
Central Alteration in Peripheral Neuropathy of Trembler-J Mice: Hippocampal pmp22 Expression and Behavioral Profile in Anxiety Tests

 , , ,
, , ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Mice Phenotyping
2.3. Behavioral Experiments
2.3.1. Open Field Test
2.3.2. Elevated Plus Maze Test
2.4. Brain Processing to Cryostat and Vibratome Sectioning
2.5. Probe Synthesis for In Situ Hybridization
2.6. In Situ Hybridization
2.7. Immunostaining Post-ISH
2.8. Immunohistochemistry of Soluble and Aggregated PMP22 to Fluorescent Signal Quantification
2.9. Hippocampal Dissection
2.10. RNA Purification and RT—qPCR in Hippocampal Tissue
- pmp22 forward: 5′-GAATTCCTGTTCCTGTTCTTCTGCCAGCTC-3′
- pmp22 reverse: 5′-AAGCTTGTAGATGGCCGCTGCACTCATC-3′
- β-actin forward: 5′-TATGTTGCCCTAGACTTCGAGC-3′
- β-actin reverse: 5′-CAGCTCATAGCTCTTCTCCAGG-3′
2.11. Confocal Microscopy
2.12. Fluorescent Image Analysis
2.13. Statistical Analysis
3. Results
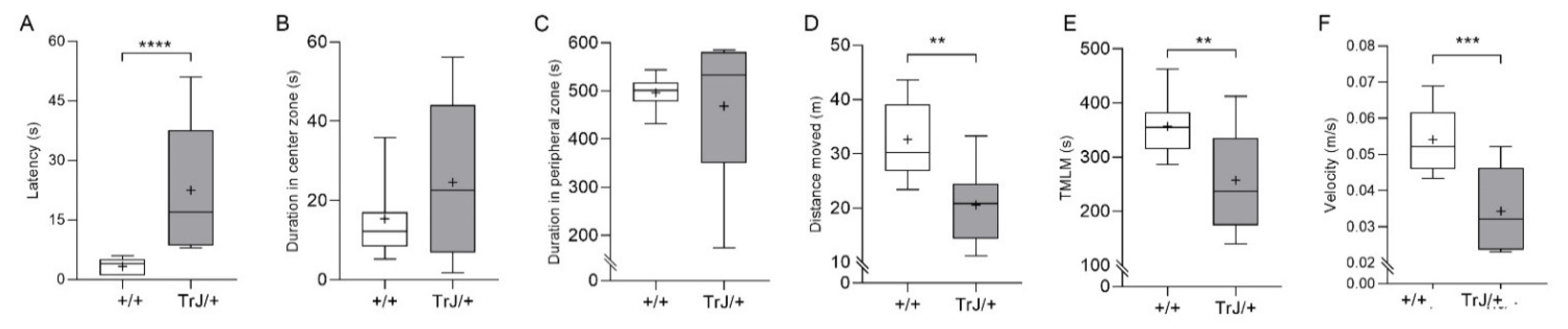
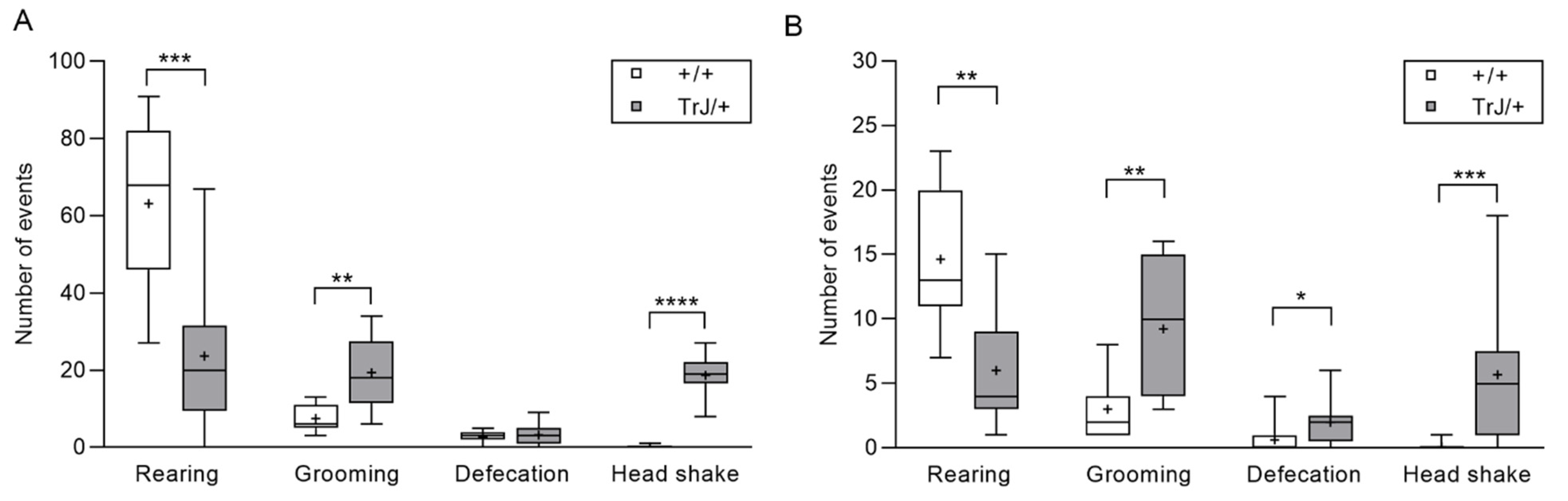
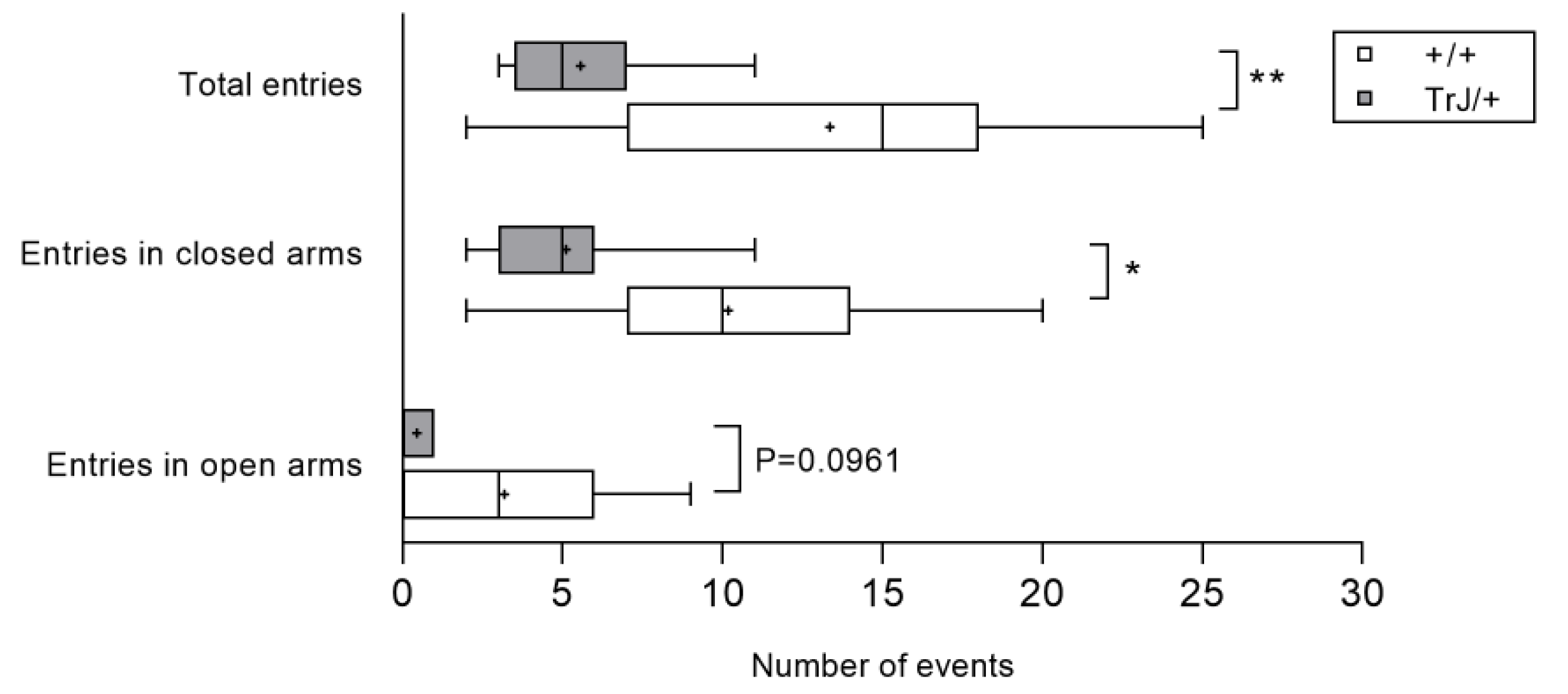
3.1. Behavior in Anxiety Tests Reveal Phenotypic Differences between TrJ/+ and +/+
- Open field test
- Elevated plus maze test
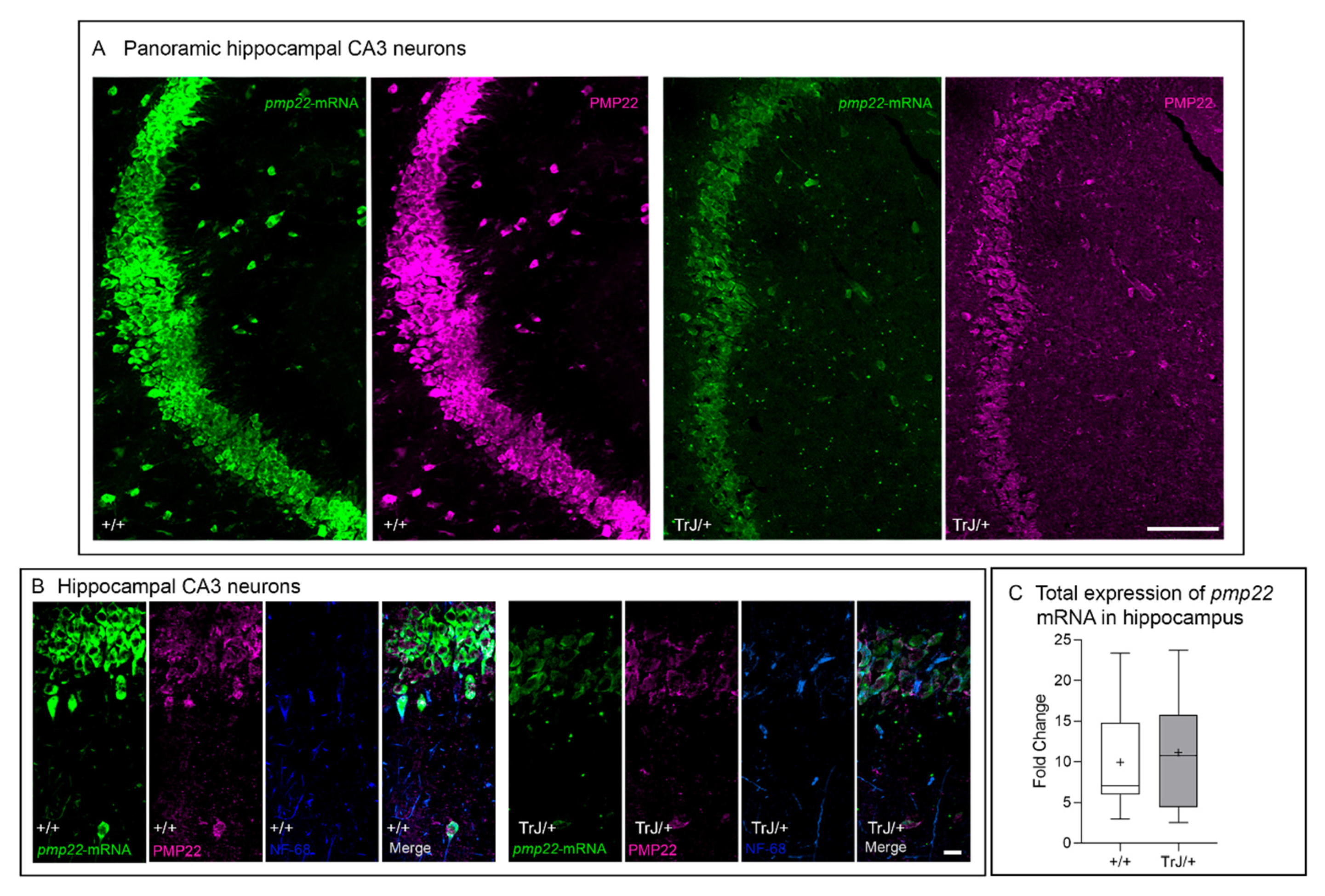
3.2. Pmp22 mRNA and PMP22 Are Expressed in Murine Hippocampal CA3 Neurons
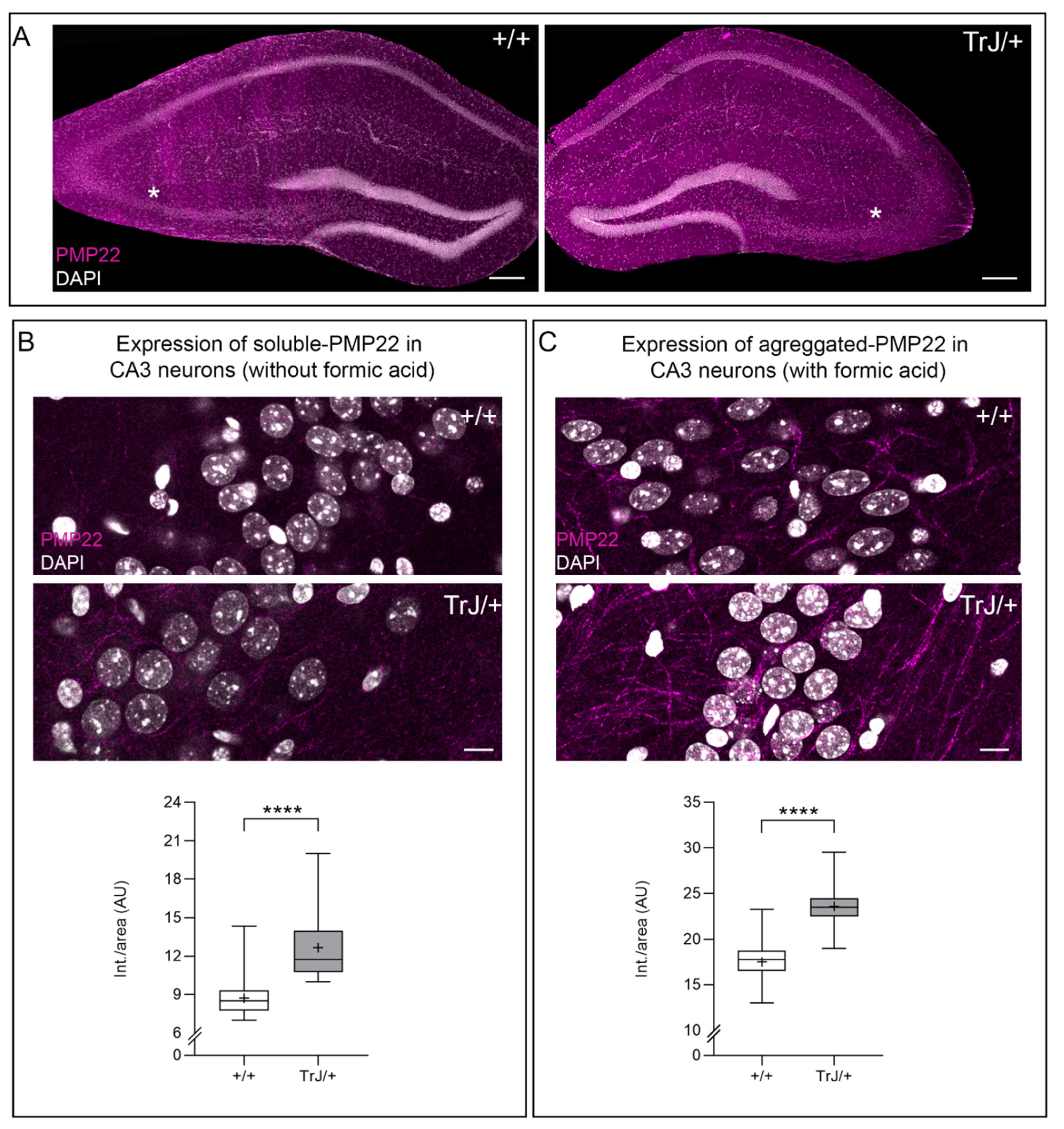
3.3. Soluble and Aggregates PMP22 Are Present in Adult Hippocampal CA3 Neurons
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kamholz, J.; Menichella, D.; Jani, A.; Garbern, J.; Lewis, R.A.; Krajewski, K.M.; Lilien, J.; Scherer, S.S.; Shy, M.E. Charcot-Marie-Tooth disease type 1. Molecular pathogenesis to gene therapy. Brain 2000, 123, 222–233. [Google Scholar] [CrossRef] [Green Version]
- Pareyson, D.; Marchesi, C. Diagnosis, natural history, and management of Charcot-Marie-Tooth disease. Lancet Neurol. 2009, 8, 654–667. [Google Scholar] [CrossRef]
- Verhamme, C.; Van Schaik, I.N.; Koelman, J.H.T.M.; De Haan, R.J.; De Visser, M. The natural history of Charcot-Marie-Tooth type 1A in adults: A 5-year follow-up study. Brain 2009, 132, 3252–3262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watila, M.M.; Balarabe, S.A. Molecular and clinical features of inherited neuropathies due to PMP22 duplication. J. Neurol. Sci. 2015, 355, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Juneja, M.; Burns, J.; Saporta, M.A.; Timmerman, V. Challenges in modelling the Charcot-Marie-Tooth neuropathies for therapy development. J. Neurol. Neurosurg. Psychiatry 2019, 90, 58–67. [Google Scholar] [CrossRef]
- Li, J.; Parker, B.; Martyn, C.; Natarajan, C.; Guo, J. The PMP22 gene and its related diseases. Mol. Neurobiol. 2013, 47, 673–698. [Google Scholar] [CrossRef] [Green Version]
- Bird, T.D. Charcot-Marie-Tooth (CMT) Hereditary Neuropathy Overview; Adam, M.P., Ardinger, H.H., Pagon, R.A., Wallace, S.E., Bean, L.J.H., Mirzaa, G., Amemiya, A., Eds.; GeneReviews®; University of Washington: Seattle, WA, USA, 1998. [Google Scholar]
- Jouaud, M.; Mathis, S.; Richard, L.; Lia, A.S.; Magy, L.; Vallat, J.M. Rodent models with expression of PMP22: Relevance to dysmyelinating CMT and HNPP. J. Neurol. Sci. 2019, 398, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Snipes, G.J.; Suter, U.; Welcher, A.A.; Shooter, E.M. Characterization of a novel peripheral nervous system myelin protein (PMP- 22/SR13). J. Cell Biol. 1992, 117, 225–238. [Google Scholar] [CrossRef]
- Parmantier, E.; Cabon, F.; Braun, C.; D’Urso, D.; Müller, H.W.; Zalc, B. Peripheral Myelin Protein-22 is Expressed in Rat and Mouse Brain and Spinal Cord Motoneurons. Eur. J. Neurosci. 1995, 7, 1080–1088. [Google Scholar] [CrossRef]
- Mobley, C.K.; Myers, J.K.; Hadziselimovic, A.; Ellis, C.D.; Sanders, C.R. Purification and initiation of structural characterization of human peripheral myelin protein 22, an integral membrane protein linked to peripheral neuropathies. Biochemistry 2007, 46, 11185–11195. [Google Scholar] [CrossRef]
- Myers, J.K.; Mobley, C.K.; Sanders, C.R. The Peripheral Neuropathy-Linked Trembler and Trembler-J Mutant Forms of Peripheral Myelin Protein 22 are Folding- Destabilized. Biochemistry 2008, 47, 10620–10629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spreyer, P.; Kuhn, H.G.; Hanemann, C.O.; Gillen, C.P.; Schaal, H.; Kuhn, R.; Lemke, G.; Müller, H.W. Axon-regulated expression of a Schwann cell transcript that is homologous to a “growth arrest-specific” gene. EMBO 1991, 10, 3661–3668. [Google Scholar] [CrossRef]
- Welcher, A.A.; Suter, U.; De Leon, M.; Snipes, G.J.; Shooter, E.M. A myelin protein is encoded by the homologue of a growth arrest-specific gene. Proc. Natl. Acad. Sci. USA. 1991, 88, 7195–7199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosse, F.; Zoidl, G.; Wilms, S.; Gillen, C.P.; Kuhn, H.G.; Müller, H.W. Differential expression of two mRNA species indicates a dual function of peripheral myelin protein PMP22 in cell growth and myelination. J. Neurosci. Res. 1994, 37, 529–537. [Google Scholar] [CrossRef]
- Sacco, S.; Totaro, R.; Bastianello, S.; Marini, C.; Carolei, A. Brain white matter lesions in an Italian family with Charcot-Marie-Tooth disease. Eur. Neurol. 2004, 51, 168–171. [Google Scholar] [CrossRef] [PubMed]
- Ohsawa, Y.; Murakami, T.; Miyazaki, Y.; Shirabe, T.; Sunada, Y. Peripheral myelin protein 22 is expressed in human central nervous system. J. Neurol. Sci. 2006, 247, 11–15. [Google Scholar] [CrossRef]
- Xie, C.; Zhou, X.; Zhu, D.; Liu, W.; Wang, X.; Yang, H.; Li, Z.; Hao, Y.; Zhang, G.X.; Guan, Y. CNS involvement in CMTX1 caused by a novel connexin 32 mutation: A 6-year follow-up in neuroimaging and nerve conduction. Neurol. Sci. 2016, 37, 1063–1070. [Google Scholar] [CrossRef]
- Norreel, J.C.; Jamon, M.; Riviere, G.; Passage, E.; Fontés, M.; Clarac, F. Behavioural profiling of a murine Charcot-Marie-Tooth disease type 1A model. Eur. J. Neurosci. 2001, 13, 1625–1634. [Google Scholar] [CrossRef] [PubMed]
- Huxley, C.; Passage, E.; Manson, A.; Putzu, G.; Figarella-Branger, D.; Pellissier, J.F.; Fontés, M. Construction of a mouse model of Charcot-Marie-Tooth disease type 1A by pronuclear injection of human YAC DNA. Hum. Mol. Genet. 1996, 5, 563–569. [Google Scholar] [CrossRef] [Green Version]
- Fridman, V.; Bundy, B.; Reilly, M.M.; Pareyson, D.; Bacon, C.; Burns, J.; Day, J.; Feely, S.; Finkel, R.S.; Grider, T.; et al. CMT subtypes and disease burden in patients enrolled in the Inherited Neuropathies Consortium natural history study: A cross-sectional analysis. J. Neurol. Neurosurg. Psychiatry 2015, 86, 873–878. [Google Scholar] [CrossRef] [Green Version]
- Suter, U.; Moskow, J.J.; Welcher, A.A.; Snipes, G.J.; Kosaras, B.; Sidman, R.L.; Buchberg, A.M.; Shooter, E.M. A leucine-to-proline mutation in the putative first transmembrane domain of the 22-kDa peripheral myelin protein in the trembler-J mouse. Proc. Natl. Acad. Sci. USA 1992, 89, 4382–4386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quarles, R.H.; Macklin, W.B.; Morell, P. Myelin Structure and Biochemistry. In Basic Neurochemistry; Siegel, G.J., Albers, R.W., Brady, S.T., Price, D.L., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2006; pp. 51–71. ISBN 9780123749475. [Google Scholar]
- Colby, J.; Nicholson, R.; Dickson, K.M.; Orfali, W.; Naef, R.; Suter, U.; Snipes, G.J. Pmp22 carrying the trembler or trembler-J mutation is intracellularly retained in myelinating Schwann cells. Neurobiol. Dis. 2000, 7, 561–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, Y.; Hirokawa, N. Mouse models of Charcot-Marie-Tooth disease. Trends Genet. 2002, 18, 39–44. [Google Scholar] [CrossRef]
- Bouhy, D.; Timmerman, V. Animal models and therapeutic prospects for Charcot-Marie-Tooth disease. Laryngoscope 2014, 2–31. [Google Scholar] [CrossRef]
- Valentijn, L.J.; Baas, F.; Wolterman, R.A.; Hoogendijk, J.E.; van den Bosch, N.H.A.; Zorn, I.; Gabreëls-Festen, A.A.W.M.; de Visser, M.; Bolhuis, P.A. Identical point mutations of PMP-22 in Trembler-J mouse and Charcot-Marie-Tooth disease type 1A. Nat. Genet. 1992, 2, 288–291. [Google Scholar] [CrossRef] [PubMed]
- Robertson, A.M.; Huxley, C.; King, R.H.M.; Thomas, P.K. Development of early postnatal peripheral nerve abnormalities in Trembler-J and PMP22 transgenic mice. J. Anat. 1999, 195, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Henry, E.W.; Sidman, R.L. The murine mutation trembler-j: Proof of semidominant expression by use of the linked vestigial tail marker. J. Neurogenet. 1983, 1, 39–52. [Google Scholar] [CrossRef]
- Henry, E.W.; Cowen, J.S.; Sidman, R.L. Comparison of trembler and trembler-j mouse phenotypes: Varying severity of peripheral hypomyelination. J. Neuropathol. Exp. Neurol. 1983, 42, 688–706. [Google Scholar] [CrossRef]
- Rosso, G.; Cal, K.; Canclini, L.; Damián, J.P.; Ruiz, P.; Rodríguez, H.; Sotelo, J.R.; Vazquez, C.; Kun, A. Early phenotypical diagnoses in Trembler-J mice model. J. Neurosci. Methods 2010, 190, 14–19. [Google Scholar] [CrossRef]
- Carola, V.; D’Olimpio, F.; Brunamonti, E.; Mangia, F.; Renzi, P. Evaluation of the elevated plus-maze and open-field tests for the assessment of anxiety-related behaviour in inbred mice. Behav. Brain Res. 2002, 134, 49–57. [Google Scholar] [CrossRef]
- Forkman, B.; Boissy, A.; Meunier-Salaün, M.C.; Canali, E.; Jones, R.B. A critical review of fear tests used on cattle, pigs, sheep, poultry and horses. Physiol. Behav. 2007, 92, 340–374. [Google Scholar] [CrossRef] [Green Version]
- Walf, A.A.; Frye, C.A. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat. Protoc. 2007, 2, 322–328. [Google Scholar] [CrossRef] [Green Version]
- Sestakova, N.; Puzserova, A.; Kluknavsky, M.; Bernatova, I. Determination of motor activity and anxiety-related behaviour in rodents: Methodological aspects and role of nitric oxide. Interdiscip. Toxicol. 2013, 6, 126–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damián, J.P.; Acosta, V.; Da Cuña, M.; Ramírez, I.; Oddone, N.; Zambrana, A.; Bervejillo, V.; Benech, J.C. Effect of resveratrol on behavioral performance of streptozotocin-induced diabetic mice in anxiety tests. Exp. Anim. 2014, 63, 277–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damián, J.P.; de Soto, L.; Espindola, D.; Gil, J.; van Lier, E. Intranasal oxytocin affects the stress response to social isolation in sheep. Physiol. Behav. 2021, 230, 113282. [Google Scholar] [CrossRef]
- Canales, J.J. Adult neurogenesis in the hippocampus-Health, Psychopathology and Brain Disease; Academic P.; Elsevier Inc.: Amsterdam, The Netherlands, 2016; ISBN 978-0-12-801977-1. [Google Scholar]
- Hannula, D.E.; Duff, M.C. The Hippocampus from Cells to Systems: Structure, Connectivity, and Functional Contributions to Memory and Flexible Cognition; Hannula, D.E., Duff, M.C., Eds.; Springer International Publishing: Cham, Switzerland, 2017; ISBN 978-3-319-50406-3. [Google Scholar]
- Fan, J.; Guang, H.; Zhang, H.; Chen, D.; Ding, L.; Fan, X.; Xue, F.; Gan, Z.; Wang, Y.; Mao, S.; et al. SIRT1 Mediates Apelin-13 in Ameliorating Chronic Normobaric Hypoxia-induced Anxiety-like Behavior by Suppressing NF-κB Pathway in Mice Hippocampus. Neuroscience 2018, 381, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Żmudzka, E.; Sałaciak, K.; Sapa, J.; Pytka, K. Serotonin receptors in depression and anxiety: Insights from animal studies. Life Sci. 2018, 210, 106–124. [Google Scholar] [CrossRef]
- Zhang, W.J.; Cao, W.Y.; Huang, Y.Q.; Cui, Y.H.; Tu, B.X.; Wang, L.F.; Zou, G.J.; Liu, Y.; Hu, Z.L.; Hu, R.; et al. The Role of miR-150 in Stress-Induced Anxiety-Like Behavior in Mice. Neurotox. Res. 2019, 35, 160–172. [Google Scholar] [CrossRef] [PubMed]
- DeLuca, R.R.; Alexandre, S.R.; Marques, T.; Lopez de Souza, N.; Merusse, J.L.B.; Neves, S.P. Manual para técnicos em bioterismo. In Manual Para Técnicos em Bioterismo; Winner Graph: São Paulo, Brazil, 1996; p. 259. [Google Scholar]
- Fraga, M.C.; Moura, E.G.; Silva, J.O.; Bonomo, I.T.; Filgueiras, C.C.; Abreu-Villaça, Y.; Passos, M.C.F.; Lisboa, P.C.; Manhães, A.C. Maternal prolactin inhibition at the end of lactation affects learning/memory and anxiety-like behaviors but not novelty-seeking in adult rat progeny. Pharmacol. Biochem. Behav. 2011, 100, 165–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kun, A.; Canclini, L.; Rosso, G.; Bresque, M.; Romeo, C.; Hanusz, A.; Cal, K.; Calliari, A.; Sotelo Silveira, J.; Sotelo, J.R. F-actin distribution at nodes of Ranvier and Schmidt-Lanterman incisures in mammalian sciatic nerves. Cytoskeleton 2012, 69, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Kun, A.; Rosso, G.; Canclini, L.; Bresque, M.; Romeo, C.; Cal, K.; Calliari, A.; Hanuz, A.; Roberto, J.; Roberto, J. The Schwann Cell-Axon Link in Normal Condition or Neuro-Degenerative Diseases: An Immunocytochemical Approach. In Applications of Immunocytochemistry; Dehghani, H., Ed.; InTech: London, Switzerland, 2012; pp. 249–266. ISBN 978-953-51-5235-4. [Google Scholar]
- Olivera, S.; Rodriguez-Ithurralde, D.; Henley, J.M. Acetylcholinesterase promotes neurite elongation, synapse formation, and surface expression of AMPA receptors in hippocampal neurones. Mol. Cell. Neurosci. 2003, 23, 96–106. [Google Scholar] [CrossRef] [Green Version]
- Canclini, L.; Farias, J.; Di Paolo, A.; Sotelo-Silveira, J.R.; Folle, G.; Kun, A.; Sotelo, J.R. Association of microtubules and axonal RNA transferred from myelinating Schwann cells in rat sciatic nerve. PLoS ONE 2020, 15, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Harro, J. Animals, anxiety, and anxiety disorders: How to measure anxiety in rodents and why. Behav. Brain Res. 2018, 352, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Bhat, Z.A.; Kumar, D. Animal models of anxiety: A comprehensive review. J. Pharmacol. Toxicol. Methods 2013, 68, 175–183. [Google Scholar] [CrossRef]
- Ennaceur, A. Tests of unconditioned anxiety—Pitfalls and disappointments. Physiol. Behav. 2014, 135, 55–71. [Google Scholar] [CrossRef]
- Hart, P.C.; Bergner, C.L.; Smolinsky, A.N.; Dufour, B.D.; Egan, R.J.; LaPorte, J.L.; Kalueff, A.V. Experimental models of anxiety for drug discovery and brain research. In Mouse Models for Drug Discovery-Methods in Molecular Biology (Methods and Protocols); Proetzel, G., Wiles, M., Eds.; Humana Press Inc.: Cham, Switzerland, 2010; Volume 602, pp. 299–321. ISBN 978-1-60761-058-8. [Google Scholar]
- Chad Hoyle, J.; Isfort, M.C.; Roggenbuck, J.; David Arnold, W. The genetics of Charcot–Marie–Tooth disease: Current trends and future implications for diagnosis and management. Appl. Clin. Genet. 2015, 8, 235–243. [Google Scholar] [CrossRef] [Green Version]
- McGrath, M.C. Charcot-Marie-Tooth 1A: A narrative review with clinical and anatomical perspectives. Clin. Anat. 2016, 29, 547–554. [Google Scholar] [CrossRef]
- Suter, U.; Welcher, A.A.; Özcelik, T.; Snipes, G.J.; Kosaras, B.; Francke, U.; Billings-Gagliardi, S.; Sidman, R.L.; Shooter, E.M. Trembler mouse carries a point mutation in a myelin gene. Nature 1992, 356, 241–244. [Google Scholar] [CrossRef]
- Notterpek, L.; Tolwani, R.J. Experimental models of peripheral neuropathies. Lab. Anim. Sci. 1999, 49, 588–599. [Google Scholar]
- Cornwell-Jones, C.A.; Palfai, T.; Krasenbaum, D.; Byer, E.; Clark, R.; Kinnard, K. Housing influences exploration and social interaction of control and DSP-4-treated rats. Physiol. Behav. 1992, 52, 271–276. [Google Scholar] [CrossRef]
- To, C.T.; Bagdy, G. Anxiogenic effect of central CCK administration is attenuated by chronic fluoxetine or ipsapirone treatment. Neuropharmacology 1999, 38, 279–282. [Google Scholar] [CrossRef]
- Hawkins, M.F.; Uzelac, S.M.; Hearn, J.K.; Baumeister, A.A. Effects of selective serotonin2 ligands on behaviors evoked by stress in the rat. Pharmacol. Biochem. Behav. 2008, 90, 632–639. [Google Scholar] [CrossRef]
- Leggio, G.M.; Micale, V.; Le Foll, B.; Mazzola, C.; Nobrega, J.N.; Drago, F. Dopamine D3 receptor knock-out mice exhibit increased behavioral sensitivity to the anxiolytic drug diazepam. Eur. Neuropsychopharmacol. 2011, 21, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Deacon, R.M.J.; Rawlins, J.N.P. Hippocampal lesions, species-typical behaviours and anxiety in mice. Behav. Brain Res. 2005, 156, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Deacon, R.M.J.; Croucher, A.; Rawlins, J.N.P. Hippocampal cytotoxic lesion effects on species-typical behaviours in mice. Behav. Brain Res. 2002, 132, 203–213. [Google Scholar] [CrossRef]
- Gispen, W.H.; Isaacson, R.L. ACTH-induced excessive grooming in the rat. Pharmacol. Ther. 1981, 12, 209–246. [Google Scholar] [CrossRef] [Green Version]
- Colbern, D.; Isaacson, R.L.; Bohus, B.; Gispen, W.H. Limbic-midbrain lesions and acth-induced excessive grooming. Life Sci. 1977, 21, 393–401. [Google Scholar] [CrossRef] [Green Version]
- Oades, R.; Isaacson, R.L. p-Chlorophenylalanine-produced effects on behavior in intact and brain-damaged rats. Behav. Biol. 1977, 20, 500–506. [Google Scholar] [CrossRef] [Green Version]
- Moody, T.W.; Merali, Z.; Crawley, J.N. The Effects of Anxiolytics and Other Agents on Rat Grooming Behavior. Ann. N. Y. Acad. Sci. 1988, 525, 281–290. [Google Scholar] [CrossRef]
- Sachs, B.D. The Development of Grooming and Its Expression in Adult Animals. Ann. N. Y. Acad. Sci. 1988, 525, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Kalueff, A.V.; Tuohimaa, P. Experimental modeling of anxiety and depression. Acta Neurobiol. Exp. 2004, 64, 439–448. [Google Scholar]
- Le Foll, B.; Diaz, J.; Sokoloff, P. Neuroadaptations to hyperdopaminergia in dopamine D3 receptor-deficient mice. Life Sci. 2005, 76, 1281–1296. [Google Scholar] [CrossRef]
- Beninger, R.J.; Mazurski, E.J.; Hoffman, D.C. Receptor subtype-specific dopaminergic agents and unconditioned behavior. Pol. J. Pharmacol. Pharm. 1991, 43, 507–528. [Google Scholar] [PubMed]
- Chinen, C.C.; Frussa-Filho, R. Conditioning to injection procedures and repeated testing increase SCH 23390-induced catalepsy in mice. Neuropsychopharmacology 1999, 21, 670–678. [Google Scholar] [CrossRef] [Green Version]
- Gewirtz, J.C.; Marek, G.J. Behavioral evidence for interactions between a hallucinogenic drug and group II metabotropic glutamate receptors. Neuropsychopharmacology 2000, 23, 569–576. [Google Scholar] [CrossRef]
- Peroutka, S.J.; Lebovitz, R.M.; Snyder, S.H. Two distinct central serotonin receptors with different physiological functions. Science 1981, 212, 827–829. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Fattah, M.; Matsumoto, K.; Gammaz, H.A.K.; Watanabe, H. Hypothermic effect of harmala alkaloid in rats: Involvement of serotonergic mechanism. Pharmacol. Biochem. Behav. 1995, 52, 421–426. [Google Scholar] [CrossRef]
- Van Oekelen, D.; Luyten, W.H.M.L.; Leysen, J.E. 5-HT2A and 5-HT2C receptors and their atypical regulation properties. Life Sci. 2003, 72, 2429–2449. [Google Scholar] [CrossRef]
- Islam, A.; Thompson, K.S.J.; Akhtar, S.; Handley, S.L. Increased 5-HT2A receptor expression and function following central glucocorticoid receptor knockdown in vivo. Eur. J. Pharmacol. 2004, 502, 213–220. [Google Scholar] [CrossRef]
- De Gasperi, R.; Gama Sosa, M.A.; Naumowicz, Z.; Hof, P.R.; Notterpek, L.; Davis, K.L.; Buxbaum, J.D.; Elder, G.S. Peripheral mylein protein-22 is expressed in CNS myelin. Transl. Neurosci. 2010, 1, 282–285. [Google Scholar] [CrossRef] [Green Version]
- Zeier, Z.; Madorsky, I.; Xu, Y.; Ogle, W.O.; Notterpek, L.; Foster, T.C. Gene expression in the Hippocampus: Regionally Specific Effects of Aging and Caloric Restriction. Mech. Ageing Dev. 2011, 132, 8–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Notterpek, L. Dietary restriction supports peripheral nerve health by enhancing endogenous protein quality control mechanisms. Exp. Gerontol. 2013, 48, 1085–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekins, S.; Litterman, N.K.; Arnold, R.J.G.; Burgess, R.W.; Freundlich, J.S.; Gray, S.J.; Higgins, J.J.; Langley, B.; Willis, D.E.; Notterpek, L.; et al. A brief review of recent Charcot-Marie-Tooth research and priorities. F1000Research 2015, 4, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.C.; Tsai, P.C.; Lin, T.S.; Hsiao, C.T.; Chao, N.C.; Lin, K.P.; Lee, Y.C. Clinical and Molecular Characterization of PMP22 point mutations in Taiwanese patients with Inherited Neuropathy. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Miles, J.R.; Tavori, H.; Lin, M.; Khoshbouei, H.; Borchelt, D.R.; Bazick, H.; Landreth, G.E.; Lee, S.; Fazio, S.; et al. PMP22 regulates cholesterol trafficking and ABCA1-mediated cholesterol efflux. J. Neurosci. 2019, 39, 5404–5418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantera, H.; Shy, M.E.; Svaren, J. Regulating PMP22 expression as a dosage sensitive neuropathy gene. Brain Res. 2020, 1726. [Google Scholar] [CrossRef] [PubMed]
- Manfioletti, G.; Ruaro, M.E.; Del Sal, G.; Philipson, L.; Schneider, C. A growth arrest-specific (gas) gene codes for a membrane protein. Mol. Cell. Biol. 1990, 10, 2924–2930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.; King, R.M.; Philipson, L. Genes specifically expressed at growth arrest of mammalian cells. Cell 1988, 54, 787–793. [Google Scholar] [CrossRef]
- Rosso, G.; Negreira, C.; Sotelo, J.R.; Kun, A. Myelinating and demyelinating phenotype of Trembler-J mouse (a model of Charcot-Marie-Tooth human disease) analyzed by atomic force microscopy and confocal microscopy. J. Mol. Recognit. 2012, 25, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Fabbretti, E.; Edomi, P.; Brancolini, C.; Schneider, C. Apoptotic phenotype induced by overexpression of wild-type gas3/PMP22: Its relation to the demyelinating peripheral neuropathy CMT1A. Genes Dev. 1995, 9, 1846–1856. [Google Scholar] [CrossRef] [Green Version]
- Suter, U.; Snipes, G.J.; Schoener-Scott, R.; Welcher, A.A.; Pareek, S.; Lupski, J.R.; Murphy, R.A.; Shooter, E.M.; Patel, P.I. Regulation of tissue-specific expression of alternative peripheral myelin protein-22 (PMP22) gene transcripts by two promoters. J. Biol. Chem. 1994, 269, 25795–25808. [Google Scholar] [CrossRef]
- Suter, U.; Welcher, A.A.; Jackson Snipes, G. Progress in the molecular understanding of hereditary peripheral neuropathies reveals new insights into the biology of the peripheral nervous system. Trends Neurosci. 1993, 16, 50–56. [Google Scholar] [CrossRef]
- Jessen, K.R.; Mirsky, R. Schwann cell precursors and their development. Glia 1991, 4, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Attardi, L.D.; Reczek, E.E.; Cosmas, C.; Demicco, E.G.; McCurrach, M.E.; Lowe, S.W.; Jacks, T. Erratum: PERP, an apoptosis-associated target of p53, is a novel member of the PMP-22/gas3 family. Genes Dev. 2000, 14, 1835. [Google Scholar]
- Parmantier, E.; Braun, C.; Thomas, J.L.; Peyron, F.; Martinez, S.; Zalc, B. PMP-22 expression in the central nervous system of the embryonic mouse defines potential transverse segments and longitudinal columns. J. Comp. Neurol. 1997, 378, 159–172. [Google Scholar] [CrossRef]
- Thomas, P.K.; Marques, W.; Davis, M.B.; Sweeney, M.G.; King, R.H.M.; Bradley, J.L.; Muddle, J.R.; Tyson, J.; Malcolm, S.; Harding, A.E. The phenotypic manifestations of chromosome 17p11.2 duplication. Brain 1997, 120, 465–478. [Google Scholar] [CrossRef] [Green Version]
- Chanson, J.B.; Echaniz-Laguna, A.; Blanc, F.; Lacour, A.; Ballonzoli, L.; Kremer, S.; Namer, I.J.; Lannes, B.; Tranchant, C.; Vermersch, P.; et al. Central nervous system abnormalities in patients with PMP22 gene mutations: A prospective study. J. Neurol. Neurosurg. Psychiatry 2013, 84, 392–397. [Google Scholar] [CrossRef]
- Brandt, A.U.; Meinert-Bohn, E.; Rinnenthal, J.L.; Zimmermann, H.; Mikolajczak, J.; Oberwahrenbrock, T.; Papazoglou, S.; Pfüller, C.F.; Schinzel, J.; Tackenberg, B.; et al. Afferent visual pathway affection in patients with PMP22 deletion-related hereditary neuropathy with liability to pressure palsies. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damián, J.P.; Vázquez Alberdi, L.; Canclini, L.; Rosso, G.; Bravo, S.O.; Martínez, M.; Uriarte, N.; Ruiz, P.; Calero, M.; Di Tomaso, M.V.; et al. Central Alteration in Peripheral Neuropathy of Trembler-J Mice: Hippocampal pmp22 Expression and Behavioral Profile in Anxiety Tests. Biomolecules 2021, 11, 601. https://doi.org/10.3390/biom11040601
Damián JP, Vázquez Alberdi L, Canclini L, Rosso G, Bravo SO, Martínez M, Uriarte N, Ruiz P, Calero M, Di Tomaso MV, et al. Central Alteration in Peripheral Neuropathy of Trembler-J Mice: Hippocampal pmp22 Expression and Behavioral Profile in Anxiety Tests. Biomolecules. 2021; 11(4):601. https://doi.org/10.3390/biom11040601
Chicago/Turabian StyleDamián, Juan Pablo, Lucia Vázquez Alberdi, Lucía Canclini, Gonzalo Rosso, Silvia Olivera Bravo, Mariana Martínez, Natalia Uriarte, Paul Ruiz, Miguel Calero, María Vittoria Di Tomaso, and et al. 2021. "Central Alteration in Peripheral Neuropathy of Trembler-J Mice: Hippocampal pmp22 Expression and Behavioral Profile in Anxiety Tests" Biomolecules 11, no. 4: 601. https://doi.org/10.3390/biom11040601