Cation Transporters of Candida albicans—New Targets to Fight Candidiasis?

 and
and
Abstract
:1. Introduction
2. Alkali and Alkali Earth Metals
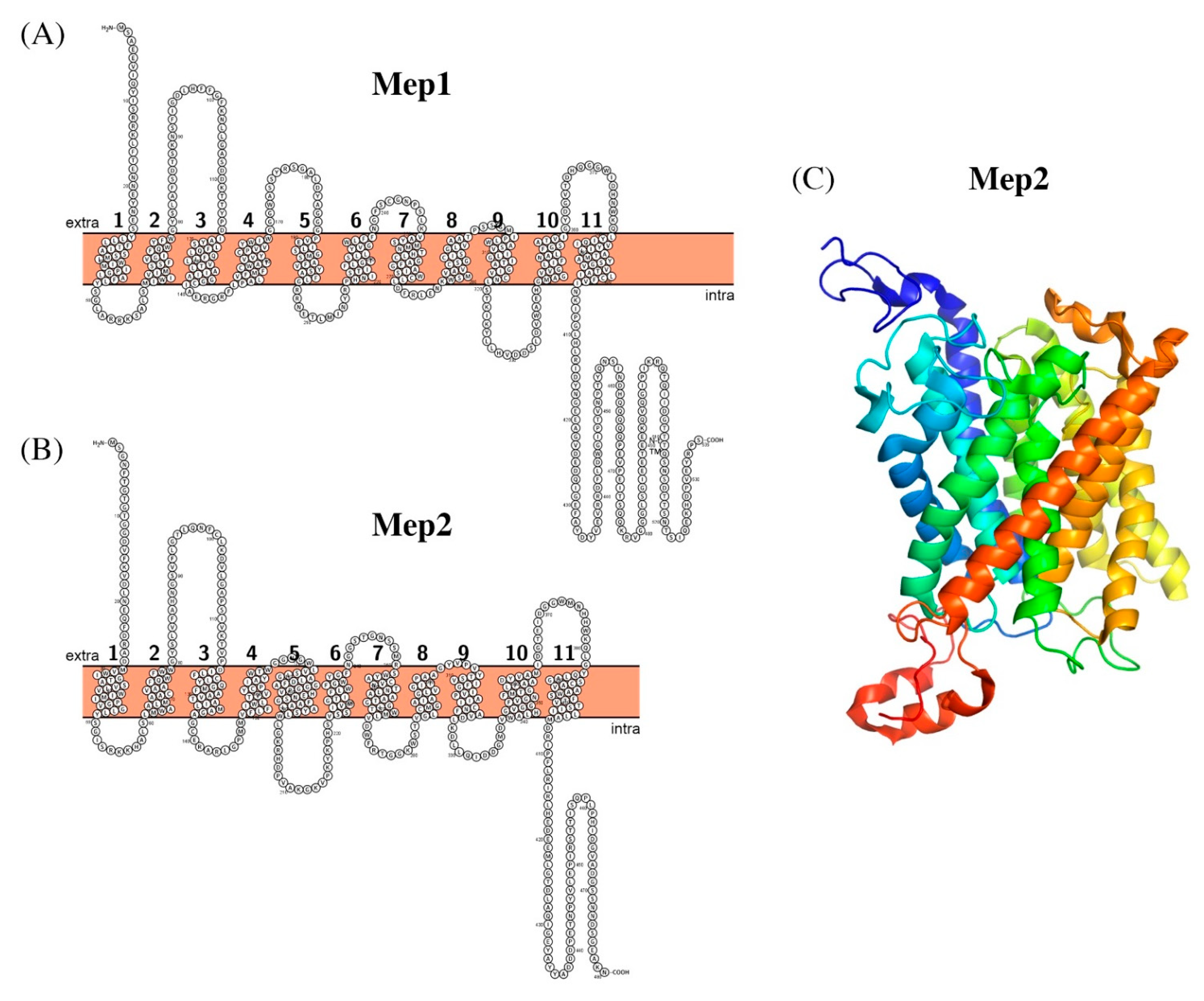
2.1. Ammonium Transporting Systems
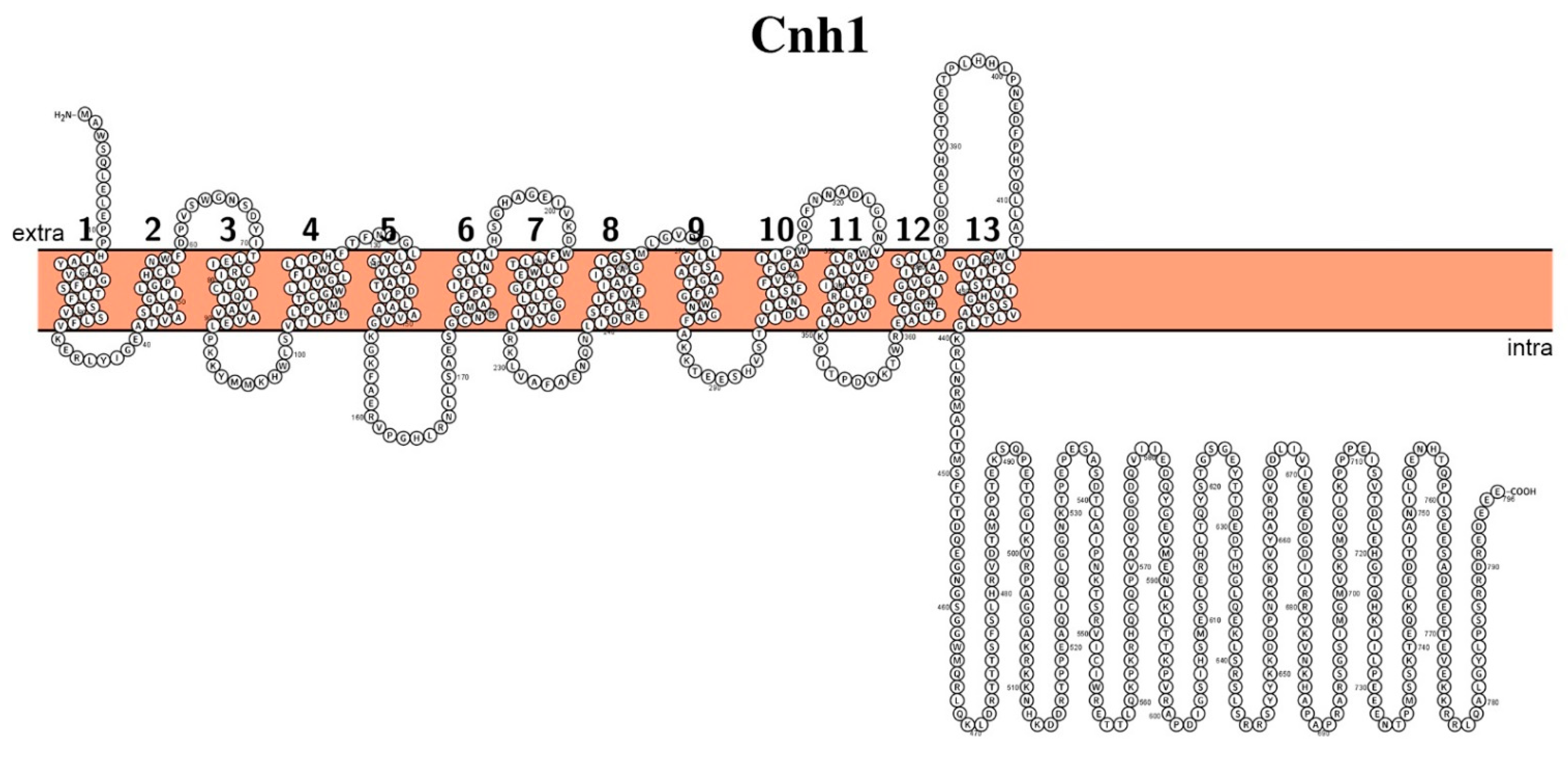
2.2. Sodium Transporting Systems
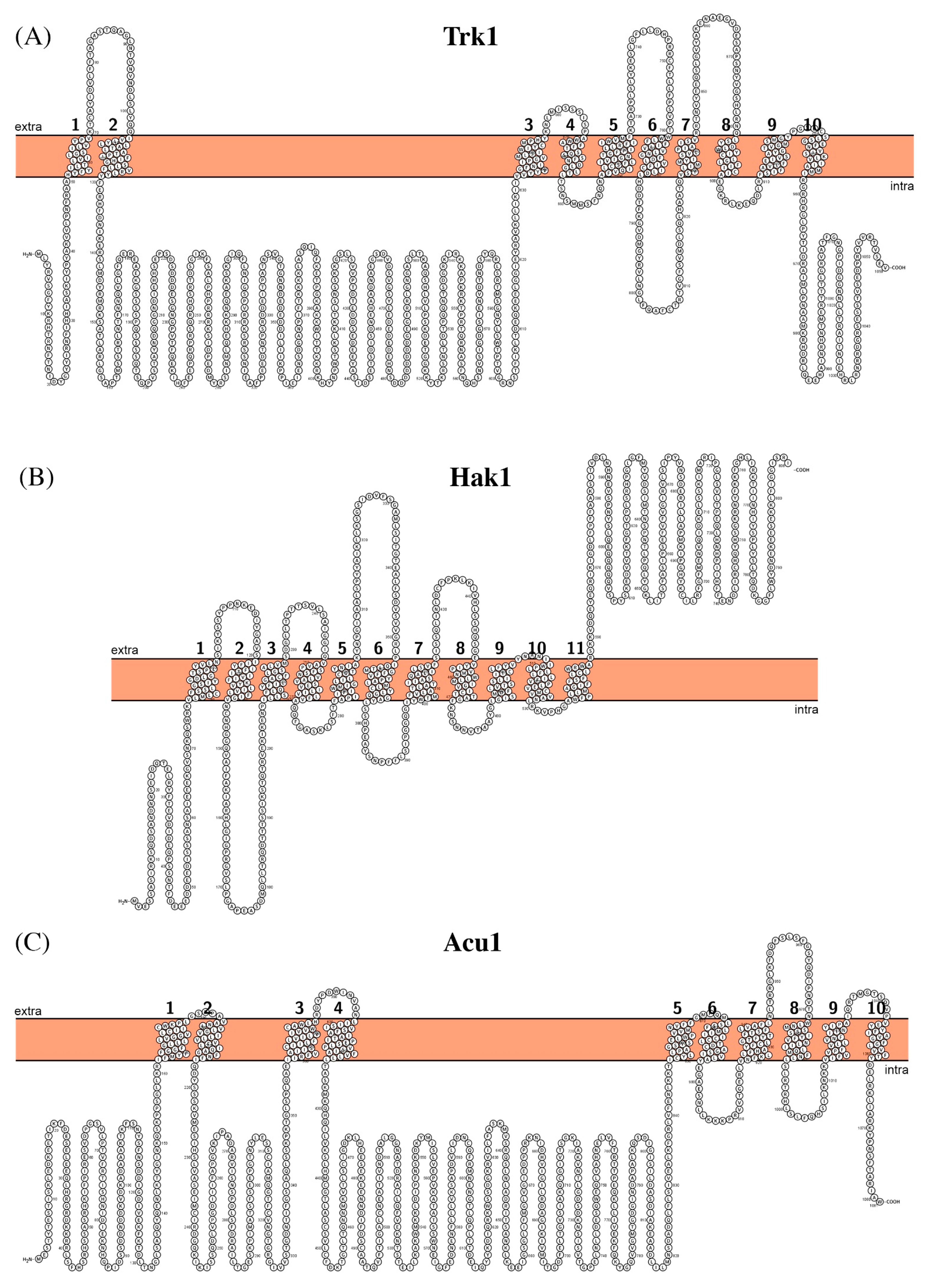
2.3. Potassium Transporting Systems
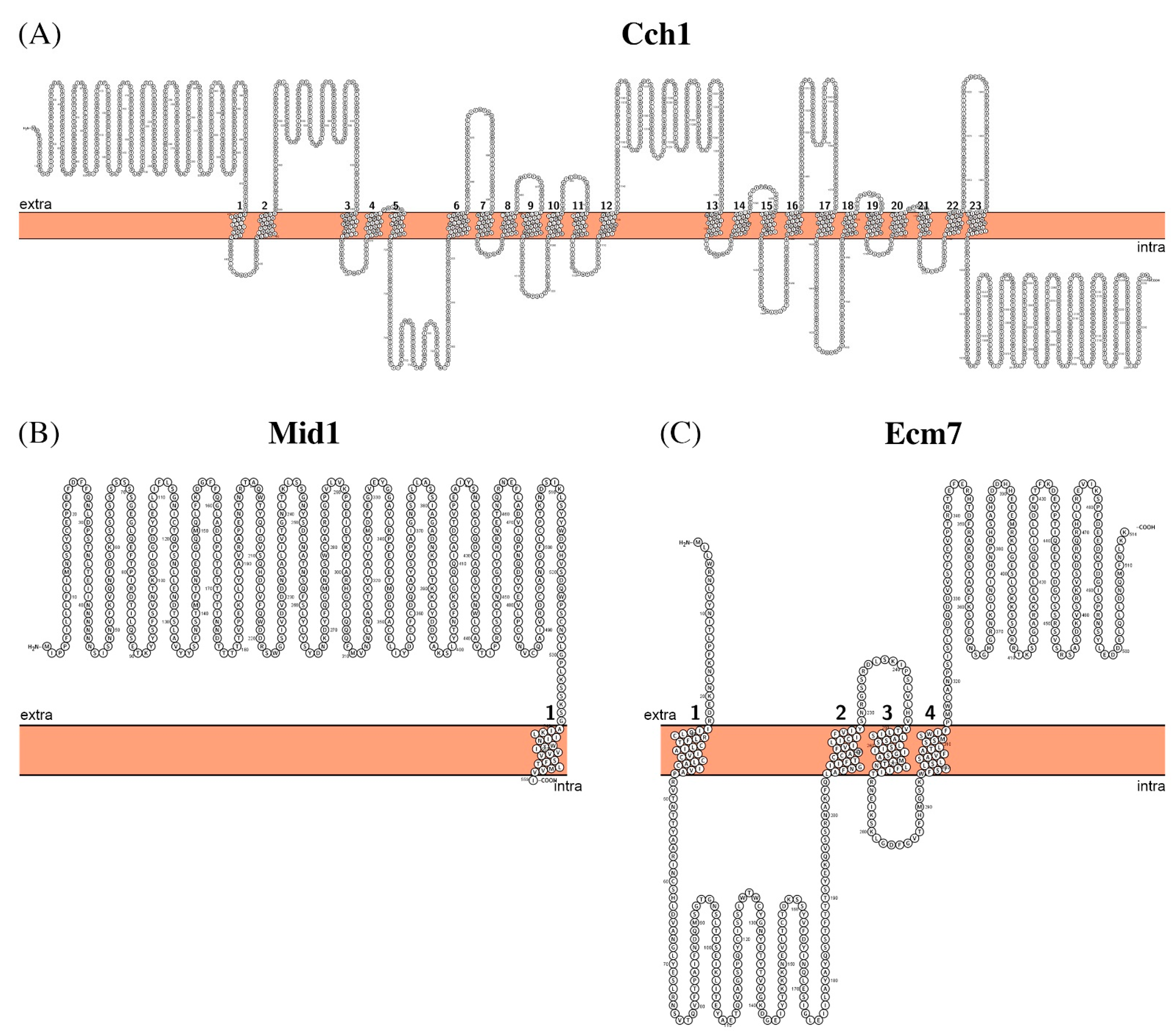
2.4. Calcium Transport Systems
3. Heavy Metals
3.1. Iron Transport Systems
3.2. Zinc Transporting Systems
3.3. Copper Transporting Systems
4. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
Methodology
References
- Gow, N.A.R.; Yadav, B. Microbe Profile: Candida albicans: A shape-changing, opportunistic pathogenic fungus of humans. Microbiology 2017, 163, 1145–1147. [Google Scholar] [CrossRef] [PubMed]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida albicans pathogenicity mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lalla, R.V.; Patton, L.L.; Dongari-Bagtzoglou, A. Oral candidiasis: Pathogenesis, clinical presentation, diagnosis and treatment strategies. J. Calif. Dent. Assoc. 2013, 41, 263–268. [Google Scholar] [PubMed]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Executive Summary: Clinical Practice Guideline for the Management of Candidiasis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2016, 62, 409–417. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.; Lewis, M. Pathogenesis and treatment of oral candidosis. J. Oral Microbiol. 2011, 3. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, B.; Ferreira, C.; Alves, C.T.; Henriques, M.; Azeredo, J.; Silva, S. Vulvovaginal candidiasis: Epidemiology, microbiology and risk factors. Crit. Rev. Microbiol. 2016, 42, 905–927. [Google Scholar] [CrossRef] [Green Version]
- Beigi, R.H.; Meyn, L.A.; Moore, D.M.; Krohn, M.A.; Hillier, S.L. Vaginal yeast colonization in nonpregnant women: A longitudinal study. Obstet. Gynecol. 2004, 104, 926–930. [Google Scholar] [CrossRef]
- Falagas, M.E.; Apostolou, K.E.; Pappas, V.D. Attributable mortality of candidemia: A systematic review of matched cohort and case-control studies. Eur. J. Clin. Microbiol. Infect. Dis. 2006, 25, 419–425. [Google Scholar] [CrossRef]
- Moran, C.; Grussemeyer, C.A.; Spalding, J.R.; Benjamin, D.K.; Reed, S.D. Candida albicans and non-albicans bloodstream infections in adult and pediatric patients: Comparison of mortality and costs. Pediatr. Infect. Dis. J. 2009, 28, 433–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benedict, K.; Jackson, B.R.; Chiller, T.; Beer, K.D. Estimation of Direct Healthcare Costs of Fungal Diseases in the United States. Clin. Infect. Dis. 2019, 68, 1791–1797. [Google Scholar] [CrossRef] [Green Version]
- Spampinato, C.; Leonardi, D. Candida infections, causes, targets, and resistance mechanisms: Traditional and alternative antifungal agents. BioMed Res. Int. 2013, 2013, 204237. [Google Scholar] [CrossRef] [Green Version]
- Kramer, K.M.; Skaar, D.J.; Ackerman, B.H. The fluconazole era: Management of hematogenously disseminated candidiasis in the nonneutropenic patient. Pharmacotherapy 1997, 17, 538–548. [Google Scholar] [PubMed]
- Laniado-Laborín, R.; Cabrales-Vargas, M.N. Amphotericin B: Side effects and toxicity. Rev. Iberoam. Micol. 2009, 26, 223–227. [Google Scholar] [CrossRef]
- Cappelletty, D.; Eiselstein-McKitrick, K. The echinocandins. Pharmacotherapy 2007, 27, 369–388. [Google Scholar] [CrossRef]
- Ariz, I.; Boeckstaens, M.; Gouveia, C.; Martins, A.P.; Sanz-Luque, E.; Fernández, E.; Soveral, G.; von Wirén, N.; Marini, A.M.; Aparicio-Tejo, P.M.; et al. Nitrogen isotope signature evidences ammonium deprotonation as a common transport mechanism for the AMT-Mep-Rh protein superfamily. Sci. Adv. 2018, 4, eaar3599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morschhäuser, J. Nitrogen regulation of morphogenesis and protease secretion in Candida albicans. Int. J. Med. Microbiol. 2011, 301, 390–394. [Google Scholar] [CrossRef] [PubMed]
- Xianyan, L.; Wenyan, Z.; Zhi, Z.; Jian, C.; Du, G. Effect of H2O2 stress on glutathione production by Candida utilis. Sheng Wu Gong Cheng Xue Bao 2008, 24, 1046–1050. [Google Scholar]
- Limjindaporn, T.; Khalaf, R.A.; Fonzi, W.A. Nitrogen metabolism and virulence of Candida albicans require the GATA-type transcriptional activator encoded by GAT1. Mol. Microbiol. 2003, 50, 993–1004. [Google Scholar] [CrossRef]
- Andrade, S.L.A.; Einsle, O. The Amt/Mep/Rh family of ammonium transport proteins. Mol. Membr. Biol. 2007, 24, 357–365. [Google Scholar] [CrossRef]
- Holsbeeks, I.; Lagatie, O.; Van Nuland, A.; Van de Velde, S.; Thevelein, J.M. The eukaryotic plasma membrane as a nutrient-sensing device. Trends Biochem. Sci. 2004, 29, 556–564. [Google Scholar] [CrossRef]
- Conrad, M.; Schothorst, J.; Kankipati, H.N.; Van Zeebroeck, G.; Rubio-Texeira, M.; Thevelein, J.M. Nutrient sensing and signaling in the yeast Saccharomyces cerevisiae. FEMS Microbiol. Rev. 2014, 38, 254–299. [Google Scholar] [CrossRef] [Green Version]
- Van den Berg, B.; Chembath, A.; Jefferies, D.; Basle, A.; Khalid, S.; Rutherford, J.C. Structural basis for Mep2 ammonium transceptor activation by phosphorylation. Nat. Commun. 2016, 7, 11337. [Google Scholar] [CrossRef] [Green Version]
- Dunkel, N.; Biswas, K.; Hiller, E.; Fellenberg, K.; Satheesh, S.V.; Rupp, S.; Morschhäuser, J. Control of morphogenesis, protease secretion and gene expression in Candida albicans by the preferred nitrogen source ammonium. Microbiology 2014, 160, 1599–1608. [Google Scholar] [CrossRef]
- Biswas, K.; Morschhäuser, J. The Mep2p ammonium permease controls nitrogen starvation-induced filamentous growth in Candida albicans. Mol. Microbiol. 2005, 56, 649–669. [Google Scholar] [CrossRef]
- Marini, A.M.; Soussi-Boudekou, S.; Vissers, S.; Andre, B. A family of ammonium transporters in Saccharomyces cerevisiae. Mol. Cell. Biol. 1997, 17, 4282–4293. [Google Scholar] [CrossRef] [Green Version]
- Biswas, S.; Van Dijck, P.; Datta, A. Environmental sensing and signal transduction pathways regulating morphopathogenic determinants of Candida albicans. Microbiol. Mol. Biol. Rev. 2007, 71, 348–376. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, A.; Ganesan, K.; Datta, A. Induction of secretory acid proteinase in Candida albicans. J. Gen. Microbiol. 1991, 137, 2455–2461. [Google Scholar] [CrossRef] [Green Version]
- Csank, C.; Schröppel, K.; Leberer, E.; Harcus, D.; Mohamed, O.; Meloche, S.; Thomas, D.Y.; Whiteway, M. Roles of the Candida albicans mitogen-activated protein kinase homolog, Cek1p, in hyphal development and systemic candidiasis. Infect. Immun. 1998, 66, 2713–2721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hube, B.; Monod, M.; Schofield, D.A.; Brown, A.J.; Gow, N.A. Expression of seven members of the gene family encoding secretory aspartyl proteinases in Candida albicans. Mol. Microbiol. 1994, 14, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Ross, I.K.; De Bernardis, F.; Emerson, G.W.; Cassone, A.; Sullivan, P.A. The secreted aspartate proteinase of Candida albicans: Physiology of secretion and virulence of a proteinase-deficient mutant. J. Gen. Microbiol. 1990, 136, 687–694. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, G.; Wiltshire, C.; Macaskill, S.; Tournu, H.; Budge, S.; Brown, A.J. Gcn4 co-ordinates morphogenetic and metabolic responses to amino acid starvation in Candida albicans. EMBO J. 2002, 21, 5448–5456. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, M.C.; Heitman, J. The MEP2 ammonium permease regulates pseudohyphal differentiation in Saccharomyces cerevisiae. EMBO J. 1998, 17, 1236–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krauke, Y.; Sychrova, H. Functional comparison of plasma-membrane Na+/H+ antiporters from two pathogenic Candida species. BMC Microbiol. 2008, 8, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinclova-Zimmermannova, O.; Sychrová, H. Plasma-membrane Cnh1 Na+/H+ antiporter regulates potassium homeostasis in Candida albicans. Microbiology 2007, 153, 2603–2612. [Google Scholar] [CrossRef] [Green Version]
- Dutta, D.; Ullah, A.; Bibi, S.; Fliegel, L. Functional Analysis of Conserved Transmembrane Charged Residues and a Yeast Specific Extracellular Loop of the Plasma Membrane Na. Sci. Rep. 2019, 9, 6191. [Google Scholar] [CrossRef] [Green Version]
- Hermann, P.; Forgács, K.; Gál, E.; Lenkey, B.; Nagy, G.; Rozgonyi, F. Effects of alkali metal ions on some virulence traits of Candida albicans. Folia Microbiol. 2003, 48, 173–176. [Google Scholar] [CrossRef]
- Watanabe, H.; Azuma, M.; Igarashi, K.; Ooshima, H. Relationship between cell morphology and intracellular potassium concentration in Candida albicans. J. Antibiot. 2006, 59, 281–287. [Google Scholar] [CrossRef] [Green Version]
- Kinclová, O.; Potier, S.; Sychrová, H. The Candida albicans Na(+)/H(+) antiporter exports potassium and rubidium. FEBS Lett. 2001, 504, 11–15. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Navarro, A. Potassium transport in fungi and plants. Biochim. Biophys. Acta 2000, 1469, 1–30. [Google Scholar] [CrossRef]
- Soong, T.W.; Yong, T.F.; Ramanan, N.; Wang, Y. The Candida albicans antiporter gene CNH1 has a role in Na+ and H+ transport, salt tolerance, and morphogenesis. Microbiology 2000, 146, 1035–1044. [Google Scholar] [CrossRef] [Green Version]
- Smidova, A.; Stankova, K.; Petrvalska, O.; Lazar, J.; Sychrova, H.; Obsil, T.; Zimmermannova, O.; Obsilova, V. The activity of Saccharomyces cerevisiae Na. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 118534. [Google Scholar] [CrossRef]
- Ariño, J.; Ramos, J.; Sychrova, H. Monovalent cation transporters at the plasma membrane in yeasts. Yeast 2019, 36, 177–193. [Google Scholar] [CrossRef]
- Hunte, C.; Screpanti, E.; Venturi, M.; Rimon, A.; Padan, E.; Michel, H. Structure of a Na+/H+ antiporter and insights into mechanism of action and regulation by pH. Nature 2005, 435, 1197–1202. [Google Scholar] [CrossRef]
- Lee, C.; Kang, H.J.; von Ballmoos, C.; Newstead, S.; Uzdavinys, P.; Dotson, D.L.; Iwata, S.; Beckstein, O.; Cameron, A.D.; Drew, D. A two-domain elevator mechanism for sodium/proton antiport. Nature 2013, 501, 573–577. [Google Scholar] [CrossRef] [Green Version]
- Hisamitsu, T.; Ben Ammar, Y.; Nakamura, T.Y.; Wakabayashi, S. Dimerization is crucial for the function of the Na+/H+ exchanger NHE1. Biochemistry 2006, 45, 13346–13355. [Google Scholar] [CrossRef] [PubMed]
- Ariño, J.; Ramos, J.; Sychrová, H. Alkali metal cation transport and homeostasis in yeasts. Microbiol. Mol. Biol. Rev. 2010, 74, 95–120. [Google Scholar] [CrossRef] [Green Version]
- Hušeková, B.; Elicharová, H.; Sychrová, H. Pathogenic Candida species differ in the ability to grow at limiting potassium concentrations. Can. J. Microbiol. 2016, 62, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Durell, S.R.; Hao, Y.; Nakamura, T.; Bakker, E.P.; Guy, H.R. Evolutionary relationship between K(+) channels and symporters. Biophys. J. 1999, 77, 775–788. [Google Scholar] [CrossRef] [Green Version]
- Shamayeva, K.; Spurna, K.; Kulik, N.; Kale, D.; Munko, O.; Spurny, P.; Zayats, V.; Ludwig, J. MPM motifs of the yeast SKT protein Trk1 can assemble to form a functional K. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183513. [Google Scholar] [CrossRef] [PubMed]
- Doyle, D.A.; Morais Cabral, J.; Pfuetzner, R.A.; Kuo, A.; Gulbis, J.M.; Cohen, S.L.; Chait, B.T.; MacKinnon, R. The structure of the potassium channel: Molecular basis of K+ conduction and selectivity. Science 1998, 280, 69–77. [Google Scholar] [CrossRef] [Green Version]
- Zayats, V.; Stockner, T.; Pandey, S.K.; Wörz, K.; Ettrich, R.; Ludwig, J. A refined atomic scale model of the Saccharomyces cerevisiae K+-translocation protein Trk1p combined with experimental evidence confirms the role of selectivity filter glycines and other key residues. Biochim. Biophys. Acta 2015, 1848, 1183–1195. [Google Scholar] [CrossRef] [Green Version]
- Gaber, R.F.; Styles, C.A.; Fink, G.R. TRK1 encodes a plasma membrane protein required for high-affinity potassium transport in Saccharomyces cerevisiae. Mol. Cell. Biol. 1988, 8, 2848–2859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baev, D.; Rivetta, A.; Vylkova, S.; Sun, J.N.; Zeng, G.F.; Slayman, C.L.; Edgerton, M. The TRK1 potassium transporter is the critical effector for killing of Candida albicans by the cationic protein, Histatin 5. J. Biol. Chem. 2004, 279, 55060–55072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuroda, T.; Bihler, H.; Bashi, E.; Slayman, C.L.; Rivetta, A. Chloride channel function in the yeast TRK-potassium transporters. J. Membr. Biol. 2004, 198, 177–192. [Google Scholar] [CrossRef]
- Bañuelos, M.A.; Klein, R.D.; Alexander-Bowman, S.J.; Rodríguez-Navarro, A. A potassium transporter of the yeast Schwanniomyces occidentalis homologous to the Kup system of Escherichia coli has a high concentrative capacity. EMBO J. 1995, 14, 3021–3027. [Google Scholar] [CrossRef]
- Cabrera, E.; Álvarez, M.C.; Martín, Y.; Siverio, J.M.; Ramos, J. K(+) uptake systems in the yeast Hansenula polymorpha. Transcriptional and post-translational mechanisms involved in high-affinity K(+) transporter regulation. Fungal Genet. Biol. 2012, 49, 755–763. [Google Scholar] [CrossRef]
- Haro, R.; Sainz, L.; Rubio, F.; Rodríguez-Navarro, A. Cloning of two genes encoding potassium transporters in Neurospora crassa and expression of the corresponding cDNAs in Saccharomyces cerevisiae. Mol. Microbiol. 1999, 31, 511–520. [Google Scholar] [CrossRef]
- Bañuelos, M.A.; Madrid, R.; Rodríguez-Navarro, A. Individual functions of the HAK and TRK potassium transporters of Schwanniomyces occidentalis. Mol. Microbiol. 2000, 37, 671–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, J.L.; Sychrova, H.; Ramos, J. Monovalent cations regulate expression and activity of the Hak1 potassium transporter in Debaryomyces hansenii. Fungal Genet. Biol. 2011, 48, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Prista, C.; González-Hernández, J.C.; Ramos, J.; Loureiro-Dias, M.C. Cloning and characterization of two K+ transporters of Debaryomyces hansenii. Microbiology 2007, 153, 3034–3043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, Y.; Nanatani, K.; Hamamoto, S.; Shimizu, M.; Takahashi, M.; Tabuchi-Kobayashi, M.; Mizutani, A.; Schroeder, J.I.; Souma, S.; Uozumi, N. Defining membrane spanning domains and crucial membrane-localized acidic amino acid residues for K⁺ transport of a Kup/HAK/KT-type Escherichia coli potassium transporter. J. Biochem. 2014, 155, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Santa-María, G.E.; Oliferuk, S.; Moriconi, J.I. KT-HAK-KUP transporters in major terrestrial photosynthetic organisms: A twenty years tale. J. Plant. Physiol. 2018, 226, 77–90. [Google Scholar] [CrossRef]
- Tascón, I.; Sousa, J.S.; Corey, R.A.; Mills, D.J.; Griwatz, D.; Aumüller, N.; Mikusevic, V.; Stansfeld, P.J.; Vonck, J.; Hänelt, I. Structural basis of proton-coupled potassium transport in the KUP family. Nat. Commun. 2020, 11, 626. [Google Scholar] [CrossRef] [PubMed]
- Elicharová, H.; Hušeková, B.; Sychrová, H. Three Candida albicans potassium uptake systems differ in their ability to provide Saccharomyces cerevisiae trk1trk2 mutants with necessary potassium. FEMS Yeast Res. 2016, 16. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Castilla, F.J.; Bieber, J.; Caro, G.; Michán, C.; Sychrova, H.; Ramos, J. Regulation and activity of CaTrk1, CaAcu1 and CaHak1, the three plasma membrane potassium transporters in Candida albicans. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183486. [Google Scholar] [CrossRef] [PubMed]
- Cyert, M.S.; Philpott, C.C. Regulation of cation balance in Saccharomyces cerevisiae. Genetics 2013, 193, 677–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, J.; Kaandorp, J.A.; Sloot, P.M.; Lloyd, C.M.; Filatov, M.V. Calcium homeostasis and signaling in yeast cells and cardiac myocytes. FEMS Yeast Res. 2009, 9, 1137–1147. [Google Scholar] [CrossRef] [Green Version]
- Clapham, D.E. Calcium signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonilla, M.; Cunningham, K.W. Calcium release and influx in yeast: TRPC and VGCC rule another kingdom. Sci. STKE 2002, 2002, pe17. [Google Scholar] [CrossRef]
- Viladevall, L.; Serrano, R.; Ruiz, A.; Domenech, G.; Giraldo, J.; Barceló, A.; Ariño, J. Characterization of the calcium-mediated response to alkaline stress in Saccharomyces cerevisiae. J. Biol. Chem. 2004, 279, 43614–43624. [Google Scholar] [CrossRef] [Green Version]
- Batiza, A.F.; Schulz, T.; Masson, P.H. Yeast respond to hypotonic shock with a calcium pulse. J. Biol. Chem. 1996, 271, 23357–23362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, D.C.; Kim, H.; Mackin, N.A.; Maldonado-Báez, L.; Evangelista, C.C.; Beaudry, V.G.; Dudgeon, D.D.; Naiman, D.Q.; Erdman, S.E.; Cunningham, K.W. New regulators of a high affinity Ca2+ influx system revealed through a genome-wide screen in yeast. J. Biol. Chem. 2011, 286, 10744–10754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catterall, W.A. Structure and regulation of voltage-gated Ca2+ channels. Annu. Rev. Cell Dev. Biol. 2000, 16, 521–555. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.K.; Ellsmore, A.J.; Cessna, S.G.; Low, P.S.; Pardo, J.M.; Bressan, R.A.; Hasegawa, P.M. An osmotically induced cytosolic Ca2+ transient activates calcineurin signaling to mediate ion homeostasis and salt tolerance of Saccharomyces cerevisiae. J. Biol. Chem. 2002, 277, 33075–33080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Q.; Wang, H.; Cheng, X.; Xu, N.; Ding, X.; Xing, L.; Li, M. Roles of Cch1 and Mid1 in morphogenesis, oxidative stress response and virulence in Candida albicans. Mycopathologia 2012, 174, 359–369. [Google Scholar] [CrossRef]
- Liu, M.; Du, P.; Heinrich, G.; Cox, G.M.; Gelli, A. Cch1 mediates calcium entry in Cryptococcus neoformans and is essential in low-calcium environments. Eukaryot. Cell 2006, 5, 1788–1796. [Google Scholar] [CrossRef] [Green Version]
- Locke, E.G.; Bonilla, M.; Liang, L.; Takita, Y.; Cunningham, K.W. A homolog of voltage-gated Ca(2+) channels stimulated by depletion of secretory Ca(2+) in yeast. Mol. Cell. Biol. 2000, 20, 6686–6694. [Google Scholar] [CrossRef] [Green Version]
- Peiter, E.; Fischer, M.; Sidaway, K.; Roberts, S.K.; Sanders, D. The Saccharomyces cerevisiae Ca2+ channel Cch1pMid1p is essential for tolerance to cold stress and iron toxicity. FEBS Lett. 2005, 579, 5697–5703. [Google Scholar] [CrossRef]
- Van Itallie, C.M.; Anderson, J.M. Claudins and epithelial paracellular transport. Annu. Rev. Physiol. 2006, 68, 403–429. [Google Scholar] [CrossRef]
- Ding, X.; Yu, Q.; Xu, N.; Wang, Y.; Cheng, X.; Qian, K.; Zhao, Q.; Zhang, B.; Xing, L.; Li, M. Ecm7, a regulator of HACS, functions in calcium homeostasis maintenance, oxidative stress response and hyphal development in Candida albicans. Fungal Genet. Biol. 2013, 57, 23–32. [Google Scholar] [CrossRef]
- Popa, C.V.; Dumitru, I.; Ruta, L.L.; Danet, A.F.; Farcasanu, I.C. Exogenous oxidative stress induces Ca2+ release in the yeast Saccharomyces cerevisiae. FEBS J. 2010, 277, 4027–4038. [Google Scholar] [CrossRef]
- Kaur, R.; Castaño, I.; Cormack, B.P. Functional genomic analysis of fluconazole susceptibility in the pathogenic yeast Candida glabrata: Roles of calcium signaling and mitochondria. Antimicrob. Agents Chemother. 2004, 48, 1600–1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, M.P.; Vu, K.; Bautos, J.; Gelli, A. Cch1 restores intracellular Ca2+ in fungal cells during endoplasmic reticulum stress. J. Biol. Chem. 2010, 285, 10951–10958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonilla, M.; Cunningham, K.W. Mitogen-activated protein kinase stimulation of Ca(2+) signaling is required for survival of endoplasmic reticulum stress in yeast. Mol. Biol. Cell 2003, 14, 4296–4305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonilla, M.; Nastase, K.K.; Cunningham, K.W. Essential role of calcineurin in response to endoplasmic reticulum stress. EMBO J. 2002, 21, 2343–2353. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, K.W.; Fink, G.R. Ca2+ transport in Saccharomyces cerevisiae. J. Exp. Biol. 1994, 196, 157–166. [Google Scholar]
- Wang, Y.; Wang, J.; Cheng, J.; Xu, D.; Jiang, L. Genetic interactions between the Golgi Ca2+/H+ exchanger Gdt1 and the plasma membrane calcium channel Cch1/Mid1 in the regulation of calcium homeostasis, stress response and virulence in Candida albicans. FEMS Yeast Res. 2015, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar, P.S.; Engel, A.; Walter, P. The plasma membrane proteins Prm1 and Fig1 ascertain fidelity of membrane fusion during yeast mating. Mol. Biol. Cell 2007, 18, 547–556. [Google Scholar] [CrossRef] [Green Version]
- Muller, E.M.; Mackin, N.A.; Erdman, S.E.; Cunningham, K.W. Fig1p facilitates Ca2+ influx and cell fusion during mating of Saccharomyces cerevisiae. J. Biol. Chem. 2003, 278, 38461–38469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Brand, A.; Srikantha, T.; Daniels, K.J.; Soll, D.R.; Gow, N.A. Fig1 facilitates calcium influx and localizes to membranes destined to undergo fusion during mating in Candida albicans. Eukaryot. Cell 2011, 10, 435–444. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Alber, J.; Wang, J.; Du, W.; Yang, X.; Li, X.; Sanglard, D.; Geyer, J. The Candida albicans plasma membrane protein Rch1p, a member of the vertebrate SLC10 carrier family, is a novel regulator of cytosolic Ca2+ homoeostasis. Biochem. J. 2012, 444, 497–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alber, J.; Jiang, L.; Geyer, J. CaRch1p does not functionally interact with the high-affinity Ca(2+) influx system (HACS) of Candida albicans. Yeast 2013, 30, 449–457. [Google Scholar] [CrossRef]
- Erdman, S.; Lin, L.; Malczynski, M.; Snyder, M. Pheromone-regulated genes required for yeast mating differentiation. J. Cell Biol. 1998, 140, 461–483. [Google Scholar] [CrossRef] [Green Version]
- Solis, N.V.; Park, Y.N.; Swidergall, M.; Daniels, K.J.; Filler, S.G.; Soll, D.R. Candida albicans White-Opaque Switching Influences Virulence but Not Mating during Oropharyngeal Candidiasis. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef] [Green Version]
- Daniels, K.J.; Srikantha, T.; Lockhart, S.R.; Pujol, C.; Soll, D.R. Opaque cells signal white cells to form biofilms in Candida albicans. EMBO J. 2006, 25, 2240–2252. [Google Scholar] [CrossRef]
- Brand, A.; Lee, K.; Veses, V.; Gow, N.A. Calcium homeostasis is required for contact-dependent helical and sinusoidal tip growth in Candida albicans hyphae. Mol. Microbiol. 2009, 71, 1155–1164. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.N.; Dudgeon, D.D.; Paliwal, S.; Levchenko, A.; Grote, E.; Cunningham, K.W. Multiple signaling pathways regulate yeast cell death during the response to mating pheromones. Mol. Biol. Cell 2006, 17, 3409–3422. [Google Scholar] [CrossRef]
- Arellano, V.J.; Martinell García, P.; Rodríguez Plaza, J.G.; Lara Ortiz, M.T.; Schreiber, G.; Volkmer, R.; Klipp, E.; Rio, G.D. An Antimicrobial Peptide Induces. Front. Microbiol. 2018, 9, 1240. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Cheng, J.; Cao, C.; Wang, L.; Jiang, L. Genetic interactions between Rch1 and the high-affinity calcium influx system Cch1/Mid1/Ecm7 in the regulation of calcium homeostasis, drug tolerance, hyphal development and virulence in Candida albicans. FEMS Yeast Res. 2015, 15. [Google Scholar] [CrossRef] [Green Version]
- Sanglard, D.; Ischer, F.; Marchetti, O.; Entenza, J.; Bille, J. Calcineurin A of Candida albicans: Involvement in antifungal tolerance, cell morphogenesis and virulence. Mol. Microbiol. 2003, 48, 959–976. [Google Scholar] [CrossRef] [PubMed]
- Bairwa, G.; Hee Jung, W.; Kronstad, J.W. Iron acquisition in fungal pathogens of humans. Metallomics 2017, 9, 215–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, D.H. Acquisition, transport, and storage of iron by pathogenic fungi. Clin. Microbiol. Rev. 1999, 12, 394–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philpott, C.C. Iron uptake in fungi: A system for every source. Biochim. Biophys. Acta 2006, 1763, 636–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutak, R.; Lesuisse, E.; Tachezy, J.; Richardson, D.R. Crusade for iron: Iron uptake in unicellular eukaryotes and its significance for virulence. Trends Microbiol. 2008, 16, 261–268. [Google Scholar] [CrossRef]
- Soares, M.P.; Weiss, G. The Iron age of host-microbe interactions. EMBO Rep. 2015, 16, 1482–1500. [Google Scholar] [CrossRef] [Green Version]
- Devaux, F.; Thiébaut, A. The regulation of iron homeostasis in the fungal human pathogen. Microbiology 2019, 165, 1041–1060. [Google Scholar] [CrossRef]
- Collins, H.L. Withholding iron as a cellular defence mechanism—Friend or foe? Eur. J. Immunol. 2008, 38, 1803–1806. [Google Scholar] [CrossRef]
- Crichton, R.R.; Ward, R.J. Iron homeostasis. Met. Ions Biol. Syst. 1998, 35, 633–665. [Google Scholar]
- Valenti, P.; Visca, P.; Antonini, G.; Orsi, N. Interaction between lactoferrin and ovotransferrin and Candida cells. FEMS Microbiol. Lett. 1986, 33, 271–275. [Google Scholar] [CrossRef]
- Chaffin, W.L.; López-Ribot, J.L.; Casanova, M.; Gozalbo, D.; Martínez, J.P. Cell wall and secreted proteins of Candida albicans: Identification, function, and expression. Microbiol. Mol. Biol. Rev. 1998, 62, 130–180. [Google Scholar]
- Fratti, R.A.; Belanger, P.H.; Ghannoum, M.A.; Edwards, J.E.; Filler, S.G. Endothelial cell injury caused by Candida albicans is dependent on iron. Infect. Immun. 1998, 66, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Gerwien, F.; Skrahina, V.; Kasper, L.; Hube, B.; Brunke, S. Metals in fungal virulence. FEMS Microbiol. Rev. 2018, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drakesmith, H.; Prentice, A.M. Hepcidin and the iron-infection axis. Science 2012, 338, 768–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganz, T.; Nemeth, E. Iron homeostasis in host defence and inflammation. Nat. Rev. Immunol. 2015, 15, 500–510. [Google Scholar] [CrossRef] [Green Version]
- Malavia, D.; Crawford, A.; Wilson, D. Nutritional Immunity and Fungal Pathogenesis: The Struggle for Micronutrients at the Host-Pathogen Interface. Adv. Microb. Physiol. 2017, 70, 85–103. [Google Scholar] [CrossRef]
- Li, Y.; Sun, L.; Lu, C.; Gong, Y.; Li, M.; Sun, S. Promising Antifungal Targets Against. Front. Cell. Infect. Microbiol. 2018, 8, 286. [Google Scholar] [CrossRef] [PubMed]
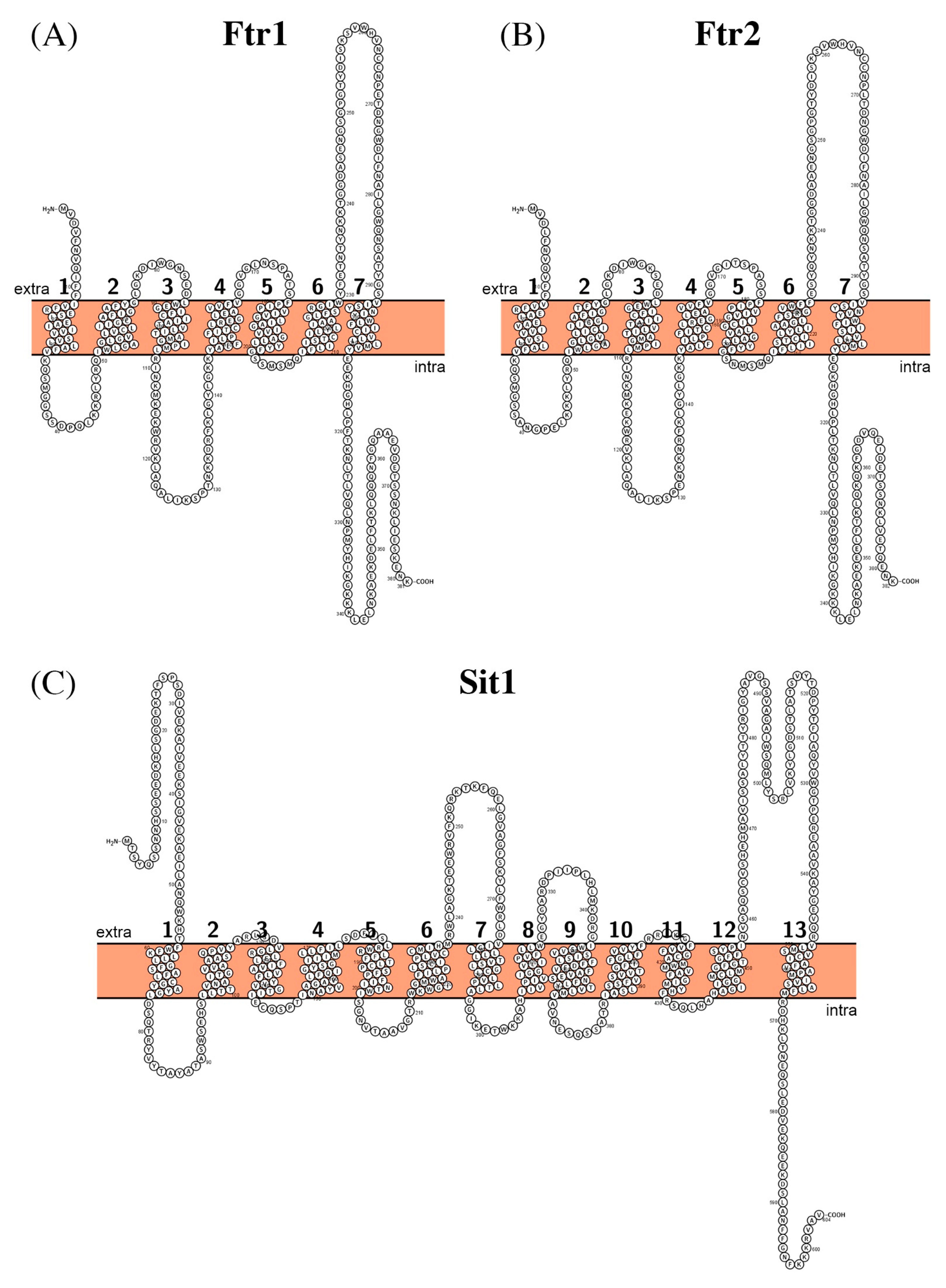
- Ramanan, N.; Wang, Y. A high-affinity iron permease essential for Candida albicans virulence. Science 2000, 288, 1062–1064. [Google Scholar] [CrossRef]
- Fang, H.M.; Wang, Y. Characterization of iron-binding motifs in Candida albicans high-affinity iron permease CaFtr1p by site-directed mutagenesis. Biochem. J. 2002, 368, 641–647. [Google Scholar] [CrossRef] [Green Version]
- Severance, S.; Chakraborty, S.; Kosman, D.J. The Ftr1p iron permease in the yeast plasma membrane: Orientation, topology and structure-function relationships. Biochem. J. 2004, 380, 487–496. [Google Scholar] [CrossRef] [Green Version]
- Askwith, C.; Eide, D.; Van Ho, A.; Bernard, P.S.; Li, L.; Davis-Kaplan, S.; Sipe, D.M.; Kaplan, J. The FET3 gene of S. cerevisiae encodes a multicopper oxidase required for ferrous iron uptake. Cell 1994, 76, 403–410. [Google Scholar] [CrossRef]
- Dancis, A.; Yuan, D.S.; Haile, D.; Askwith, C.; Eide, D.; Moehle, C.; Kaplan, J.; Klausner, R.D. Molecular characterization of a copper transport protein in S. cerevisiae: An unexpected role for copper in iron transport. Cell 1994, 76, 393–402. [Google Scholar] [CrossRef]
- Spizzo, T.; Byersdorfer, C.; Duesterhoeft, S.; Eide, D. The yeast FET5 gene encodes a FET3-related multicopper oxidase implicated in iron transport. Mol. Gen. Genet. 1997, 256, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Severance, S.; Kaur, N.; Wiltsie, W.; Kosman, D.J. Assembly, activation, and trafficking of the Fet3p.Ftr1p high affinity iron permease complex in Saccharomyces cerevisiae. J. Biol. Chem. 2006, 281, 13355–13364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mamouei, Z.; Zeng, G.; Wang, Y.M.; Wang, Y. Candida albicans possess a highly versatile and dynamic high-affinity iron transport system important for its commensal-pathogenic lifestyle. Mol. Microbiol. 2017, 106, 986–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, C.J.; Bai, C.; Zheng, X.D.; Wang, Y.M.; Wang, Y. Characterization and functional analysis of the siderophore-iron transporter CaArn1p in Candida albicans. J. Biol. Chem. 2002, 277, 30598–30605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesuisse, E.; Simon-Casteras, M.; Labbe, P. Siderophore-mediated iron uptake in Saccharomyces cerevisiae: The SIT1 gene encodes a ferrioxamine B permease that belongs to the major facilitator superfamily. Microbiology 1998, 144, 3455–3462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, C.W.; Tiedeman, J.S.; Moore, R.E.; Philpott, C.C. Siderophore-iron uptake in saccharomyces cerevisiae. Identification of ferrichrome and fusarinine transporters. J. Biol. Chem. 2000, 275, 16354–16359. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.S.; Kim, J.Y.; Yun, C.W. Identification of ferrichrome- and ferrioxamine B-mediated iron uptake by Aspergillus fumigatus. Biochem. J. 2016, 473, 1203–1213. [Google Scholar] [CrossRef] [Green Version]
- Tangen, K.L.; Jung, W.H.; Sham, A.P.; Lian, T.; Kronstad, J.W. The iron- and cAMP-regulated gene SIT1 influences ferrioxamine B utilization, melanization and cell wall structure in Cryptococcus neoformans. Microbiology 2007, 153, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Dias, P.J.; Sá-Correia, I. The drug:H⁺ antiporters of family 2 (DHA2), siderophore transporters (ARN) and glutathione:H⁺ antiporters (GEX) have a common evolutionary origin in hemiascomycete yeasts. BMC Genom. 2013, 14, 901. [Google Scholar] [CrossRef] [Green Version]
- Ardon, O.; Bussey, H.; Philpott, C.; Ward, D.M.; Davis-Kaplan, S.; Verroneau, S.; Jiang, B.; Kaplan, J. Identification of a Candida albicans ferrichrome transporter and its characterization by expression in Saccharomyces cerevisiae. J. Biol. Chem. 2001, 276, 43049–43055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heymann, P.; Gerads, M.; Schaller, M.; Dromer, F.; Winkelmann, G.; Ernst, J.F. The siderophore iron transporter of Candida albicans (Sit1p/Arn1p) mediates uptake of ferrichrome-type siderophores and is required for epithelial invasion. Infect. Immun. 2002, 70, 5246–5255. [Google Scholar] [CrossRef] [Green Version]
- Yun, C.W.; Bauler, M.; Moore, R.E.; Klebba, P.E.; Philpott, C.C. The role of the FRE family of plasma membrane reductases in the uptake of siderophore-iron in Saccharomyces cerevisiae. J. Biol. Chem. 2001, 276, 10218–10223. [Google Scholar] [CrossRef] [Green Version]
- Manns, J.M.; Mosser, D.M.; Buckley, H.R. Production of a hemolytic factor by Candida albicans. Infect. Immun. 1994, 62, 5154–5156. [Google Scholar] [CrossRef] [Green Version]
- Moors, M.A.; Stull, T.L.; Blank, K.J.; Buckley, H.R.; Mosser, D.M. A role for complement receptor-like molecules in iron acquisition by Candida albicans. J. Exp. Med. 1992, 175, 1643–1651. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Samaranayake, L.P.; Yau, J.Y. Candida species exhibit differential in vitro hemolytic activities. J. Clin. Microbiol. 2001, 39, 2971–2974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pendrak, M.L.; Yan, S.S.; Roberts, D.D. Sensing the host environment: Recognition of hemoglobin by the pathogenic yeast Candida albicans. Arch. Biochem. Biophys. 2004, 426, 148–156. [Google Scholar] [CrossRef]
- Rüchel, R.; Tegeler, R.; Trost, M. A comparison of secretory proteinases from different strains of Candida albicans. Sabouraudia 1982, 20, 233–244. [Google Scholar] [CrossRef]
- Knight, S.A.; Vilaire, G.; Lesuisse, E.; Dancis, A. Iron acquisition from transferrin by Candida albicans depends on the reductive pathway. Infect. Immun. 2005, 73, 5482–5492. [Google Scholar] [CrossRef] [Green Version]
- Andreini, C.; Bertini, I.; Rosato, A. Metalloproteomes: A bioinformatic approach. Acc. Chem. Res. 2009, 42, 1471–1479. [Google Scholar] [CrossRef]
- Hood, M.I.; Skaar, E.P. Nutritional immunity: Transition metals at the pathogen-host interface. Nat. Rev. Microbiol. 2012, 10, 525–537. [Google Scholar] [CrossRef]
- Becker, K.W.; Skaar, E.P. Metal limitation and toxicity at the interface between host and pathogen. FEMS Microbiol. Rev. 2014, 38, 1235–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gati, C.; Stetsenko, A.; Slotboom, D.J.; Scheres, S.H.W.; Guskov, A. The structural basis of proton driven zinc transport by ZntB. Nat. Commun. 2017, 8, 1313. [Google Scholar] [CrossRef]
- Yike, I. Fungal proteases and their pathophysiological effects. Mycopathologia 2011, 171, 299–323. [Google Scholar] [CrossRef] [PubMed]
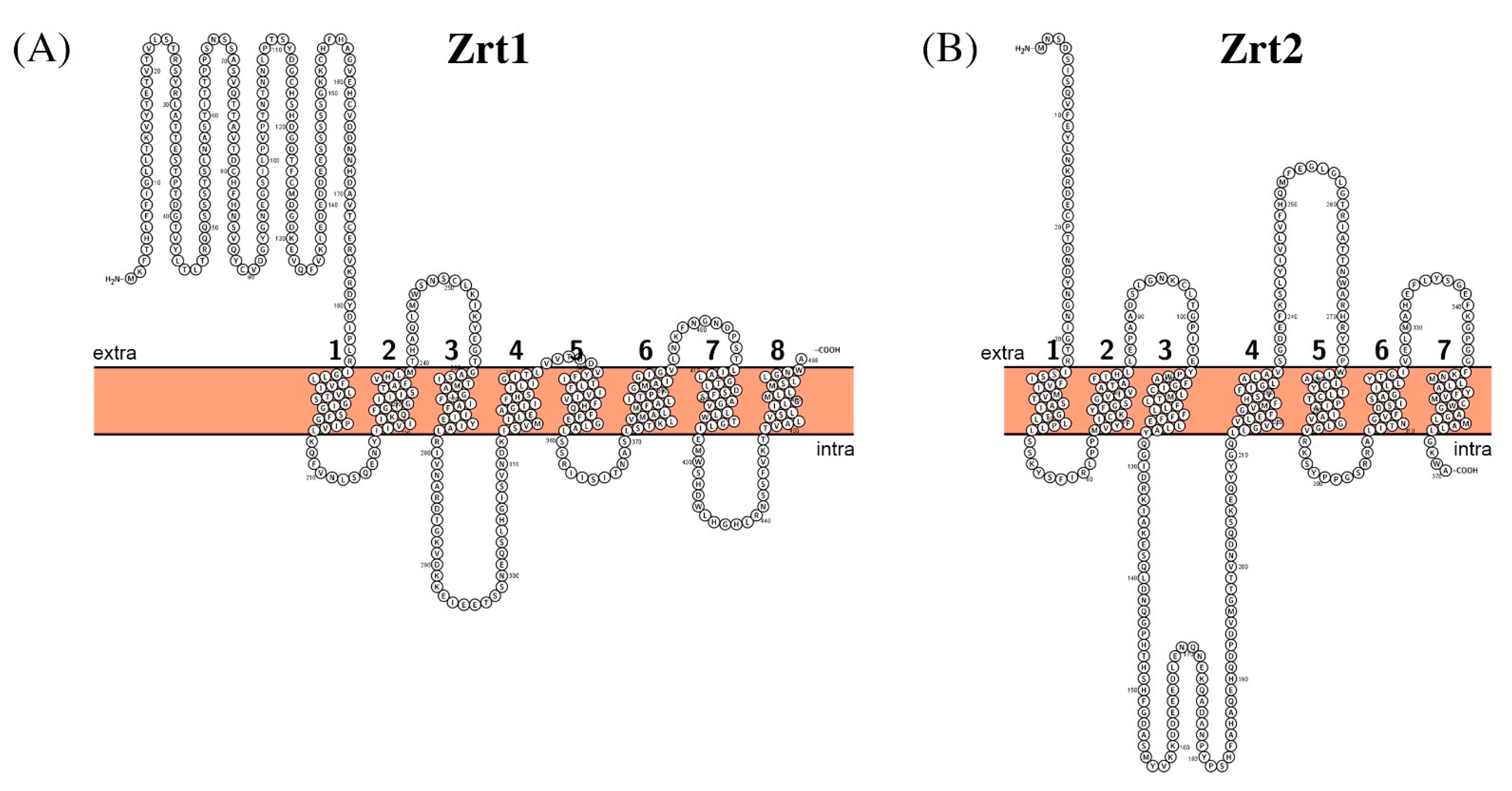
- Korshunova, Y.O.; Eide, D.; Clark, W.G.; Guerinot, M.L.; Pakrasi, H.B. The IRT1 protein from Arabidopsis thaliana is a metal transporter with a broad substrate range. Plant. Mol. Biol. 1999, 40, 37–44. [Google Scholar] [CrossRef]
- Schaaf, G.; Honsbein, A.; Meda, A.R.; Kirchner, S.; Wipf, D.; von Wirén, N. AtIREG2 encodes a tonoplast transport protein involved in iron-dependent nickel detoxification in Arabidopsis thaliana roots. J. Biol. Chem. 2006, 281, 25532–25540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huynh, C.; Sacks, D.L.; Andrews, N.W. A Leishmania amazonensis ZIP family iron transporter is essential for parasite replication within macrophage phagolysosomes. J. Exp. Med. 2006, 203, 2363–2375. [Google Scholar] [CrossRef] [PubMed]
- Crawford, A.C.; Lehtovirta-Morley, L.E.; Alamir, O.; Niemiec, M.J.; Alawfi, B.; Alsarraf, M.; Skrahina, V.; Costa, A.C.B.P.; Anderson, A.; Yellagunda, S.; et al. Biphasic zinc compartmentalisation in a human fungal pathogen. PLoS Pathog. 2018, 14, e1007013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sentandreu, M.; Elorza, M.V.; Sentandreu, R.; Fonzi, W.A. Cloning and characterization of PRA1, a gene encoding a novel pH-regulated antigen of Candida albicans. J. Bacteriol. 1998, 180, 282–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Łoboda, D.; Rowińska-Żyrek, M. Zinc binding sites in Pra1, a zincophore from Candida albicans. Dalton Trans. 2017, 46, 13695–13703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Citiulo, F.; Jacobsen, I.D.; Miramón, P.; Schild, L.; Brunke, S.; Zipfel, P.; Brock, M.; Hube, B.; Wilson, D. Candida albicans scavenges host zinc via Pra1 during endothelial invasion. PLoS Pathog. 2012, 8, e1002777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amich, J.; Vicentefranqueira, R.; Mellado, E.; Ruiz-Carmuega, A.; Leal, F.; Calera, J.A. The ZrfC alkaline zinc transporter is required for Aspergillus fumigatus virulence and its growth in the presence of the Zn/Mn-chelating protein calprotectin. Cell. Microbiol. 2014, 16, 548–564. [Google Scholar] [CrossRef]
- Dade, J.; DuBois, J.C.; Pasula, R.; Donnell, A.M.; Caruso, J.A.; Smulian, A.G.; Deepe, G.S. HcZrt2, a zinc responsive gene, is indispensable for the survival of Histoplasma capsulatum in vivo. Med. Mycol. 2016, 54, 865–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Do, E.; Hu, G.; Caza, M.; Kronstad, J.W.; Jung, W.H. The ZIP family zinc transporters support the virulence of Cryptococcus neoformans. Med. Mycol. 2016, 54, 605–615. [Google Scholar] [CrossRef] [Green Version]
- Schneider, R.O.; Diehl, C.; Dos Santos, F.M.; Piffer, A.C.; Garcia, A.W.A.; Kulmann, M.I.R.; Schrank, A.; Kmetzsch, L.; Vainstein, M.H.; Staats, C.C. Effects of zinc transporters on Cryptococcus gattii virulence. Sci. Rep. 2015, 5, 10104. [Google Scholar] [CrossRef] [Green Version]
- Festa, R.A.; Thiele, D.J. Copper: An essential metal in biology. Curr. Biol. 2011, 21, R877–R883. [Google Scholar] [CrossRef] [Green Version]
- Gunther, M.R.; Hanna, P.M.; Mason, R.P.; Cohen, M.S. Hydroxyl radical formation from cuprous ion and hydrogen peroxide: A spin-trapping study. Arch. Biochem. Biophys. 1995, 316, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Niewerth, M.; Kunze, D.; Seibold, M.; Schaller, M.; Korting, H.C.; Hube, B. Ciclopirox olamine treatment affects the expression pattern of Candida albicans genes encoding virulence factors, iron metabolism proteins, and drug resistance factors. Antimicrob. Agents Chemother. 2003, 47, 1805–1817. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Zhang, L.X.; Zhang, J.D.; Cao, Y.B.; Yu, Y.Y.; Wang, D.J.; Ying, K.; Chen, W.S.; Jiang, Y.Y. cDNA microarray analysis of differential gene expression and regulation in clinically drug-resistant isolates of Candida albicans from bone marrow transplanted patients. Int. J. Med. Microbiol. 2006, 296, 421–434. [Google Scholar] [CrossRef]
- Mackie, J.; Szabo, E.K.; Urgast, D.S.; Ballou, E.R.; Childers, D.S.; MacCallum, D.M.; Feldmann, J.; Brown, A.J. Host-Imposed Copper Poisoning Impacts Fungal Micronutrient Acquisition during Systemic Candida albicans Infections. PLoS ONE 2016, 11, e0158683. [Google Scholar] [CrossRef] [Green Version]
- Djoko, K.Y.; Ong, C.L.; Walker, M.J.; McEwan, A.G. The Role of Copper and Zinc Toxicity in Innate Immune Defense against Bacterial Pathogens. J. Biol. Chem. 2015, 290, 18954–18961. [Google Scholar] [CrossRef] [Green Version]
- Marvin, M.E.; Mason, R.P.; Cashmore, A.M. The CaCTR1 gene is required for high-affinity iron uptake and is transcriptionally controlled by a copper-sensing transactivator encoded by CaMAC1. Microbiology 2004, 150, 2197–2208. [Google Scholar] [CrossRef]
- Woodacre, A.; Mason, R.P.; Jeeves, R.E.; Cashmore, A.M. Copper-dependent transcriptional regulation by Candida albicans Mac1p. Microbiology 2008, 154, 1502–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bensen, E.S.; Martin, S.J.; Li, M.; Berman, J.; Davis, D.A. Transcriptional profiling in Candida albicans reveals new adaptive responses to extracellular pH and functions for Rim101p. Mol. Microbiol. 2004, 54, 1335–1351. [Google Scholar] [CrossRef]
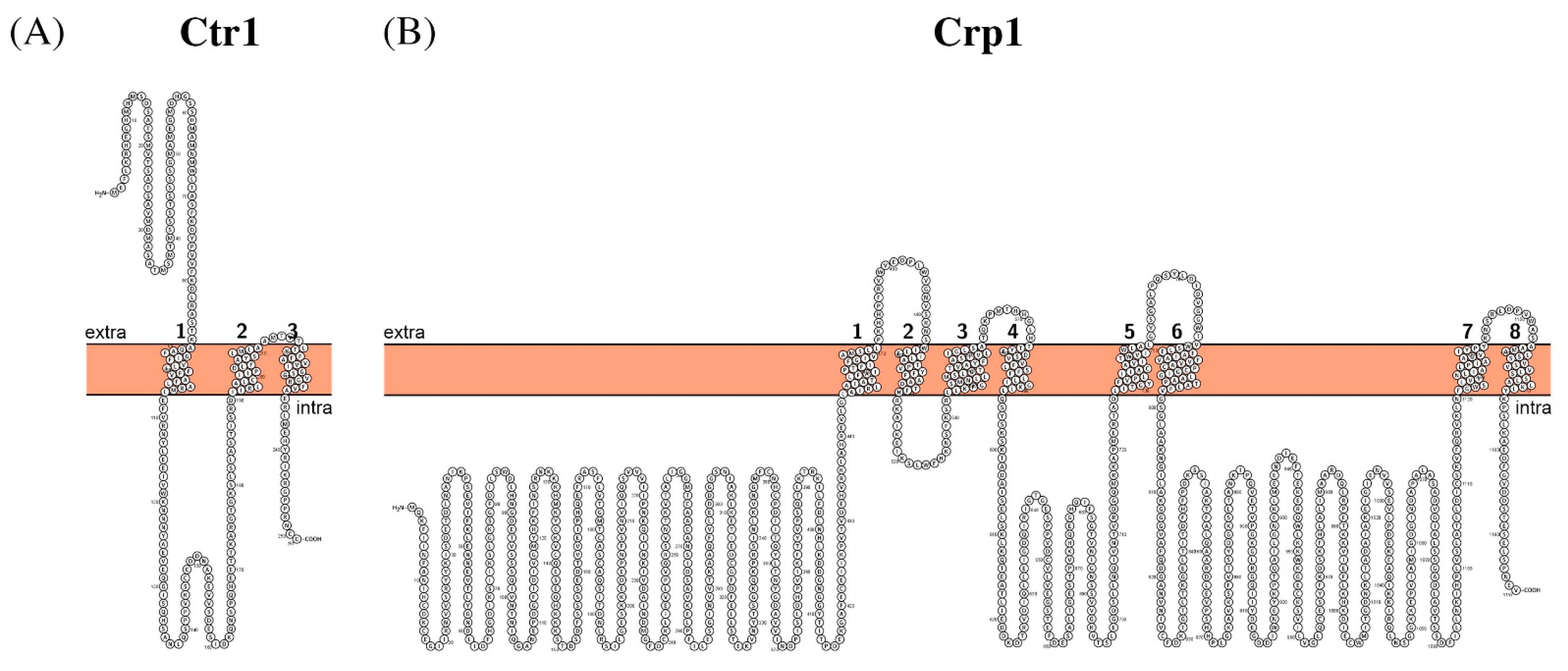
- Aller, S.G.; Unger, V.M. Projection structure of the human copper transporter CTR1 at 6-A resolution reveals a compact trimer with a novel channel-like architecture. Proc. Natl. Acad. Sci. USA 2006, 103, 3627–3632. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Peña, M.M.; Nose, Y.; Thiele, D.J. Biochemical characterization of the human copper transporter Ctr1. J. Biol. Chem. 2002, 277, 4380–4387. [Google Scholar] [CrossRef] [Green Version]
- Dong, Z.; Wang, Y.; Wang, C.; Xu, H.; Guan, L.; Li, Z.; Li, F. Self-Assembly of the Second Transmembrane Domain of hCtr1 in Micelles and Interaction with Silver Ion. J. Phys. Chem. B 2015, 119, 8302–8312. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Wedd, A.G. A C-terminal domain of the membrane copper pump Ctr1 exchanges copper(I) with the copper chaperone Atx1. Chem. Commun. 2002, 588–589. [Google Scholar] [CrossRef] [PubMed]
- Eisses, J.F.; Kaplan, J.H. The mechanism of copper uptake mediated by human CTR1: A mutational analysis. J. Biol. Chem. 2005, 280, 37159–37168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khemiri, I.; Tebbji, F.; Sellam, A. Transcriptome Analysis Uncovers a Link Between Copper Metabolism, and Both Fungal Fitness and Antifungal Sensitivity in the Opportunistic Yeast. Front. Microbiol. 2020, 11, 935. [Google Scholar] [CrossRef]
- Liu, X.F.; Supek, F.; Nelson, N.; Culotta, V.C. Negative control of heavy metal uptake by the Saccharomyces cerevisiae BSD2 gene. J. Biol. Chem. 1997, 272, 11763–11769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, W.J.; Loguinov, A.; Chang, M.; Wintz, H.; Nislow, C.; Arkin, A.P.; Giaever, G.; Vulpe, C.D. Identification of genes involved in the toxic response of Saccharomyces cerevisiae against iron and copper overload by parallel analysis of deletion mutants. Toxicol. Sci. 2008, 101, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Weissman, Z.; Berdicevsky, I.; Cavari, B.Z.; Kornitzer, D. The high copper tolerance of Candida albicans is mediated by a P-type ATPase. Proc. Natl. Acad. Sci. USA 2000, 97, 3520–3525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gourdon, P.; Liu, X.Y.; Skjørringe, T.; Morth, J.P.; Møller, L.B.; Pedersen, B.P.; Nissen, P. Crystal structure of a copper-transporting PIB-type ATPase. Nature 2011, 475, 59–64. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Uniprot Accession Number | Candida Genome Database Accession Number | Transporter Classification Database Identifier |
|---|---|---|---|
| Mep1 | A0A1D8PJF2 | C3_02310W | 1.A.11.3 |
| Mep2 | Q59UP8 | C4_00430W | 1.A.11.3 |
| Cnh1 | Q9P937 | C4_00040W | 2.A.36.4.4 |
| Trk1 | A0A1D8PTL7 | CR_07960C | 2.A.38.2 |
| Hak1 | A0A1D8PDU7 | C1_06610C | 2.A.72.2 |
| Acu1 | Q5A9B2 | CR_01640C | 3.A.3 |
| Cch1 | Q5A936 | C1_01100W | 1.A.1.11 |
| Mid1 | A0A1D8PNU4 | C5_03990W | 8.A.41.1 |
| Ecm7 | Q59US7 | C4_00180W | 1.H.1.4 |
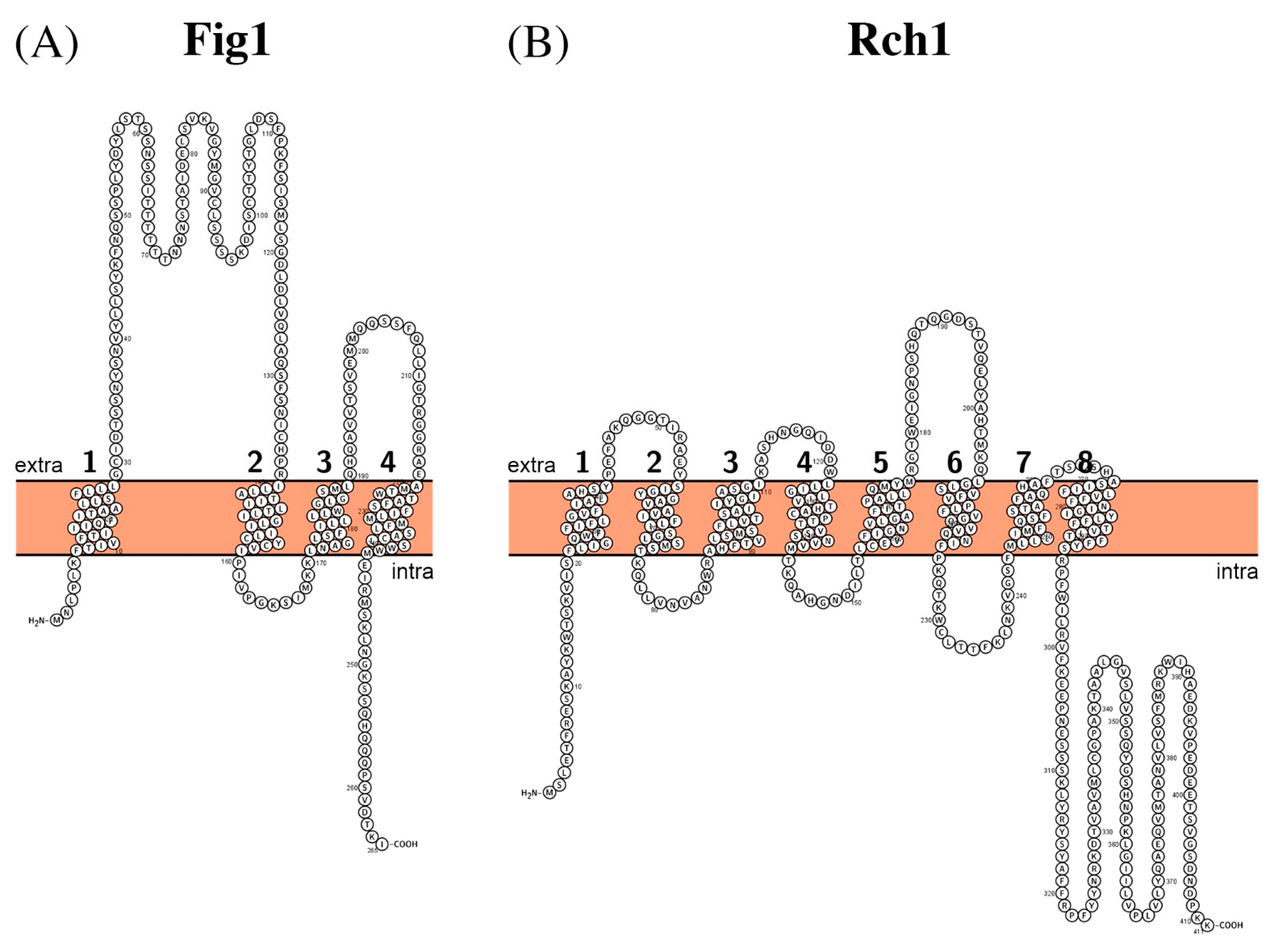
| Fig1 | Q59WR6 | C6_01310W | 1.A.81.2.1 |
| Rch1 | Q59UQ7 | C4_00360C | 2.A.28.3.7 |
| Ftr1 | A0A1D8PFV0 | C1_14130W | 2.A.108.1.2 |
| Ftr2 | A0A1D8PFV2 | C1_14220C | 2.A.108.1.3 |
| Sit1 | Q5A2T6 | C2_08050C | 2.A.1.16 |
| Zrt1 | A0A1D8PMR6 | C4_06970C | 2.A.5.1 |
| Zrt2 | A0A1D8PGN5 | C2_02590W | 2.A.5.1 |
| Ctr1 | Q59NP1 | C6_00790C | 1.A.56.2.2 |
| Crp1 | A0A1D8PEI4 | C1_09250W | 3.A.3.5.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Volkova, M.; Atamas, A.; Tsarenko, A.; Rogachev, A.; Guskov, A. Cation Transporters of Candida albicans—New Targets to Fight Candidiasis? Biomolecules 2021, 11, 584. https://doi.org/10.3390/biom11040584
Volkova M, Atamas A, Tsarenko A, Rogachev A, Guskov A. Cation Transporters of Candida albicans—New Targets to Fight Candidiasis? Biomolecules. 2021; 11(4):584. https://doi.org/10.3390/biom11040584
Chicago/Turabian StyleVolkova, Marina, Anastasia Atamas, Alexey Tsarenko, Andrey Rogachev, and Albert Guskov. 2021. "Cation Transporters of Candida albicans—New Targets to Fight Candidiasis?" Biomolecules 11, no. 4: 584. https://doi.org/10.3390/biom11040584