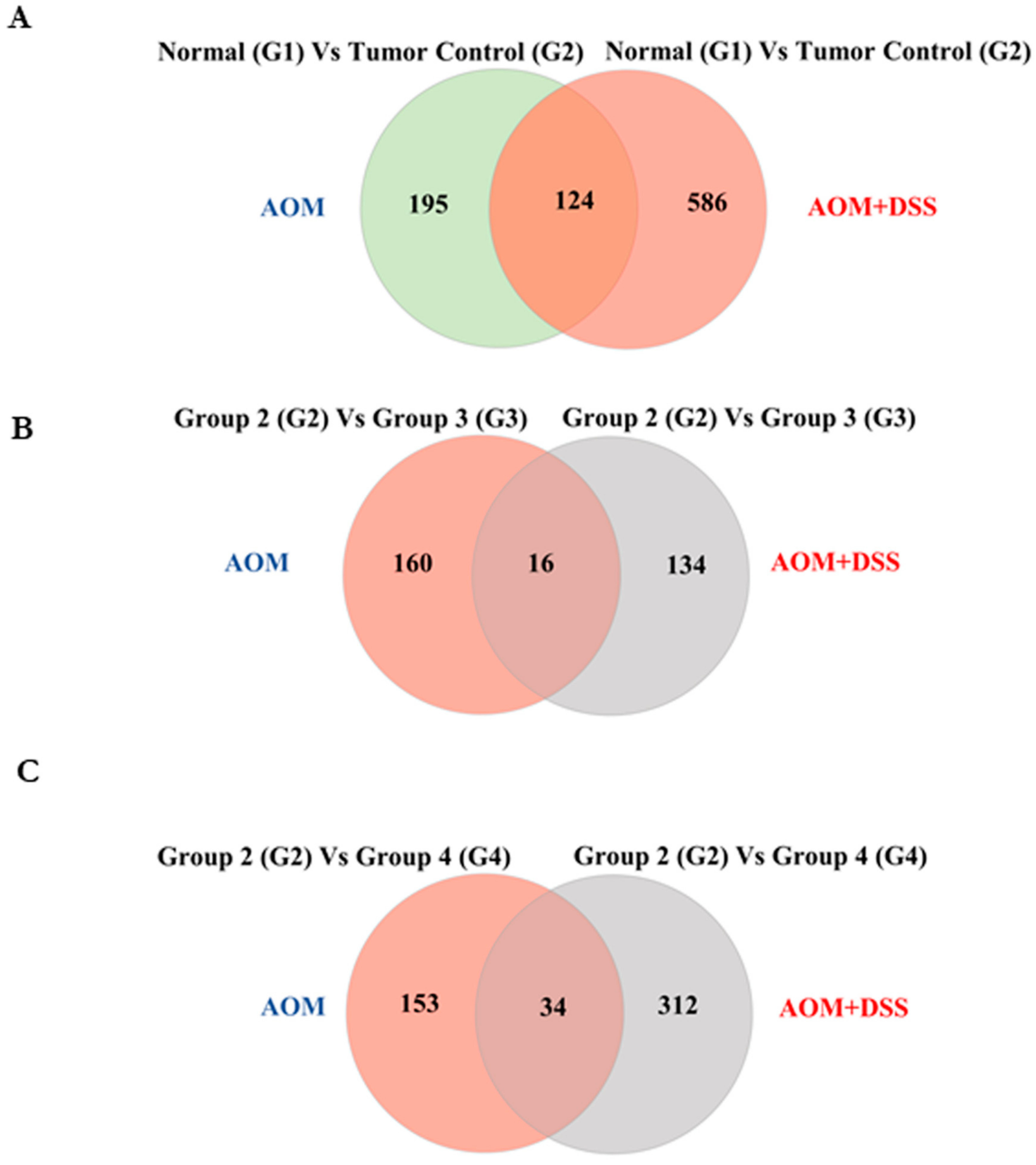
3.6. Cardamonin Altered the miRNA Expression
3.7. Validation of miRNA Microarray Data Using Their Targets
3.8. Developing an miRNA–mRNA Gene Interaction Network
3.9. Cardamonin Altered Crucial Pathways Involved in Cancer and Inflammation
3.10. Predicted Signaling Pathways Altered by miRNA Targets
3.11. Cardamonin Altered miRNA Expression and Regulated the Crucial Molecules in the Cellular Pathways
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Torre, L.A.; Soerjomataram, I.; Hayes, R.B.; Bray, F.; Weber, T.K.; Jemal, A. Global patterns and trends in colorectal cancer incidence in young adults. Gut 2019, 68, 2179–2185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nebbia, M.; Yassin, N.A.; Spinelli, A. Colorectal Cancer in Inflammatory Bowel Disease. Clin. Colon Rectal Surg. 2020, 33, 305–317. [Google Scholar] [CrossRef]
- Keller, D.S.; Windsor, A.; Cohen, R.; Chand, M. Colorectal cancer in inflammatory bowel disease: Review of the evidence. Tech. Coloproctol. 2019, 23, 3–13. [Google Scholar] [CrossRef]
- De Campos Silva, E.F.; Baima, J.P.; de Barros, J.R.; Tanni, S.E.; Schreck, T.; Saad-Hossne, R.; Sassaki, L.Y. Risk factors for ulcerative colitis-associated colorectal cancer: A retrospective cohort study. Medicine 2020, 99, e21686. [Google Scholar] [CrossRef] [PubMed]
- Leena, M.M.; Silvia, M.G.; Vinitha, K.; Moses, J.A.; Anandharamakrishnan, C. Synergistic potential of nutraceuticals: Mechanisms and prospects for futuristic medicine. Food Funct. 2020, 11, 9317–9337. [Google Scholar] [CrossRef] [PubMed]
- Sung, B.; Prasad, S.; Yadav, V.R.; Lavasanifar, A.; Aggarwal, B.B. Cancer and diet: How are they related? Free Radic. Res. 2011, 45, 864–879. [Google Scholar] [CrossRef] [Green Version]
- Sung, B.; Prasad, S.; Yadav, V.R.; Aggarwal, B.B. Cancer Cell Signaling Pathways Targeted by Spice-Derived Nutraceuticals. Nutr. Cancer 2012, 64, 173–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aggarwal, B.B.; Kunnumakkara, A.B.; Harikumar, K.B.; Tharakan, S.T.; Sung, B.; Anand, P. Potential of Spice-Derived Phytochemicals for Cancer Prevention. Planta Med. 2008, 74, 1560–1569. [Google Scholar] [CrossRef] [Green Version]
- Liao, N.C.; Shih, Y.L.; Ho, M.T.; Lu, T.J.; Lee, C.H.; Peng, S.F.; Leu, S.J.; Chung, J.G. Cardamonin induces immune responses and enhances survival rate in WEHI-3 cell–generated mouse leukemia in vivo. Environ. Toxicol. 2019, 35, 457–467. [Google Scholar] [CrossRef]
- Jin, J.; Qiu, S.; Wang, P.; Liang, X.; Huang, F.; Wu, H.; Zhang, B.; Zhang, W.; Tian, X.; Xu, R.; et al. Cardamonin inhibits breast cancer growth by repressing HIF-1α-dependent metabolic reprogramming. J. Exp. Clin. Cancer Res. 2019, 38, 377. [Google Scholar] [CrossRef] [Green Version]
- Badroon, N.A.; Abdul Majid, N.; Alshawsh, M.A. Antiproliferative and Apoptotic Effects of Cardamonin against Hepatocellular Carcinoma HepG2 Cells. Nutrients 2020, 12, 1757. [Google Scholar] [CrossRef]
- Nie, X.; Chen, H.; Niu, P.; Zhu, Y.; Zhou, J.; Jiang, L.; Li, D.; Lin, M.; Chen, Z.; Shi, D. DAP1 negatively regulates autophagy induced by cardamonin in SKOV3 cells. Cell Biol. Int. 2020, 44, 2192–2201. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Liu, L.; Liu, P.; Li, Y.; Lu, H.; Li, Y.; Zhang, G.; Duan, X. Cardamonin as a potential treatment for melanoma induces human melanoma cell apoptosis. Oncol. Lett. 2019, 19, 1393–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, W.; Li, C.; Qi, Q.; Shen, J.; Chang, K. Cardamonin induces G2/M arrest and apoptosis via activation of the JNK–FOXO3a pathway in breast cancer cells. Cell Biol. Int. 2020, 44, 177–188. [Google Scholar] [CrossRef]
- Zhang, J.; Sikka, S.; Siveen, K.S.; Lee, J.H.; Um, J.-Y.; Kumar, A.P.; Chinnathambi, A.; Alharbi, S.A.; Basappa; Rangappa, K.S.; et al. Cardamonin represses proliferation, invasion, and causes apoptosis through the modulation of signal transducer and activator of transcription 3 pathway in prostate cancer. Apoptosis 2017, 22, 158–168. [Google Scholar] [CrossRef]
- James, S.; Aparna, J.S.; Paul, A.M.; Lankadasari, M.B.; Mohammed, S.; Binu, V.S.; Santhoshkumar, T.R.; Reshmi, G.; Harikumar, K.B. Cardamonin inhibits colonic neoplasia through modulation of MicroRNA expression. Sci. Rep. 2017, 7, 13945. [Google Scholar] [CrossRef]
- Niu, P.; Li, J.; Chen, H.; Zhu, Y.; Zhou, J.; Shi, D. Anti-proliferative effect of cardamonin on mTOR inhibitor-resistant cancer cells. Mol. Med. Rep. 2019, 21, 1399–1407. [Google Scholar] [CrossRef] [PubMed]
- Hou, G.; Yuan, X.; Li, Y.; Hou, G.; Liu, X. Cardamonin, a natural chalcone, reduces 5-fluorouracil resistance of gastric cancer cells through targeting Wnt/β-catenin signal pathway. Investig. New Drugs 2020, 38, 329–339. [Google Scholar] [CrossRef]
- Qi, W.; Boliang, W.; Xiaoxi, T.; Guoqiang, F.; Jianbo, X.; Gang, W. Cardamonin protects against doxorubicin-induced cardiotoxicity in mice by restraining oxidative stress and inflammation associated with Nrf2 signaling. Biomed. Pharmacother. 2020, 122, 109547. [Google Scholar] [CrossRef] [PubMed]
- Kunnumakkara, A.B.; Shabnam, B.; Girisa, S.; Harsha, C.; Banik, K.; Devi, T.B.; Choudhury, R.; Sahu, H.; Parama, D.; Sailo, B.L.; et al. Inflammation, NF-κB, and Chronic Diseases: How are They Linked? Crit. Rev. Immunol. 2020, 40, 1–39. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, T.; Wu, J.; Otsuka, M.; Kishikawa, T.; Suzuki, N.; Takata, A.; Ohno, M.; Ishibashi, R.; Yamagami, M.; Nakagawa, R.; et al. Repression of MicroRNA Function Mediates Inflammation-associated Colon Tumorigenesis. Gastroenterology 2017, 152, 631–643. [Google Scholar] [CrossRef] [Green Version]
- Snider, A.J.; Bialkowska, A.B.; Ghaleb, A.M.; Yang, V.W.; Obeid, L.M.; Hannun, Y.A. Murine Model for Colitis-Associated Cancer of the Colon. Methods Mol. Biol. 2016, 1438, 245–254. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Nagahashi, M.; Kim, E.Y.; Harikumar, K.B.; Yamada, A.; Huang, W.-C.; Hait, N.C.; Allegood, J.C.; Price, M.M.; Avni, D.; et al. Sphingosine-1-Phosphate Links Persistent STAT3 Activation, Chronic Intestinal Inflammation, and Development of Colitis-Associated Cancer. Cancer Cell 2013, 23, 107–120. [Google Scholar] [CrossRef] [Green Version]
- Bender, D.; Schwarz, G. Nitrite-dependent nitric oxide synthesis by molybdenum enzymes. FEBS Lett. 2018, 592, 2126–2139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giustarini, D.; Rossi, R.; Milzani, A.; Dalle-Donne, I. Nitrite and Nitrate Measurement by Griess Reagent in Human Plasma: Evaluation of Interferences and Standardization. Methods Enzymol. 2008, 440, 361–380. [Google Scholar] [CrossRef] [PubMed]
- Karin, M.; Cao, Y.; Greten, F.R.; Li, Z.-W. NF-κB in cancer: From innocent bystander to major culprit. Nat. Rev. Cancer 2002, 2, 301–310. [Google Scholar] [CrossRef]
- Baltimore, D. NF-κB is 25. Nat. Immunol. 2011, 12, 683–685. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Rimm, D.L.; Lin, Y.; Shih, W.J.; Reiss, M. Loss of Smad Signaling in Human Colorectal Cancer Is Associated with Advanced Disease and Poor Prognosis. Cancer J. 2003, 9, 302–312. [Google Scholar] [CrossRef]
- Xu, D.; Wang, W.; Bian, T.; Yang, W.; Shao, M.; Yang, H. Increased expression of O-GlcNAc transferase (OGT) is a biomarker for poor prognosis and allows tumorigenesis and invasion in colon cancer. Int. J. Clin. Exp. Pathol. 2019, 12, 1305–1314. [Google Scholar] [PubMed]
- Zhang, Y.; Liu, Y.; Ye, Y.; Shen, D.; Zhang, H.; Huang, H.; Li, S.; Wang, S.; Ren, J. Quantitative proteome analysis of colorectal cancer-related differential proteins. J. Cancer Res. Clin. Oncol. 2017, 143, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hu, H.; Wang, Y.; Fan, Y.; Yang, Y.; Guo, B.; Xie, X.; Lian, J.; Jiang, B.; Han, B.; et al. CUL4B contributes to cancer stemness by repressing tumor suppressor miR34a in colorectal cancer. Oncogenesis 2020, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Maceyka, M.; Spiegel, S. Sphingolipid metabolites in inflammatory disease. Nat. Cell Biol. 2014, 510, 58–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, S.; Yuan, Q.; Yu, N.; Liu, B.; Huang, G.; Yuan, X. Cardamonin attenuates chronic inflammation and tumorigenesis in colon. Cell Cycle 2019, 18, 3275–3287. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Sun, A.; Deng, C.; Zhang, J.; Wu, X.; Wei, X.; Mani, S.; Dou, W.; Wang, Z. The anti-inflammatory effect and potential mechanism of cardamonin in DSS-induced colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G517–G527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.A.; Abd Al Haleem, E.N.; Khaleel, S.A.-H.; Sallam, A.S. Protective effect of cardamonin against acetic acid-induced ulcerative colitis in rats. Pharmacol. Rep. 2017, 69, 268–275. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
James, S.; Aparna, J.S.; Babu, A.; Paul, A.M.; Lankadasari, M.B.; Athira, S.R.; Kumar, S.S.; Vijayan, Y.; Namitha, N.N.; Mohammed, S.; et al. Cardamonin Attenuates Experimental Colitis and Associated Colorectal Cancer. Biomolecules 2021, 11, 661. https://doi.org/10.3390/biom11050661
James S, Aparna JS, Babu A, Paul AM, Lankadasari MB, Athira SR, Kumar SS, Vijayan Y, Namitha NN, Mohammed S, et al. Cardamonin Attenuates Experimental Colitis and Associated Colorectal Cancer. Biomolecules. 2021; 11(5):661. https://doi.org/10.3390/biom11050661
Chicago/Turabian StyleJames, Shirley, Jayasekharan S. Aparna, Anu Babu, Aswathy Mary Paul, Manendra Babu Lankadasari, Subha R. Athira, Sreesha S. Kumar, Yadu Vijayan, Narayanan N. Namitha, Sabira Mohammed, and et al. 2021. "Cardamonin Attenuates Experimental Colitis and Associated Colorectal Cancer" Biomolecules 11, no. 5: 661. https://doi.org/10.3390/biom11050661