1. Introduction
Bacteria live in enormous numbers in almost every environment on earth, from deep-sea vents to the digestive tracts of humans. As important decomposers in the earth’s ecosystem, bacteria are responsible for breaking down dead plants and animals into organic compounds. However, bacterial pathogens pose one of the most urgent global health threats because of their growing resistance to current antibacterial drugs [
1]. Compared with Gram-positive bacteria, the Gram-negative bacteria are harder to kill because of the presence of the outer membrane, which contributes to the prevention of the penetration of antibacterial drugs into the cells [
2]. Infections caused by drug-resistant bacteria have aroused wide concern because of the lack of effective antimicrobial agents [
3]. Compared with conventional antibiotics, antimicrobial proteins (AMPs) directed to the bacterial membrane are assumed to be more effective at targeting bacteria and preventing resistance [
4].
AMPs are evolutionarily conserved components of the innate immune response that provide host defense at the skin and mucosal surface [
5]. The human AMPs identified to date include mainly histatins, hepcidins, defensins, and LL-37. Histatins, which are found in human saliva, possess antifungal properties by binding to a receptor on the fungal cell membrane [
6]. Hepcidins are produced by the human liver and serve as potential antibacterial and antifungal agents [
7]. Human defensins, including α-defensins and β-defensins, contribute to the antimicrobial action of granulocytes, the mucosal host defense in the small intestine, as well as the epithelial host defense in the skin and elsewhere [
8]. LL-37 is a 37-residue, amphipathic, helical peptide that is found throughout the body and exhibits a broad spectrum of antimicrobial activities [
9]. Moreover, increasing evidence has demonstrated that AMPs have many other biological and physiological functions. For example, LL-37 not only exerts a bactericidal effect but also plays an important role in immune modulation and wound healing [
10].
Human genome projects have produced a large amount of genomic sequence data. However, the biological functions of many proteins encoded by the sequences obtained remain unknown. Functional research on these proteins may help us uncover novel molecular pathways and potential drug targets. Previously, we identified an uncharacterized peptide, chromosome 10 open reading frame 99 (
C10orf99), as a type of human AMP, which was termed AP-57 [
11]. Here, using a bioinformatics analysis, we found that another gene, chromosome 5 open reading frame 46 (
C5orf46), encodes an amphiphilic small secretory peptide. Through prokaryotic expression and purification, as well as an antibacterial assay,
C5orf46 was identified as a type of AMP.
2. Materials and Methods
2.1. Cell Culture
The cell lines used in this study were purchased from the American Type Culture Collection (ATCC). The cells were cultured in DMEM or RPMI 1640 medium containing 10% fetal bovine serum (Gibco, Grand Island, NY, USA) and 100 μg/mL penicillin–streptomycin (ZS808; Zomanbio, Beijing, China) at 37 °C in a humidified atmosphere of 5% CO2.
2.2. Expression and Purification of Recombinant Proteins
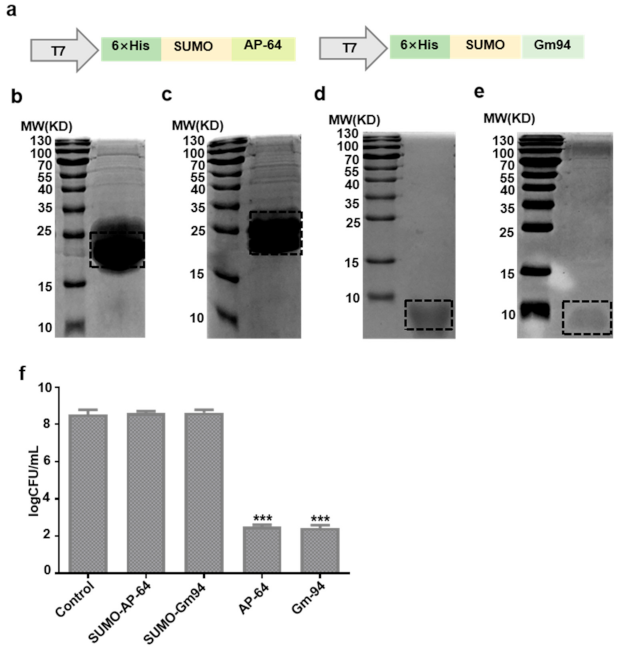
The sequences of human
C5orf46 (AP-64) and mouse
C5orf46 (Gm94 named by Harney et al.) [
12] were obtained from the National Center for Biotechnology Information database and synthesized by GENEWIZ (Beijing, China). A small ubiquitin-related modifier (SUMO) tag was fused to the N terminus of AP-64 or Gm94. The fragments were subcloned into the pET28a vector (69864-3; Addgene, Watertown, MA, USA) to construct the recombinant plasmids pET28a-SUMO-AP-64 and pET28a-SUMO-Gm94. Recombinant protein expression and purification were performed generally according to a previously reported method [
13]. Briefly, the recombinant plasmid was transformed into
E. coli Rosetta (DE3) cells. The cells were cultured in lysogeny broth (LB) medium at 37 °C to an optical density at 600 nm (OD600) of 0.6–0.8. Isopropyl β-
d-1-thiogalactopyranoside (IPTG, 1 mM) was added to the culture to induce recombinant protein expression. The cells were harvested and lysed by sonication 16 h after induction. The lysates were fractionated by centrifugation at 15,000 rpm for 15 min at 4 °C. The supernatant was applied to a Ni-nitrilotriacetate (Ni-NTA) Sepharose column, and 6× His-tagged SUMO-AP-64 or SUMO-Gm94 was eluted using elution buffer (500 mM NaCl and 250 mM imidazole, pH 8.0). The fusion proteins diluted in phosphate-buffered saline (PBS) at a final concentration of 1 mg/mL were cleaved by SUMO protease (12588018; Invitrogen, Shanghai, China) at 4 °C for 6 h. The cleaved samples were reloaded onto the Ni-NTA resin to obtain the AP-64 or Gm94 protein. The protein concentration was measured with a BCA (P0012S; Beyotime, Shanghai, China) assay kit according to the manufacturer’s instructions.
2.3. Antibacterial Activity Detection of Purification Proteins
E. coli DH5α cells were purchased from Invitrogen (18288019; Shanghai, China) for antibacterial activity detection of purified proteins. After overnight growth in LB at 37 °C, E. coli DH5α cells were washed and diluted in LB to an OD600 of 0.1. After adding SUMO-AP-64 (21 μg/mL), SUMO-Gm94 (21.4 μg/mL), AP-64 (7.2 μg/mL), or Gm94 (7.6 μg/mL) (1 μM, the same order of magnitude as known for other AMPs) to DH5α cells (100 µL), bacteria were incubated at 37 °C for 4 h and then diluted with LB. The 100 µL samples of the dilutions were plated onto LB agar plates and incubated at 37 °C for 16 h. The final number of colony-forming units (CFUs) was counted.
Meanwhile, a zone of inhibition test of the proteins against
E. coli DH5α was performed using the Oxford cup method [
14]. Briefly, 100 μL of diluted inoculum (10
5 CFU/mL) from bacteria suspensions was added to and spread on the surface of LB agar plates. Sterilized Oxford cups (Φ 5 mm) were then placed on the agar medium and filled with 100 µL of SUMO-AP-64, SUMO-Gm94, AP-64, or Gm94 protein (1 μM); the plates were incubated at 37 °C for 18 h. An equivalent volume of PBS was used as a control.
2.4. Antibacterial Activity of AP-64 and Gm94 against DH5α
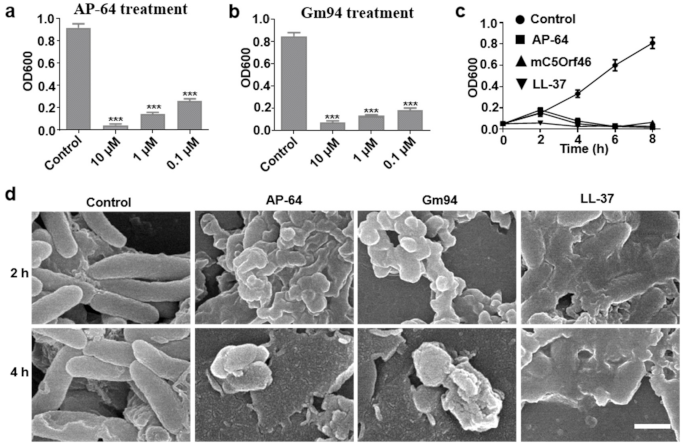
To evaluate the inhibition effect of different concentrations of AP-64 and Gm94 on growth of DH5α, the bacteria were cultured in LB at 37 °C until the OD600 reached 0.5 and were then diluted 10-fold. Subsequently, 100 μL of the cell culture was added to each well of microtiter plates containing LB medium supplemented with AP-64 or Gm94 (0.1–10 μM). After incubation at 37 °C for 8 h, OD600 measurements were performed on a universal microplate spectrophotometer (BioTek, Winooski, VT, USA).
Next, growth curve analysis was implemented. DH5α was exposed to AP-64, Gm94, and LL-37 (61302; AnaSpec, Beijing, China) at a concentration of 10 μM for 8 h, and the OD600 values were measured every 2 h. DH5α was used as the control.
2.5. SEM Detection for DH5α Cells
DH5α cells were treated with the proteins at a concentration of 10 μM and subjected to scanning electron microscopy (SEM) according to a method reported previously [
15]. Briefly, treated bacteria were harvested at different time points and centrifuged at 5000×
g for 4 min. The bacteria were fixed with 2% glutaraldehyde for 2 h before a thorough wash with PBS buffer. The fixed bacteria were dehydrated through an ascending alcohol series (30%, 50%, 65%, 85%, 90%, and 100% EtOH) for 10 min each. After a critical-point drying, the samples were visualized using SEM. DH5α cells treated with PBS were used as the blank control.
2.6. Antibacterial Activity against Pathogenic Bacteria
Escherichia coli O157:H7 (ATCC, 43895), Vibrio cholerae (ATCC, 51394), Pseudomonas aeruginosa (ATCC, 27853), Staphylococcus aureus (ATCC, 25923), and Listeria monocytogenes (ATCC, 19115) cells were treated using the method described above.
The minimum inhibitory concentrations of AP-64 or Gm94 were measured in the concentration range of 0.2–200 μM by standard double dilution methods [
16] and were taken as the lowest concentration of peptide at which no visible growth was observed (
Table S1).
2.7. Antibacterial Activity against Escherichia coli O157:H7 in Mouse
Male BALB/c mice (6 weeks, 18–22 g) were obtained from GemPharmatech (Jiangsu, China). Mice were randomly assigned to 4 groups with 10 mice in each group. O157:H7 (5 × 106 CFU/mouse) was administered intraperitoneally to the mice. Thirty minutes later, the mice were injected intraperitoneally with AP-64, Gm94, or LL-37 at the dose of 500 μg/kg. An equivalent volume of PBS was used as a control. All animal experiments were approved by the West China Hospital of Sichuan University Biomedical Ethics Committee (ethic approval document: 2020166A), and all experiments conformed to all relevant regulator standards.
2.8. Antitumor Capacity
Human tumor cells (2 × 104) were cultured in RPMI 1640 or DMEM medium containing 5% fetal bovine serum with AP-64 or Gm94 at the indicated concentration. After treatment at 37 °C for 24 h, cells were stained using a Cell Counting Kit-8 (CCK-8) according to the manufacturer’s protocol (C0037; Beyotime, Shanghai, China). The following formula was used to calculate the inhibition rate of cell growth (%): cell growth inhibition (%) = (1 − (percentage of surviving cells in the AP-64 or Gm94 treatment group/percentage of surviving cells in the PBS control group)) × 100. PBS-treated cells served as a control in all experiments.
2.9. Statistical Analysis
Data are expressed as the mean ± standard deviation from three independent experiments. Statistical analyses were performed using the GraphPad Prism Software version 5.0. For in vitro analysis, differences between experimental conditions were analyzed using unpaired Student’s t-test. Significance was set at p < 0.05. For in vivo data, a survival curve of mice was obtained using the Kaplan–Meier plot with a log-rank test. Significance was set at p < 0.05.
4. Discussion
Despite the many efforts that have been devoted to the development of effective antibacterial agents, certain mammalian proteins with intrinsic antibacterial activity might be underappreciated. Here, we demonstrated that AP-64 (human C5orf46) is a type of AMP that exerts a direct antibacterial effect on Gram-negative bacteria, and our data shows that Gm94 (mouse C5orf46) also exhibits the similar antibacterial ability.
Although AMPs are promising candidates for the development of novel antibiotics, it is difficult to produce sufficient quantities of these proteins because of their toxicity toward microbial expression hosts. Previously, the SUMO tag was used widely for the prokaryotic expression of recombinant proteins, such as abaecin (a 34 amino acid long antimicrobial peptide from honeybees) [
17]. In this study, the SUMO tag was fused to the N terminus of AP-64 to express the SUMO-AP-64 fusion protein. SUMO-AP-64 was expressed in a soluble form but failed to inhibit the growth of DH5α cells. After removal of the SUMO tag, AP-64 exhibited strong antibacterial effects. Thus, fusing a large tag, such as SUMO, to the N terminus of AP-64 blocked its antibacterial activity. We observed that bacteria treated with AP-64 underwent a slow multiplication at the beginning of the culturing process, followed by a rapid decline. This growth curve implied that AP-64 possessed a similar antibacterial pattern to that of cell-envelope-interfering agents and that its action might be related to cell envelope damage [
18]. The multiplication of growth might also hint at an intracellular target of the peptide. It might need some time to become internalized into the cells before it can exert its antibacterial function.
A bioinformatics analysis using TCGA showed that AP-64 is present not only at the RNA level in the skin and salivary glands, but also in major blood vessels (aorta) and the heart [
19]. Notably, mice that were heterozygous for a null mutation of Gm94 had a significantly increased fat mass and reduced lean mass compared with wild-type mice [
20]. Recently, AP-64 was identified as a plasma protein through deep-proteome profiling of human plasma [
12]. Thus, whether AP-64 plays a role in lipid homeostasis is worthy of further study.
AP-64 exhibits some distinct features compared with some human AMPs (defensins, LL-37, histatins, and hepcidin). First, compared with other human AMPs, which normally contain less than 45 amino acid residues, AP-64 consists of 64 residues. Second, while most of the human AMPs are highly cationic, AP-64 is an anionic peptide with a PI of 4.54. Third, unlike other human AMPs (such as defensins and hepcidin), AP-64 contains no cysteines. Meanwhile, AP-64 also exhibits similar features to other human AMPs. For example, both AP-64 and dermcidin have a net negative charge and contain no cysteines. In addition, similar to Microcin E492, AP-64 possesses activity against Gram-negative bacteria and tumor cells (
https://wangapd3.com/main.php, accessed on 16 March 2021).
Although our study revealed that AP-64 exhibits strong antibacterial activity against Gram-negative bacteria, it had several limitations. First, the mRNA expression of AP-64 in the skin and salivary gland was discovered using the TCGA database; thus, a specific antibody should be generated, and this protein’s expression profile should be further investigated in detail by immunohistochemical staining. Second, the molecular mechanisms underlying the bacterial inhibition should be investigated in future studies. Third, our research only confirmed the antibacterial effect of AP-64 on the bacteria used in this study, but other bacteria and fungi should also be tested.
In summary, we demonstrated for the first time that AP-64/C5orf46 is a type of AMP. Compared with other human AMPs, AP-64 possesses distinctive characteristics, including a longer sequence length, absence of cysteine residues, highly anionic characteristics, and cell toxicity against Jurkat and Raji cells. Our data provided useful clues for further study of the physiological functions of AP-64/C5orf46.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




