
High-Throughput LC-ESI-MS/MS Metabolomics Approach Reveals Regulation of Metabolites Related to Diverse Functions in Mature Fruit of Grafted Watermelon

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Sample Preparation and Extraction Chemicals
2.3. Analytical Condition of LC-MS/MS
2.4. Statistical Analysis
3. Results
3.1. Metabolome Comparison between Pumpkin-Grafted (PGW) and Self-Rooted Watermelon (SRW)
3.2. Major Metabolites
3.3. Carbon Metabolism
3.4. Carbohydrate Metabolism
3.5. Arginine Metabolism
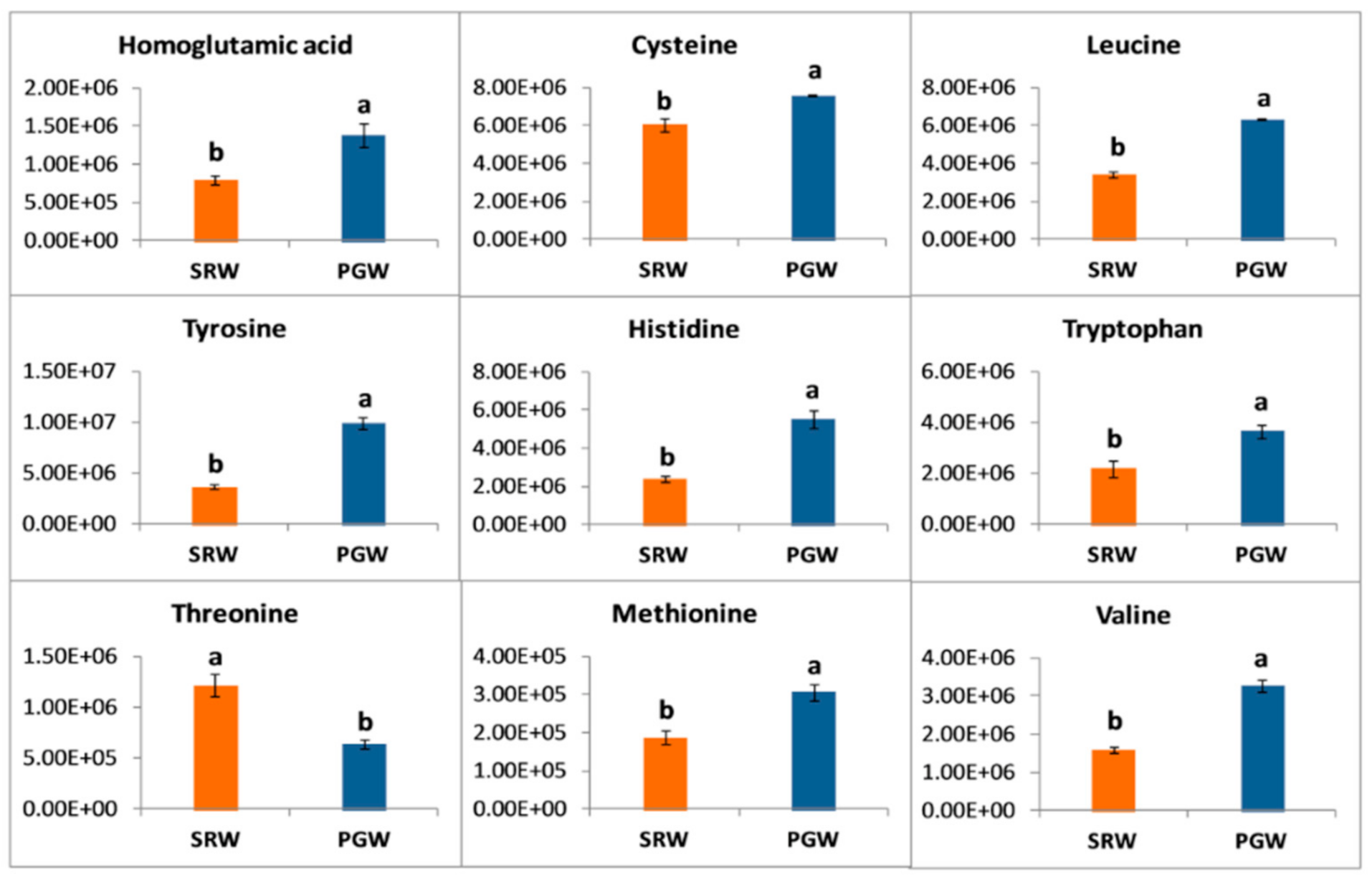
3.6. Amino Acid Metabolism
3.7. Glutathione Metabolism
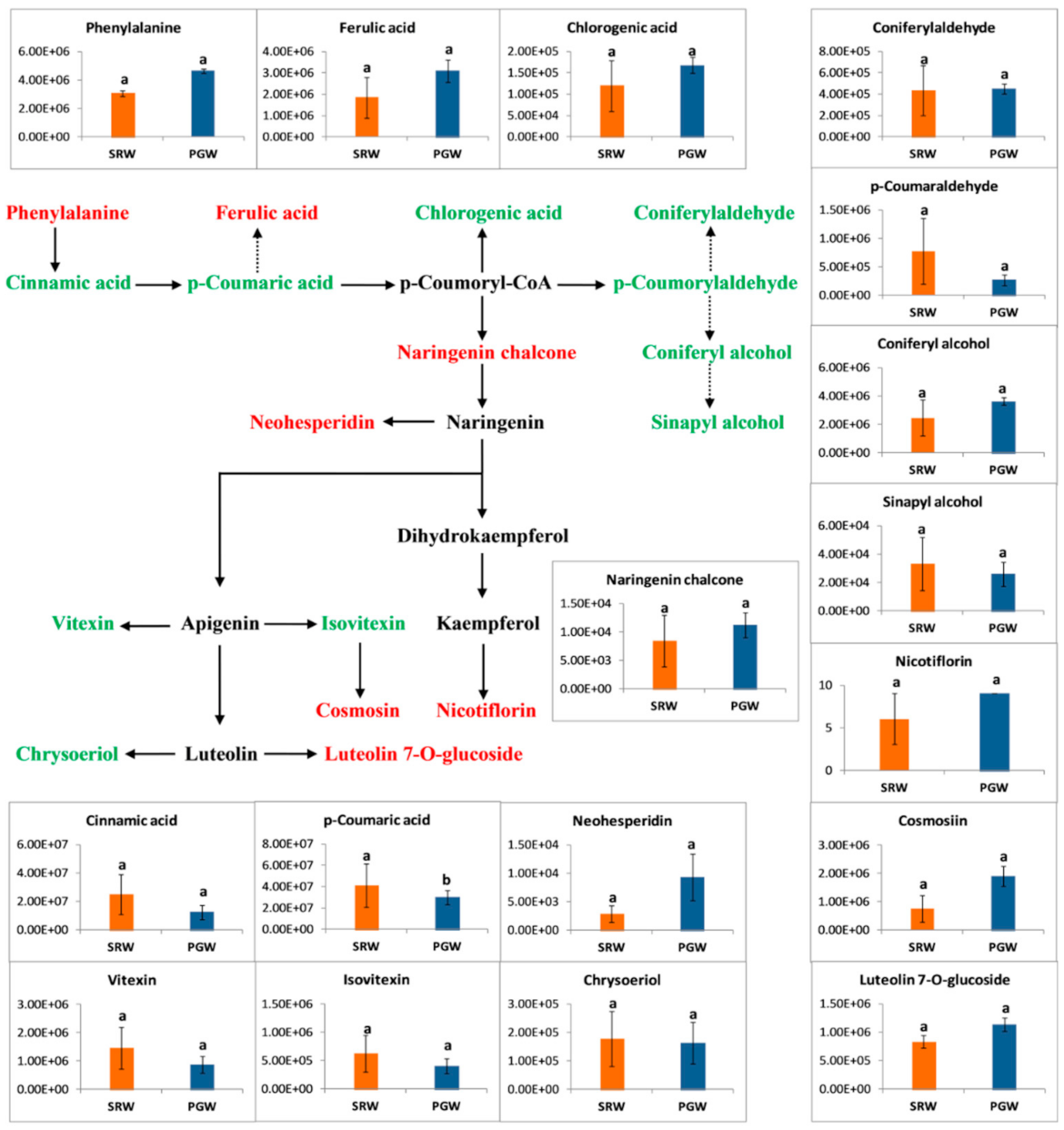
3.8. Phenylpropanoid Metabolism
3.9. Linolenic Acid Metabolism and Fatty Acids
3.10. Secondary Metabolites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collins, J.K.; Wu, G.; Perkins-Veazie, P.; Spears, K.; Claypool, P.L.; Baker, R.A.; Clevidence, B.A. Watermelon consumption increases plasma arginine concentrations in adults. Nutrition 2007, 23, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Juliet, P.A.R.; Matsui-Hirai, H.; Miyazaki, A.; Fukatsu, A.; Funami, J.; Iguchi, A.; Ignarro, L.J. L-citrulline and L-arginine supplementation retards the progression of high-cholesterol-diet-induced atherosclerosis in rabbits. Proc. Natl. Acad. Sci. USA 2005, 102, 13681–13686. [Google Scholar] [CrossRef] [Green Version]
- Perkins-Veazie, P.; Collins, J.K.; Davis, A.R.; Roberts, W. Carotenoid Content of 50 Watermelon Cultivars. J. Agric. Food Chem. 2006, 54, 2593–2597. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085. [Google Scholar] [CrossRef]
- Saito, K.; Matsuda, F. Metabolomics for Functional Genomics, Systems Biology, and Biotechnology. Annu. Rev. Plant Biol. 2010, 61, 463–489. [Google Scholar] [CrossRef]
- Cassi-Lit, M.T.; Tanner, G.J.; Nayudu, M.; Whitecross, M. Isovitexin-2’-O-β-[6-O-E-p-coumaroylglucopyranoside] from UV-B Irradiated Leaves of Rice, Oryza sativa L. Inhibits Fertility of Helicoverpa armigera. Photochem. Photobiol. 2007, 83, 1167–1173. [Google Scholar] [CrossRef] [PubMed]
- D’Auria, J.C.; Gershenzon, J. The secondary metabolism of Arabidopsis thaliana: Growing like a weed. Curr. Opin. Plant Biol. 2005, 8, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Schauer, N.; Semel, Y.; Balbo, I.; Steinfath, M.; Repsilber, D.; Selbig, J.; Pleban, T.; Zamir, D.; Fernie, A.R. Mode of Inheritance of Primary Metabolic Traits in Tomato. Plant Cell 2008, 20, 509–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, S.; Liu, J.; Zheng, Y.; Huang, M.; Zhang, H.; Gong, G.; He, H.; Ren, Y.; Zhong, S.; Fei, Z.; et al. Characterization of transcriptome dynamics during watermelon fruit development: Sequencing, assembly, annotation and gene expression profiles. BMC Genom. 2011, 12, 454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yativ, M.; Harary, I.; Wolf, S. Sucrose accumulation in watermelon fruits: Genetic variation and biochemical analysis. J. Plant Physiol. 2010, 167, 589–596. [Google Scholar] [CrossRef]
- Gao, L.; Zhao, S.; Lu, X.; He, N.; Zhu, H.; Dou, J.; Liu, W. Comparative transcriptome analysis reveals key genes potentially related to soluble sugar and organic acid accumulation in watermelon. PLoS ONE 2018, 13, e0190096. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Li, Y.; Chen, R.; Bao, J.; Yang, G. The relationship between volatile esters and free amino acids in different parts of ripe melon (Cucumis melo L.). Plant Physiol. 2008, 44, 215–220. [Google Scholar]
- Zhang, Q.; Ruan, J. Tea: Analysis and Tasting. In Encyclopedia of Food and Health; Elsevier: Amsterdam, The Netherlands, 2016; pp. 256–267. [Google Scholar]
- Yuan, P.; He, N.; Umer, M.; Zhao, S.; Diao, W.; Zhu, H.; Dou, J.; Kaseb, M.; Kuang, H.; Lu, X.; et al. Comparative Metabolomic Profiling of Citrullus spp. Fruits Provides Evidence for Metabolomic Divergence during Domestication. Metabolites 2021, 11, 78. [Google Scholar] [CrossRef] [PubMed]
- Arah, I.K.; Amaglo, H.; Kumah, E.K.; Ofori, H. Preharvest and Postharvest Factors Affecting the Quality and Shelf Life of Harvested Tomatoes: A Mini Review. Int. J. Agron. 2015, 2015, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Kader, A.A. Flavor quality of fruits and vegetables. J. Sci. Food Agric. 2008, 88, 1863–1868. [Google Scholar] [CrossRef]
- Léchaudel, M.; Joas, J. An overview of preharvest factors influencing mango fruit growth, quality and postharvest behaviour. Braz. J. Plant Physiol. 2007, 19, 287–298. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.K.; Kader, A.A. Preharvest and postharvest factors influencing vitamin C content of horticultural crops. Postharvest Biol. Technol. 2000, 20, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Guo, S.; He, H.; Zhang, H.; Gong, G.; Ren, Y.; Xu, Y. Dynamic characteristics of sugar accumulation and related enzyme activities in sweet and non-sweet watermelon fruits. Acta Physiol. Plant. 2013, 35, 3213–3222. [Google Scholar] [CrossRef]
- Lee, J.-M. Cultivation of Grafted Vegetables I. Current Status, Grafting Methods, and Benefits. HortScience 1994, 29, 235–239. [Google Scholar] [CrossRef]
- Rouphael, Y.; Venema, J.H.; Edelstein, M.; Savvas, D.; Colla, G.; Ntatsi, G.; Ben-Hur, M.; Pradeep, K.; Schwarz, D. Grafting as a Tool for Tolerance of Abiotic Stress; CABI Publishing: Wallingford, UK, 2017; pp. 171–215. [Google Scholar]
- Condurso, C.; Verzera, A.; Dima, G.; Tripodi, G.; Crinò, P.; Paratore, A.; Romano, D. Effects of different rootstocks on aroma volatile compounds and carotenoid content of melon fruits. Sci. Hortic. 2012, 148, 9–16. [Google Scholar] [CrossRef]
- Davis, A.R.; Perkins-Veazie, P. Rootstock effects on plant vigor and watermelon fruit quality. Rep. Cucurbit. Genet. Coop. 2005, 28, 39. [Google Scholar]
- Davis, A.R.; Perkins-Veazie, P.; Hassell, R.; Levi, A.; King, S.R.; Zhang, X. Grafting Effects on Vegetable Quality. HortScience 2008, 43, 1670–1672. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Rouphael, Y.; Leonardi, C.; Bie, Z. Role of grafting in vegetable crops grown under saline conditions. Sci. Hortic. 2010, 127, 147–155. [Google Scholar] [CrossRef]
- Rouphael, Y.; Cardarelli, M.; Colla, G.; Rea, E. Yield, Mineral Composition, Water Relations, and Water Use Efficiency of Grafted Mini-Watermelon Plants under Deficit Irrigation. HortScience 2008, 43, 730–736. [Google Scholar] [CrossRef] [Green Version]
- Savvas, D.; Papastavrou, D.; Ntatsi, G.; Ropokis, A.; Olympios, C.; Hartmann, H.; Schwarz, D. Interactive Effects of Grafting and Manganese Supply on Growth, Yield, and Nutrient Uptake by Tomato. HortScience 2009, 44, 1978–1982. [Google Scholar] [CrossRef] [Green Version]
- Venema, J.H.; Dijk, B.E.; Bax, J.M.; Van Hasselt, P.R.; Elzenga, J.T.M. Grafting tomato (Solanum lycopersicum) onto the rootstock of a high-altitude accession of Solanum habrochaites improves suboptimal-temperature tolerance. Environ. Exp. Bot. 2008, 63, 359–367. [Google Scholar] [CrossRef]
- Aloni, B.; Cohen, R.; Karni, L.; Aktas, H.; Edelstein, M. Hormonal signaling in rootstock–scion interactions. Sci. Hortic. 2010, 127, 119–126. [Google Scholar] [CrossRef]
- Saini, M.K.; Capalash, N.; Kaur, C.; Singh, S.P. Comprehensive metabolic profiling to decipher the influence of rootstocks on fruit juice metabolome of Kinnow (C. nobilis × C. deliciosa). Sci. Hortic. 2019, 257, 108673. [Google Scholar] [CrossRef]
- Spinardi, A.; Visai, C.; Bertazza, G. Effect of rootstock on fruit quality of two sweet cherry cultivars. Acta Hortic. 2005, 201–206. [Google Scholar] [CrossRef]
- Liu, Y.-F.; Qi, H.-Y.; Bai, C.-M.; Qi, M.-F.; Xu, C.-Q.; Hao, J.-H.; Li, Y.; Li, T.-L. Grafting Helps Improve Photosynthesis and Carbohydrate Metabolism in Leaves of Muskmelon. Int. J. Biol. Sci. 2011, 7, 1161–1170. [Google Scholar] [CrossRef] [Green Version]
- Miao, L.; Di, Q.; Sun, T.; Li, Y.; Duan, Y.; Wang, J.; Yan, Y.; He, C.; Wang, C.; Yu, X. Integrated Metabolome and Transcriptome Analysis Provide Insights into the Effects of Grafting on Fruit Flavor of Cucumber with Different Rootstocks. Int. J. Mol. Sci. 2019, 20, 3592. [Google Scholar] [CrossRef] [Green Version]
- Deng, W.-W.; Han, J.; Fan, Y.; Tai, Y.; Zhu, B.; Lu, M.; Wang, R.; Wan, X.; Zhang, Z.-Z. Uncovering tea-specific secondary metabolism using transcriptomic and metabolomic analyses in grafts of Camellia sinensis and C. oleifera. Tree Genet. Genomes 2018, 14, 23. [Google Scholar] [CrossRef]
- Bletsos, F.; Passam, H. Grafting: An environmentally friendly technique to overcome soil-borne diseases and improve the out of season production of watermelon, cucumber and melon. In Horticulture in the 21st Century; Sampson, A.N., Ed.; Nova Science: New York, NY, USA, 2010; pp. 81–120. [Google Scholar]
- Petropoulos, S.; Olympios, C.; Ropokis, A.; Vlachou, G.; Ntatsi, G.; Paraskevopoulos, A.; Passam, H. Fruit volatiles, quality, and yield of watermelon as affected by grafting. J. Agric. Sci. Technol. 2014, 16, 873–885. [Google Scholar]
- Proietti, S.; Rouphael, Y.; Colla, G.; Cardarelli, M.; De Agazio, M.; Zacchini, M.; Rea, E.; Moscatello, S.; Battistelli, A. Fruit quality of mini-watermelon as affected by grafting and irrigation regimes. J. Sci. Food Agric. 2008, 88, 1107–1114. [Google Scholar] [CrossRef]
- Tietel, Z.; Srivastava, S.; Fait, A.; Tel-Zur, N.; Carmi, N.; Raveh, E. Impact of scion/rootstock reciprocal effects on metabolomics of fruit juice and phloem sap in grafted Citrus reticulata. PLoS ONE 2020, 15, e0227192. [Google Scholar] [CrossRef]
- Grieneisen, M.L.; Aegerter, B.J.; Stoddard, C.S.; Zhang, M. Yield and fruit quality of grafted tomatoes, and their potential for soil fumigant use reduction. A meta-analysis. Agron. Sustain. Dev. 2018, 38, 29. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, T.; Qi, H.; Qi, M.; Chen, W.; Xu, C.; Li, J. Effects on carbohydrate and amino acids accumulation in muskmelon fruiT (Cucumis melo L.) with different grafting rootstocks. Acta Hortic. 2010, 871, 579–588. [Google Scholar] [CrossRef]
- Lee, J.; Steenwerth, K.L. Rootstock and vineyard floor management influence on ‘Cabernet Sauvignon’ grape yeast assimilable nitrogen (YAN). Food Chem. 2011, 127, 926–933. [Google Scholar] [CrossRef] [PubMed]
- Aslam, A.; Shengjie, Z.; Xuqiang, L.; Nan, H.; Wenge, L. Rootstock mediates transcriptional regulation of citrulline metabolism in grafted watermelon. Braz. J. Biol. 2021, 81, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Fredes, A.; Roselló, S.; Beltrán, J.; Cebolla-Cornejo, J.; Pérez-de-Castro, A.; Gisbert, C.; Picó, M.B. Fruit quality assessment of watermelons grafted onto citron melon rootstock. J. Sci. Food Agric. 2017, 97, 1646–1655. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Cardarelli, M.; Bassal, A.; Leonardi, C.; Giuffrida, F.; Colla, G. Vegetable quality as affected by genetic, agronomic and environmental factors. J. Food Agric. Environ. 2012, 10, 680–688. [Google Scholar]
- Rouphael, Y.; Schwarz, D.; Krumbein, A.; Colla, G. Impact of grafting on product quality of fruit vegetables. Sci. Hortic. 2010, 127, 172–179. [Google Scholar] [CrossRef]
- Fallik, E.; Alkalai-Tuvia, S.; Chalupowicz, D.; Zutahy, Y.; Zaaroor, M.; Beniches, M.; Gamliel, A. Effects of rootstock and soil disinfection on quality of grafted watermelon fruit (Citrullus lanatus L.): A two-year study. Isr. J. Plant Sci. 2016, 63, 38–44. [Google Scholar] [CrossRef]
- King, S.R.; Davis, A.R.; Liu, W.; Levi, A. Grafting for Disease Resistance. HortScience 2008, 43, 1673–1676. [Google Scholar] [CrossRef] [Green Version]
- Kyriacou, M.C.; Soteriou, G.A.; Rouphael, Y.; Siomos, A.S.; Gerasopoulos, D.; Soteriou, G. Configuration of watermelon fruit quality in response to rootstock-mediated harvest maturity and postharvest storage. J. Sci. Food Agric. 2015, 96, 2400–2409. [Google Scholar] [CrossRef]
- Villocino, J.S.B.; Quevedo, M. Effects of biochar on physicochemical and sensory quality of watermelon (Citrullus lanatus Thunb.) fruit from grafted and non-grafted plants. In Proceedings of the II Southeast Asia Symposium on Quality Management in Postharvest Systems 1088, Vientiane, Laos, 4–6 December 2013; pp. 481–484. [Google Scholar]
- Li, S.; Dong, X.; Fan, G.; Yang, Q.; Shi, J.; Wei, W.; Zhao, F.; Li, N.; Wang, X.; Wang, F.; et al. Comprehensive Profiling and Inheritance Patterns of Metabolites in Foxtail Millet. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Li, R.; Ren, L.; Gao, X.; Zhang, Y.; Ma, Z.; Ma, D.; Luo, Y. A comparative metabolomics study of flavonoids in sweet potato with different flesh colors (Ipomoea batatas (L.) Lam). Food Chem. 2018, 260, 124–134. [Google Scholar] [CrossRef]
- Sweetlove, L.J.; Beard, K.F.; Nunes-Nesi, A.; Fernie, A.R.; Ratcliffe, R.G. Not just a circle: Flux modes in the plant TCA cycle. Trends Plant Sci. 2010, 15, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, R.; Barchi, B.; Turlapati, S.A.; Gagne, M.; Minocha, R.; Long, S.; Minocha, S.C. Glutamate, Ornithine, Arginine, Proline, and Polyamine Metabolic Interactions: The Pathway Is Regulated at the Post-Transcriptional Level. Front. Plant Sci. 2016, 7, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slocum, R.D. Genes, enzymes and regulation of arginine biosynthesis in plants. Plant Physiol. Biochem. 2005, 43, 729–745. [Google Scholar] [CrossRef]
- Shargool, D.; Jain, J.; McKay, G. Ornithine biosynthesis, and arginine biosynthesis and degradation in plant cells. Phytochemistry 1988, 27, 1571–1574. [Google Scholar] [CrossRef]
- Micallef, B.J.; Shelp, B.J. Arginine Metabolism in Developing Soybean Cotyledons. Plant Physiol. 1989, 90, 624–630. [Google Scholar] [CrossRef] [Green Version]
- Winter, G.; Todd, C.D.; Trovato, M.; Forlani, G.; Funck, D. Physiological implications of arginine metabolism in plants. Front. Plant Sci. 2015, 6, 534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hell, R.; Bergmann, L. λ-Glutamylcysteine synthetase in higher plants: Catalytic properties and subcellular localization. Planta 1990, 180, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Xiang, C.; Oliver, D.J. Glutathione and its central role in mitigating plant stress. In Handbook of Plant and Crop Stress; Marcel Dekker Inc.: New York, NY, USA, 1999; pp. 697–707. [Google Scholar]
- Fraser, C.M.; Chapple, C. The Phenylpropanoid Pathway in Arabidopsis. Arab. Book 2011, 9, e0152. [Google Scholar] [CrossRef] [Green Version]
- Aragüez, I.; Fernández, V.V. Metabolic engineering of aroma components in fruits. Biotechnol. J. 2013, 8, 1144–1158. [Google Scholar] [CrossRef]
- Matsui, K.; Ujita, C.; Fujimoto, S.-H.; Wilkinson, J.; Hiatt, B.; Knauf, V.; Kajiwara, T.; Feussner, I. Fatty acid 9- and 13-hydroperoxide lyases from cucumber. FEBS Lett. 2000, 481, 183–188. [Google Scholar] [CrossRef] [Green Version]
- Papagianni, M.; Avramidis, N.; Filiousis, G. Glycolysis and the regulation of glucose transport in Lactococcus lactis spp. lactis in batch and fed-batch culture. Microb. Cell Factories 2007, 6, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasternack, C. Perception, signaling and cross-talk of jasmonates and the seminal contributions of the Daoxin Xie’s lab and the Chuanyou Li’s lab. Plant Cell Rep. 2014, 33, 707–718. [Google Scholar] [CrossRef]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef]
- Chen, W.; Gao, Y.; Xie, W.; Gong, L.; Lu, K.; Wang, W.; Li, Y.; Liu, X.; Zhang, H.; Dong, H.; et al. Genome-wide association analyses provide genetic and biochemical insights into natural variation in rice metabolism. Nat. Genet. 2014, 46, 714–721. [Google Scholar] [CrossRef] [PubMed]
- Rolland, F.; Baena-Gonzalez, E.; Sheen, J. Sugar sensing and signaling in plants: Conserved and Novel Mechanisms. Annu. Rev. Plant Biol. 2006, 57, 675–709. [Google Scholar] [CrossRef] [Green Version]
- Fagard, M.; Launay, A.; Clément, G.; Courtial, J.; Dellagi, A.; Farjad, M.; Krapp, A.; Soulié, M.-C.; Masclaux-Daubresse, C. Nitrogen metabolism meets phytopathology. J. Exp. Bot. 2014, 65, 5643–5656. [Google Scholar] [CrossRef] [PubMed]
- Galili, G.; Avin-Wittenberg, T.; Angelovici, R.; Fernie, A.R. The role of photosynthesis and amino acid metabolism in the energy status during seed development. Front. Plant Sci. 2014, 5, 447. [Google Scholar] [CrossRef] [PubMed]
- Häusler, R.E.; Ludewig, F.; Krueger, S. Amino acids—A life between metabolism and signaling. Plant Sci. 2014, 229, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Moe, L.A. Amino acids in the rhizosphere: From plants to microbes. Am. J. Bot. 2013, 100, 1692–1705. [Google Scholar] [CrossRef] [PubMed]
- Pratelli, R.; Pilot, G. Regulation of amino acid metabolic enzymes and transporters in plants. J. Exp. Bot. 2014, 65, 5535–5556. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Balazadeh, S.; Tohge, T.; Erban, A.; Giavalisco, P.; Kopka, J.; Mueller-Roeber, B.; Fernie, A.R.; Hoefgen, R. Comprehensive Dissection of Spatiotemporal Metabolic Shifts in Primary, Secondary, and Lipid Metabolism during Developmental Senescence in Arabidopsis. Plant Physiol. 2013, 162, 1290–1310. [Google Scholar] [CrossRef] [Green Version]
- Zeier, J. New insights into the regulation of plant immunity by amino acid metabolic pathways. Plant Cell Environ. 2013, 36, 2085–2103. [Google Scholar] [CrossRef]
- Halkier, B.A.; Gershenzon, J. Biology and biochemistry of glucosinolates. Annu. Rev. Plant Biol. 2006, 57, 303–333. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Zhang, J.; Sun, H.; Salse, J.; Lucas, W.J.; Zhang, H.; Zheng, Y.; Mao, L.; Ren, Y.; Wang, Z.; et al. The draft genome of watermelon (Citrullus lanatus) and resequencing of 20 diverse accessions. Nat. Genet. 2013, 45, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Masclaux-Daubresse, C.; Daniel-Vedele, F.; Dechorgnat, J.; Chardon, F.; Gaufichon, L.; Suzuki, A. Nitrogen uptake, assimilation and remobilization in plants: Challenges for sustainable and productive agriculture. Ann. Bot. 2010, 105, 1141–1157. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, M.A.; Wang, L.; Jiao, Y.; Chen, C.; Zhao, L.; Mei, M.; Yu, Y.; Bie, Z.; Huang, Y. Pumpkin rootstock improves nitrogen use efficiency of watermelon scion by enhancing nutrient uptake, cytokinin content, and expression of nitrate reductase genes. Plant Growth Regul. 2017, 82, 233–246. [Google Scholar] [CrossRef]
- Scheible, W.-R.; Morcuende, R.; Czechowski, T.; Fritz, C.; Osuna, D.; Palacios-Rojas, N.; Schindelasch, D.; Thimm, O.; Udvardi, M.K.; Stitt, M. Genome-Wide Reprogramming of Primary and Secondary Metabolism, Protein Synthesis, Cellular Growth Processes, and the Regulatory Infrastructure of Arabidopsis in Response to Nitrogen. Plant Physiol. 2004, 136, 2483–2499. [Google Scholar] [CrossRef] [Green Version]
- Suh, M.C.; Hahne, G.; Liu, J.R.; Stewart, C.N. Plant lipid biology and biotechnology. Plant Cell Rep. 2015, 34, 517–518. [Google Scholar] [CrossRef] [Green Version]
- Bulder, H.; Speek, E.; Van Hasselt, P.; Kuiper, P. Growth Temperature and Lipid Composition of Cucumber Genotypes Differing in Adaptation to Low Energy Conditions. J. Plant Physiol. 1991, 138, 655–660. [Google Scholar] [CrossRef]
- Mohammed, S.; Humidan, M.; Boras, M.; Abdalla, O. The Role of Grafting Tomato and Watermelon on Different Rootstocks on Their Chemical Contents. Int. J. Agric. Res. 2009, 4, 362–369. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Oxidant and antioxidant signalling in plants: A re-evaluation of the concept of oxidative stress in a physiological context. Plant Cell Environ. 2005, 28, 1056–1071. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox Homeostasis and Antioxidant Signaling: A Metabolic Interface between Stress Perception and Physiological Responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Fujita, M. Exogenous sodium nitroprusside alleviates arsenic-induced oxidative stress in wheat (Triticum aestivum L.) seedlings by enhancing antioxidant defense and glyoxalase system. Ecotoxicology 2013, 22, 584–596. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hossain, M.A.; Da Silva, J.A.T.; Fujita, M. Plant Response and Tolerance to Abiotic Oxidative Stress: Antioxidant Defense Is a Key Factor. In Crop Stress and Its Management: Perspectives and Strategies; Springer: New York, NY, USA; pp. 261–315.
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Nitric oxide modulates antioxidant defense and the methylglyoxal detoxification system and reduces salinity-induced damage of wheat seedlings. Plant Biotechnol. Rep. 2011, 5, 353–365. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Fujita, M. Exogenous nitric oxide alleviates high temperature induced oxidative stress in wheat (Triticum aestivum L.) seedlings by modulating the antioxidant defense and glyoxalase system. Aust. J. Crop. Sci. 2012, 6, 1314. [Google Scholar]
- Beier, R.C.; Nigg, H.N. Natural Toxicants in Foods. In Phytochemical Resources for Medicine and Agriculture; Springer: New York, NY, USA, 1992; pp. 247–367. [Google Scholar]
- Hahlbrock, K.; Scheel, D. Physiology and molecular biology of phenylpropanoid metabolism. Annu. Rev. Plant Biol. 1989, 40, 347–369. [Google Scholar] [CrossRef]
- Jaakola, L.; Määttä, K.; Pirttilä, A.M.; Törrönen, R.; Kärenlampi, S.; Hohtola, A. Expression of genes involved in anthocyanin biosynthesis in relation to anthocyanin, proanthocyanidin, and flavonol levels during bilberry fruit development. Plant. Physiol. 2002, 130, 729–739. [Google Scholar] [CrossRef] [Green Version]
- Kühnau, J. The Flavonoids. A Class of Semi-Essential Food Components: Their Role in Human Nutrition. World Rev. Nutr. Diet. 2015, 24, 117–191. [Google Scholar] [CrossRef]
- Paolacci, A.R.; D’Ovidio, R.; Marabottini, R.; Nali, C.; Lorenzini, G.; Abenavoli, M.R.; Badiani, M. Research Note: Ozone induces a differential accumulation of phenyalanine ammonia-lyase, chalcone synthase and chalcone isomerase RNA transcripts in sensitive and resistant bean cultivars. Funct. Plant Biol. 2001, 28, 425. [Google Scholar] [CrossRef]
- Seigler, D.S. Flavonoids. In Plant Secondary Metabolism; Metzler, J.B., Ed.; Springer: New York, NY, USA, 1998; pp. 151–192. [Google Scholar]
- Sommer, H.; Saedler, H. Structure of the chalcone synthase gene of Antirrhinum majus. Mol. Genet. Genom. 1986, 202, 429–434. [Google Scholar] [CrossRef]
- Treutter, D.; Schmid, P.; Feucht, W. Wall-bound phenols and peroxidase activity in shoots of Prunus. I. Isolation and identification of phenolic acids. Gartenbauwissenschaft 1990, 55, 69–72. [Google Scholar]
- Wienand, U.; Weydemann, U.; Niesbach-Klösgen, U.; Peterson, P.A.; Saedler, H. Molecular cloning of the c2 locus of Zea mays, the gene coding for chalcone synthase. Mol. Genet. Genom. 1986, 203, 202–207. [Google Scholar] [CrossRef]
- Evrenosoğlu, Y. Leaf phenolic content of some squash rootstocks used on watermelon (Citrullus lanatus (thunb.) Matsum and Nakai) growing and phenolic accumulation on grafted cultivar. Afr. J. Agric. Res. 2010, 5, 732–737. [Google Scholar]
- Asensi-Fabado, M.A.; Munné-Bosch, S. Vitamins in plants: Occurrence, biosynthesis and antioxidant function. Trends Plant Sci. 2010, 15, 582–592. [Google Scholar] [CrossRef]
- Ruiz, J.M.; Blasco, B.; Rivero, R.M.; Romero, L. Nicotine-free and salt-tolerant tobacco plants obtained by grafting to salinity-resistant rootstocks of tomato. Physiol. Plant. 2005, 124, 465–475. [Google Scholar] [CrossRef]
- Amirjani, M.R. Effects of drought stress on the alkaloid contents and growth parameters of Catharanthus roseus. J. Agric. Biol. Sci. 2013, 8, 745–750. [Google Scholar]
- Liu, W.; Zhao, S.; Cheng, Z.; Wan, X.; Yan, Z.; King, S. Lycopene and Citrulline Contents in Watermelon (Citrullus lanatus) Fruit with Different Ploidy and Changes during Fruit Development. Acta Hortic. 2010, 543–550. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aslam, A.; Zhao, S.; Lu, X.; He, N.; Zhu, H.; Malik, A.U.; Azam, M.; Liu, W. High-Throughput LC-ESI-MS/MS Metabolomics Approach Reveals Regulation of Metabolites Related to Diverse Functions in Mature Fruit of Grafted Watermelon. Biomolecules 2021, 11, 628. https://doi.org/10.3390/biom11050628
Aslam A, Zhao S, Lu X, He N, Zhu H, Malik AU, Azam M, Liu W. High-Throughput LC-ESI-MS/MS Metabolomics Approach Reveals Regulation of Metabolites Related to Diverse Functions in Mature Fruit of Grafted Watermelon. Biomolecules. 2021; 11(5):628. https://doi.org/10.3390/biom11050628
Chicago/Turabian StyleAslam, Ali, Shengjie Zhao, Xuqiang Lu, Nan He, Hongju Zhu, Aman Ullah Malik, Muhammad Azam, and Wenge Liu. 2021. "High-Throughput LC-ESI-MS/MS Metabolomics Approach Reveals Regulation of Metabolites Related to Diverse Functions in Mature Fruit of Grafted Watermelon" Biomolecules 11, no. 5: 628. https://doi.org/10.3390/biom11050628