
MRAP2 Interaction with Melanocortin-4 Receptor in SnakeHead (Channa argus)

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Sampling
2.2. Ligands and Plasmids
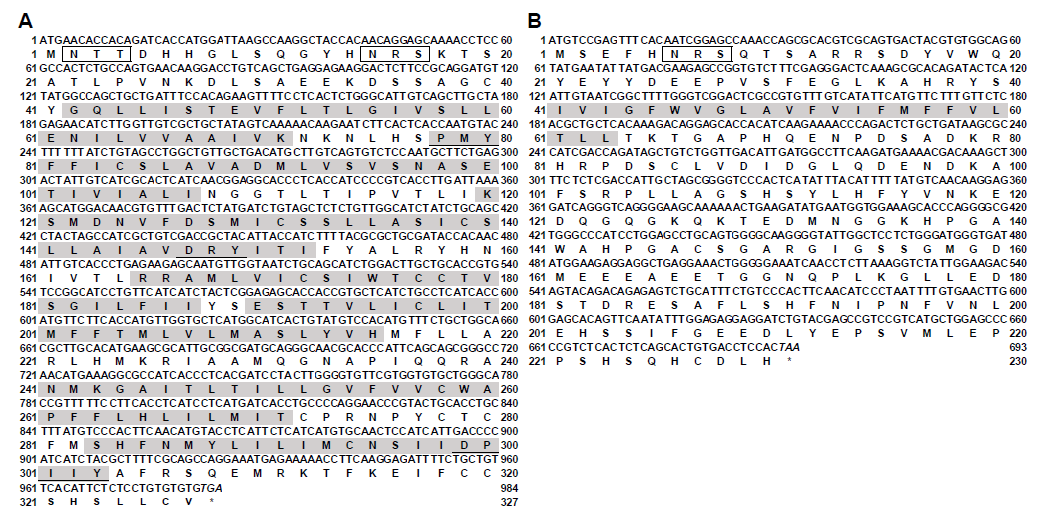
2.3. Molecular Cloning of Snakehead mc4r and mrap2 Genes
2.4. Sequence Analysis and Data Processing
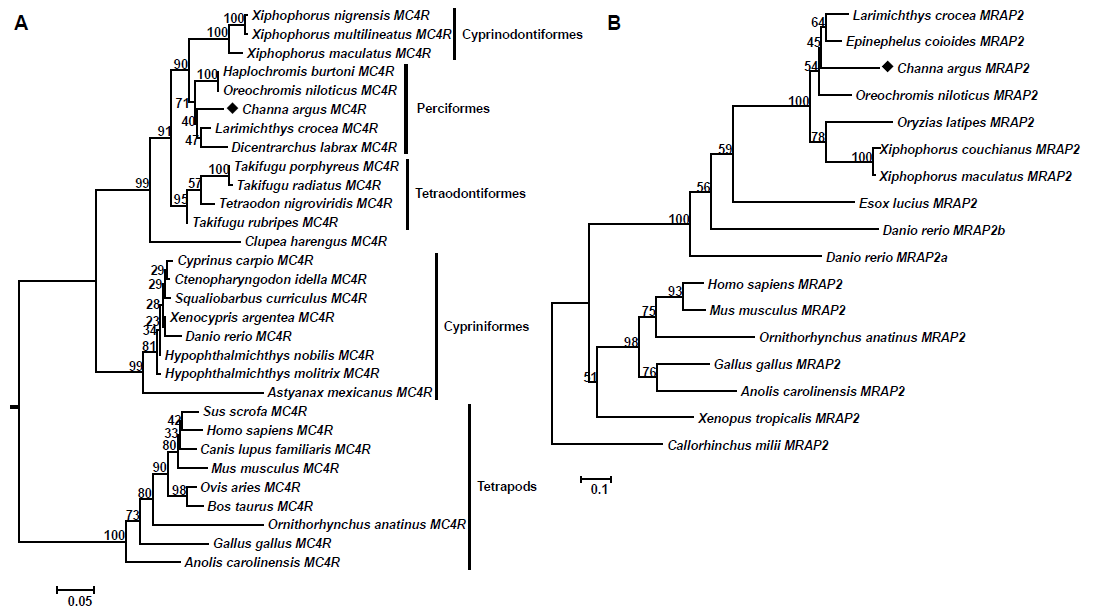
2.5. Phylogenetic Analysis
2.6. Quantitative Real-Time PCR
2.7. Cell Culture and Transfection
2.8. Flow Cytometry Assay
2.9. Ligand Binding Assays
2.10. cAMP Assays
2.11. Statistical Analysis
3. Results
3.1. Molecular Cloning of Snakehead mc4r and mrap2 Genes
3.2. Comparative Synteny of mc4r and mrap2 in Various Vertebrate Genomes
3.3. Phylogenetic Analysis
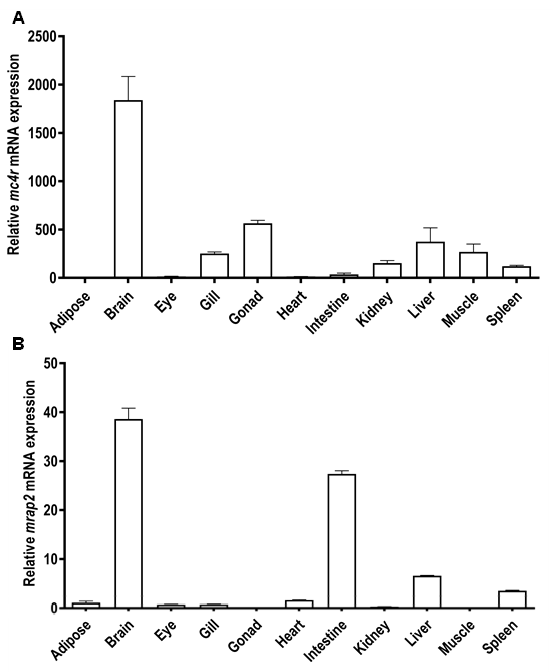
3.4. Tissue Distribution of Snakehead mc4r and mrap2 Genes
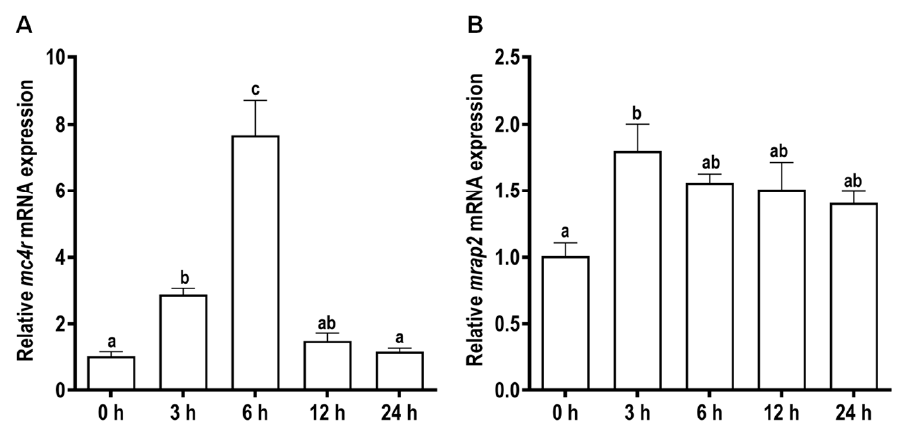
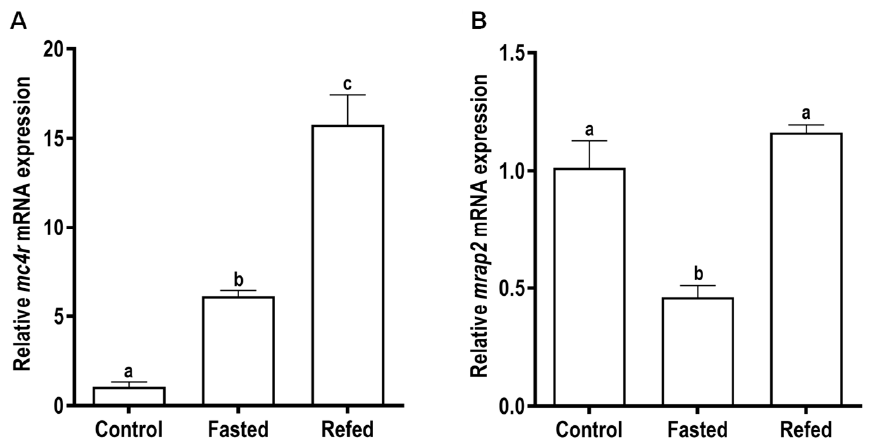
3.5. Effects of Fasting and Refeeding on the Expressions of Snakehead mc4r and mrap2 Genes
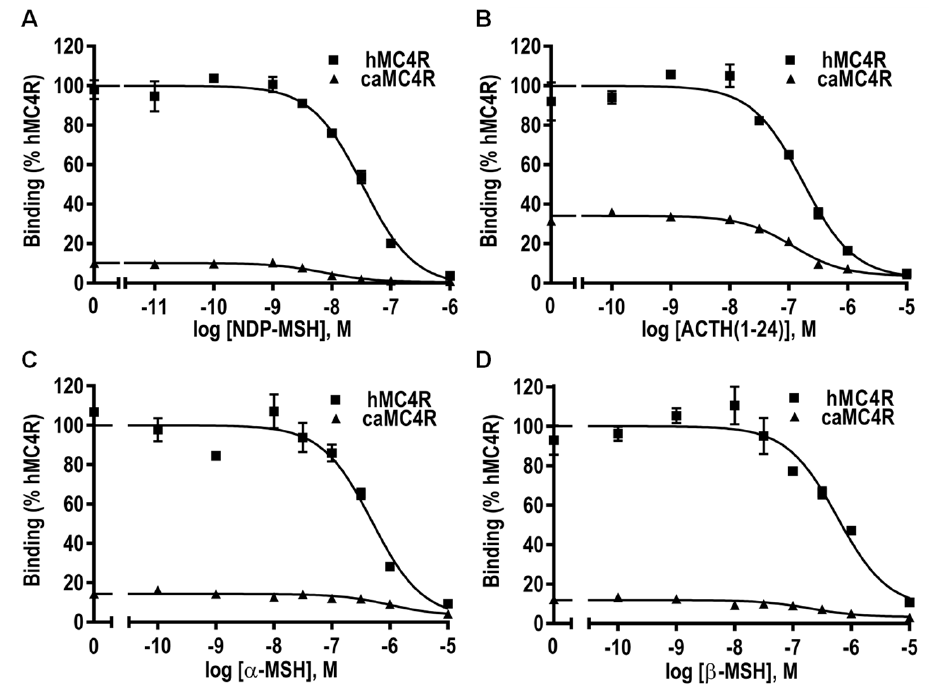
3.6. Ligand Binding Properties of caMC4R
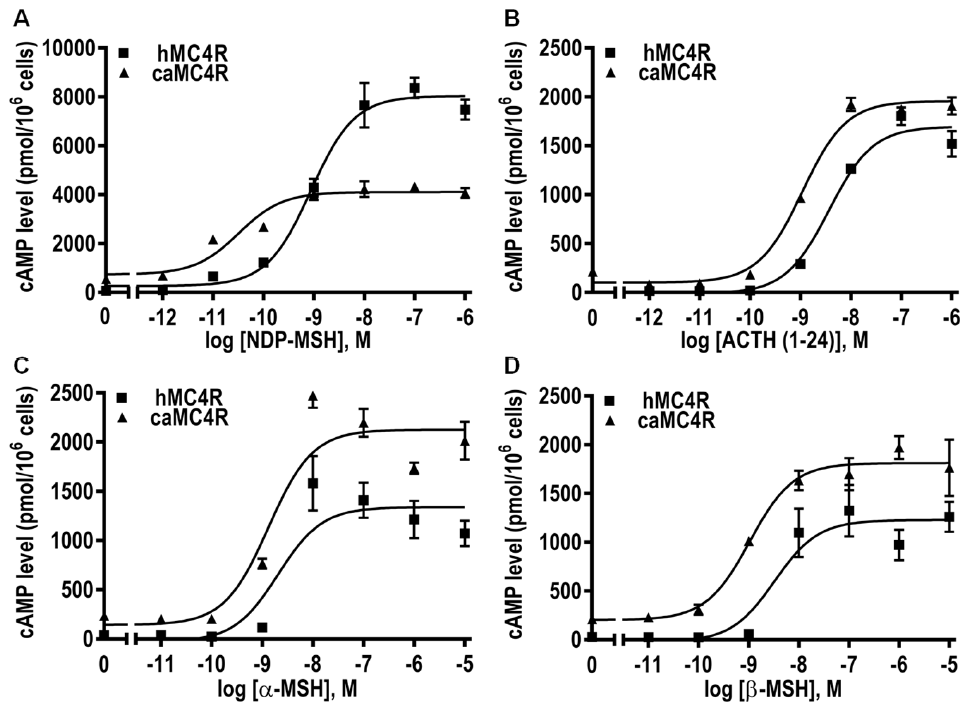
3.7. Signaling Properties of caMC4R
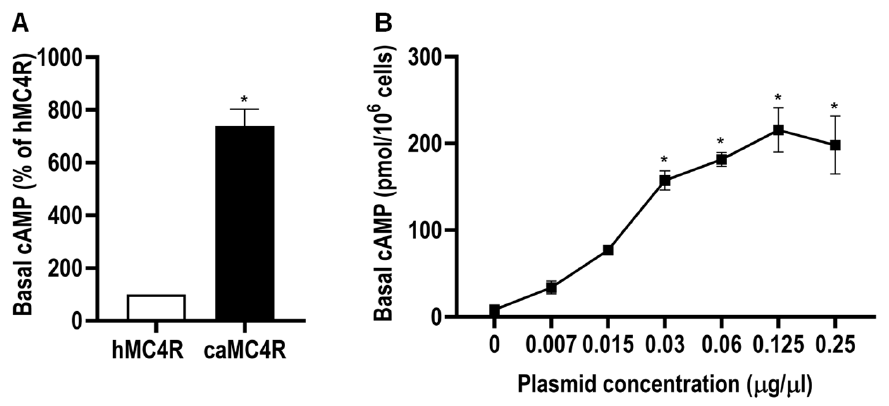
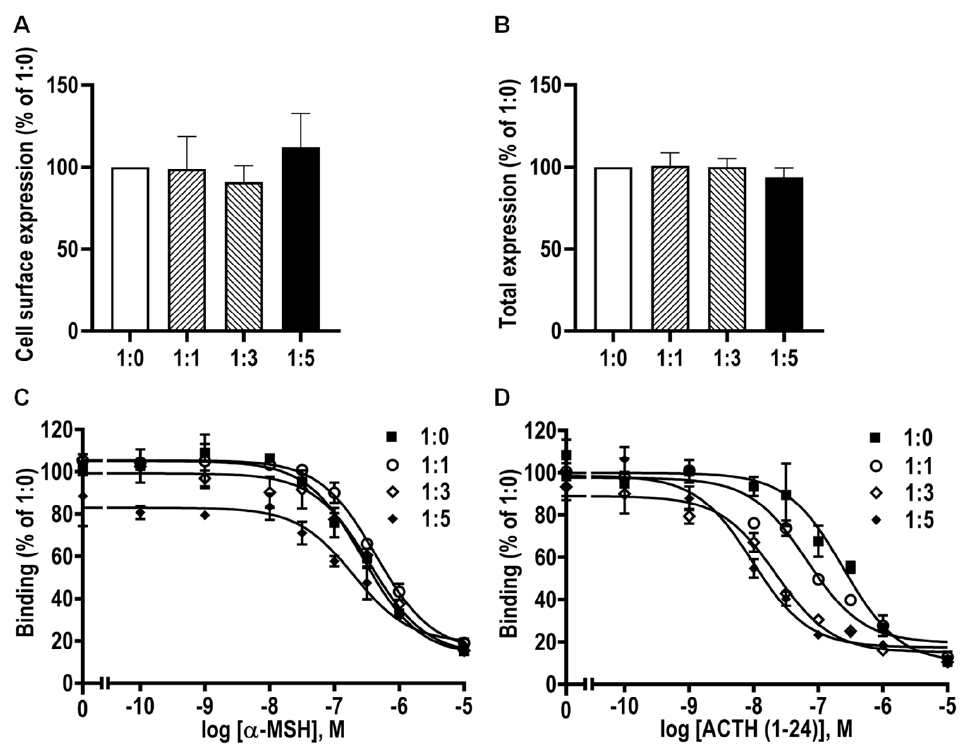
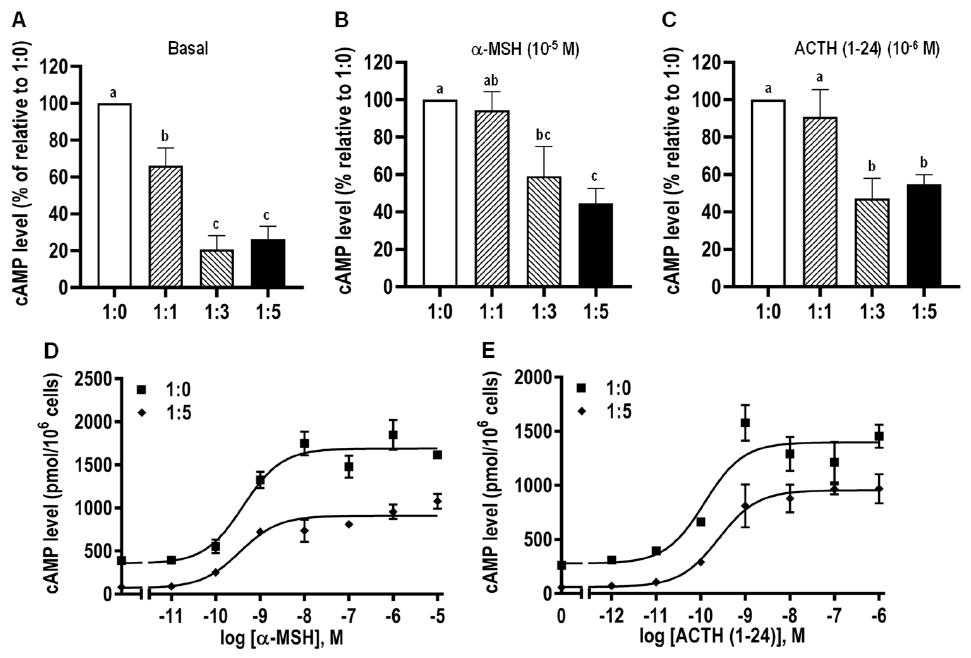
3.8. Modulation of caMC4R Expression and Pharmacological Properties by caMRAP2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Cone, R.D. Anatomy and regulation of the central melanocortin system. Nat. Neurosci. 2005, 8, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.X. The melanocortin-4 receptor: Physiology, pharmacology, and pathophysiology. Endocr. Rev. 2010, 31, 506–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gantz, I.; Miwa, H.; Konda, Y.; Shimoto, Y.; Tashiro, T.; Watson, S.J.; DelValle, J.; Yamada, T. Molecular cloning, expression, and gene localization of a fourth melanocortin receptor. J. Biol. Chem. 1993, 268, 15174–15179. [Google Scholar] [CrossRef]
- Mountjoy, K.G.; Mortrud, M.T.; Low, M.J.; Simerly, R.B.; Cone, R.D. Localization of the melanocortin-4 receptor (MC4-R) in neuroendocrine and autonomic control circuits in the brain. Mol. Endocrinol. 1994, 8, 1298–1308. [Google Scholar] [PubMed] [Green Version]
- Fan, W.; Boston, B.A.; Kesterson, R.A.; Hruby, V.J.; Cone, R.D. Role of melanocortinergic neurons in feeding and the agouti obesity syndrome. Nature 1997, 385, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Ollmann, M.M.; Wilson, B.D.; Yang, Y.K.; Kerns, J.A.; Chen, Y.; Gantz, I.; Barsh, G.S. Antagonism of central melanocortin receptors in vitro and in vivo by agouti-related protein. Science 1997, 278, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Huszar, D.; Lynch, C.A.; Fairchild-Huntress, V.; Dunmore, J.H.; Fang, Q.; Berkemeier, L.R.; Gu, W.; Kesterson, R.A.; Boston, B.A.; Cone, R.D.; et al. Targeted disruption of the melanocortin-4 receptor results in obesity in mice. Cell 1997, 88, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Vaisse, C.; Clement, K.; Guy-Grand, B.; Froguel, P. A frameshift mutation in human MC4R is associated with a dominant form of obesity. Nat. Genet. 1998, 20, 113–114. [Google Scholar] [CrossRef]
- Yeo, G.S.; Farooqi, I.S.; Aminian, S.; Halsall, D.J.; Stanhope, R.G.; O’Rahilly, S. A frameshift mutation in MC4R associated with dominantly inherited human obesity. Nat. Genet. 1998, 20, 111–112. [Google Scholar] [CrossRef]
- Loos, R.J.; Lindgren, C.M.; Li, S.; Wheeler, E.; Zhao, J.H.; Prokopenko, I.; Inouye, M.; Freathy, R.M.; Attwood, A.P.; Beckmann, J.S.; et al. Common genetic variation near MC4R is associated with waist circumference and insulin resistance. Nat. Genet. 2008, 40, 716–718. [Google Scholar]
- Van der Ploeg, L.H.; Martin, W.J.; Howard, A.D.; Nargund, R.P.; Austin, C.P.; Guan, X.; Drisko, J.; Cashen, D.; Sebhat, I.; Patchett, A.A.; et al. A role for the melanocortin 4 receptor in sexual function. Proc. Natl. Acad. Sci. USA 2002, 99, 11381–11386. [Google Scholar] [CrossRef] [Green Version]
- Irani, B.G.; Xiang, Z.; Moore, M.C.; Mandel, R.J.; Haskell-Luevano, C. Voluntary exercise delays monogenetic obesity and overcomes reproductive dysfunction of the melanocortin-4 receptor knockout mouse. Biochem. Biophys. Res. Commun. 2005, 326, 638–644. [Google Scholar] [CrossRef]
- Sandrock, M.; Schulz, A.; Merkwitz, C.; Schoneberg, T.; Spanel-Borowski, K.; Ricken, A. Reduction in corpora lutea number in obese melanocortin-4-receptor-deficient mice. Reprod. Biol. Endocrinol. 2009, 7, 24. [Google Scholar] [CrossRef] [Green Version]
- Agosti, F.; López Soto, E.J.; Cabral, A.; Castrogiovanni, D.; Schioth, H.B.; Perelló, M.; Raingo, J. Melanocortin 4 receptor activation inhibits presynaptic N-type calcium channels in amygdaloid complex neurons. Eur. J. Neurosci. 2014, 40, 2755–2765. [Google Scholar] [CrossRef]
- Podyma, B.; Sun, H.; Wilson, E.A.; Carlson, B.; Pritikin, E.; Gavrilova, O.; Weinstein, L.S.; Chen, M. The stimulatory G protein Gsalpha is required in melanocortin 4 receptor-expressing cells for normal energy balance, thermogenesis, and glucose metabolism. J. Biol. Chem. 2018, 293, 10993–11005. [Google Scholar] [CrossRef] [Green Version]
- Cortés, R.; Agulleiro, M.J.; Navarro, S.; Guillot, R.; Sánchez, E.; Cerdá-Reverter, J.M. Melanocortin receptor accessory protein 2 (MRAP2) interplays with the zebrafish melanocortin 1 receptor (MC1R) but has no effect on its pharmacological profile. Gen. Comp. Endocrinol. 2014, 201, 30–36. [Google Scholar] [CrossRef]
- Cerdá-Reverter, J.M.; Agulleiro, M.J.; Cortés, R.; Sánchez, E.; Guillot, R.; Leal, E.; Fernández-Durán, B.; Puchol, S.; Eley, M. Involvement of melanocortin receptor accessory proteins (MRAPs) in the function of melanocortin receptors. Gen. Comp. Endocrinol. 2013, 188, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Rouault, A.A.J.; Srinivasan, D.K.; Yin, T.C.; Lee, A.A.; Sebag, J.A. Melanocortin receptor accessory proteins (MRAPs): Functions in the melanocortin system and beyond. Biochim. Biophys. Acta 2017, 1864, 2322–2329. [Google Scholar] [CrossRef] [PubMed]
- Schonnop, L.; Kleinau, G.; Herrfurth, N.; Volckmar, A.L.; Cetindag, C.; Müller, A.; Peters, T.; Herpertz, S.; Antel, J.; Hebebrand, J.; et al. Decreased melanocortin-4 receptor function conferred by an infrequent variant at the human melanocortin receptor accessory protein 2 gene. Obesity 2016, 24, 1976–1982. [Google Scholar] [CrossRef]
- Asai, M.; Ramachandrappa, S.; Joachim, M.; Shen, Y.; Zhang, R.; Nuthalapati, N.; Ramanathan, V.; Strochlic, D.E.; Ferket, P.; Linhart, K.; et al. Loss of function of the melanocortin 2 receptor accessory protein 2 is associated with mammalian obesity. Science 2013, 341, 275–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, L.F.; Webb, T.R.; Chung, T.T.; Meimaridou, E.; Cooray, S.N.; Guasti, L.; Chapple, J.P.; Egertova, M.; Elphick, M.R.; Cheetham, M.E.; et al. MRAP and MRAP2 are bidirectional regulators of the melanocortin receptor family. Proc. Natl. Acad. Sci. USA 2009, 106, 6146–6151. [Google Scholar] [CrossRef] [Green Version]
- Habara, M.; Mori, N.; Okada, Y.; Kawasumi, K.; Nakao, N.; Tanaka, Y.; Arai, T.; Yamamoto, I. Molecular characterization of feline melanocortin 4 receptor and melanocortin 2 receptor accessory protein 2. Gen. Comp. Endocrinol. 2018, 261, 31–39. [Google Scholar] [CrossRef]
- Bruschetta, G.; Kim, J.D.; Diano, S.; Chan, L.F. Overexpression of melanocortin 2 receptor accessory protein 2 (MRAP2) in adult paraventricular MC4R neurons regulates energy intake and expenditure. Mol. Metab. 2018, 18, 79–87. [Google Scholar] [CrossRef]
- Cerdá-Reverter, J.M.; Ringholm, A.; Schioth, H.B.; Peter, R.E. Molecular cloning, pharmacological characterization, and brain mapping of the melanocortin 4 receptor in the goldfish: Involvement in the control of food intake. Endocrinology 2003, 144, 2336–2349. [Google Scholar] [CrossRef] [Green Version]
- Haitina, T.; Klovins, J.; Andersson, J.; Fredriksson, R.; Lagerstrom, M.C.; Larhammar, D.; Larson, E.T.; Schioth, H.B. Cloning, tissue distribution, pharmacology and three-dimensional modelling of melanocortin receptors 4 and 5 in rainbow trout suggest close evolutionary relationship of these subtypes. Biochem. J. 2004, 380, 475–486. [Google Scholar] [CrossRef] [Green Version]
- Li, J.T.; Yang, Z.; Chen, H.P.; Zhu, C.H.; Deng, S.P.; Li, G.L.; Tao, Y.X. Molecular cloning, tissue distribution, and pharmacological characterization of melanocortin-4 receptor in spotted scat, Scatophagus argus. Gen. Comp. Endocrinol. 2016, 230–231, 143–152. [Google Scholar] [CrossRef]
- Li, L.; Yang, Z.; Zhang, Y.P.; He, S.; Liang, X.F.; Tao, Y.X. Molecular cloning, tissue distribution, and pharmacological characterization of melanocortin-4 receptor in grass carp (Ctenopharyngodon idella). Domest Anim Endocrinol. 2017, 59, 140–151. [Google Scholar] [CrossRef]
- Yi, T.L.; Yang, L.K.; Ruan, G.L.; Yang, D.Q.; Tao, Y.X. Melanocortin-4 receptor in swamp eel (Monopterus albus): Cloning, tissue distribution, and pharmacology. Gene 2018, 678, 79–89. [Google Scholar] [CrossRef]
- Rao, Y.Z.; Chen, R.; Zhang, Y.; Tao, Y.X. Orange-spotted grouper melanocortin-4 receptor: Modulation of signaling by MRAP2. Gen. Comp. Endocrinol. 2019, 284, 113234. [Google Scholar] [CrossRef]
- Tao, M.; Ji, R.L.; Huang, L.; Fan, S.Y.; Liu, T.; Liu, S.J.; Tao, Y.X. Regulation of melanocortin-4 receptor pharmacology by two isoforms of melanocortin receptor accessory protein 2 in topmouth culter (Culter alburnus). Front. Endocrinol. 2020, 11, 538. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Tsuchiya, K.; Yamanome, T.; Schioth, H.B.; Kawauchi, H.; Takahashi, A. Food deprivation increases the expression of melanocortin-4 receptor in the liver of barfin flounder, Verasper moseri. Gen. Comp. Endocrinol. 2008, 155, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Yuan, D.; Zhou, C.; Wang, T.; Lin, F.; Chen, H.; Wu, H.; Xin, Z.; Yang, S.; Chen, D.; et al. Cloning, distribution and effects of fasting status of melanocortin 4 receptor (MC4R) in Schizothorax prenanti. Gene 2013, 532, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Aspiras, A.C.; Rohner, N.; Martineau, B.; Borowsky, R.L.; Tabin, C.J. Melanocortin 4 receptor mutations contribute to the adaptation of cavefish to nutrient-poor conditions. Proc. Natl. Acad. Sci. USA 2015, 112, 9668–9673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebag, J.A.; Zhang, C.; Hinkle, P.M.; Bradshaw, A.M.; Cone, R.D. Developmental control of the melanocortin-4 receptor by MRAP2 proteins in zebrafish. Science 2013, 341, 278–281. [Google Scholar] [CrossRef] [Green Version]
- Josep Agulleiro, M.; Cortes, R.; Fernandez-Duran, B.; Navarro, S.; Guillot, R.; Meimaridou, E.; Clark, A.J.; Cerdá-Reverter, J.M. Melanocortin 4 receptor becomes an ACTH receptor by coexpression of melanocortin receptor accessory protein 2. Mol. Endocrinol. 2013, 27, 1934–1945. [Google Scholar] [CrossRef]
- Tao, Y.X. Molecular chaperones and G protein-coupled receptor maturation and pharmacology. Mol. Cell Endocrinol. 2020, 511, 110862. [Google Scholar] [CrossRef]
- Zhu, M.; Xu, B.; Wang, M.; Liu, S.; Zhang, Y.; Zhang, C. Pharmacological modulation of MRAP2 protein on melanocortin receptors in the sea lamprey. Endocr. Connect 2019, 8, 378–388. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Chen, Y.J.; Zhu, M.; Xu, B.X.; Guo, W.X.; Lyu, Y.S.; Zhang, C. Pharmacological modulation of melanocortin-4 receptor by melanocortin receptor accessory protein 2 in Nile tilapia. Gen. Comp. Endocrinol. 2019, 282, 113219. [Google Scholar] [CrossRef]
- Wen, Z.Y.; Qin, C.J.; Wang, J.; He, Y.; Li, H.T.; Li, R.; Wang, X.D. Molecular characterization of two leptin genes and their transcriptional changes in response to fasting and refeeding in Northern snakehead (Channa argus). Gene 2020, 736, 144420. [Google Scholar] [CrossRef]
- Xu, J.; Bian, C.; Chen, K.C.; Liu, G.M.; Jiang, Y.L.; Luo, Q.; You, X.X.; Peng, W.Z.; Li, J.; Huang, Y.; et al. Draft genome of the Northern snakehead, Channa argus. Giga Sci. 2017, 6, gix011. [Google Scholar] [CrossRef]
- Mo, X.L.; Yang, R.; Tao, Y.X. Functions of transmembrane domain 3 of human melanocortin-4 receptor. J. Mol. Endocrinol. 2012, 49, 221–235. [Google Scholar] [CrossRef] [Green Version]
- Steiner, A.L.; Kipnis, D.M.; Utiger, R.; Parker, C. Radioimmunoassay for the measurement of adenosine 3’,5’-cyclic phosphate. Proc. Natl. Acad. Sci. USA 1969, 64, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.X.; Segaloff, D.L. Functional characterization of melanocortin-4 receptor mutations associated with childhood obesity. Endocrinology 2003, 144, 4544–4551. [Google Scholar] [CrossRef]
- Wen, Z.Y.; Liang, X.F.; He, S.; Li, L.; Shen, D.; Tao, Y.X. Molecular cloning and tissue expression of uncoupling protein 1, 2 and 3 genes in Chinese perch (Siniperca chuatsi). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2015, 185, 24–33. [Google Scholar] [CrossRef]
- Yu, J.; Gimenez, L.E.; Hernandez, C.C.; Wu, Y.; Wein, A.H.; Han, G.W.; McClary, K.; Mittal, S.R.; Burdsall, K.; Stauch, B.; et al. Determination of the melanocortin-4 receptor structure identifies Ca2+ as a cofactor for ligand binding. Science 2020, 368, 428–433. [Google Scholar] [CrossRef]
- Wen, Z.Y.; Wang, J.; Bian, C.; Zhang, X.; Li, J.; Peng, Y.; Zhan, Q.; Shi, Q.; Li, Y.Y. Molecular cloning of two kcnk3 genes from the Northern snakehead (Channa argus) for quantification of their transcriptions in response to fasting and refeeding. Gen. Comp. Endocrinol. 2019, 281, 49–57. [Google Scholar] [CrossRef]
- Wen, Z.Y.; Bian, C.; You, X.; Zhang, X.; Li, J.; Zhan, Q.; Peng, Y.; Li, Y.Y.; Shi, Q. Characterization of two kcnk3 genes in Nile tilapia (Oreochromis niloticus): Molecular cloning, tissue distribution, and transcriptional changes in various salinity of seawater. Genomics 2020, 112, 2213–2222. [Google Scholar] [CrossRef]
- Wen, Z.; Li, Y.; Bian, C.; Shi, Q.; Li, Y. Characterization of two kcnk3 genes in rabbitfish (Siganus canaliculatus): Molecular cloning, distribution patterns and their potential roles in fatty acids metabolism and osmoregulation. Gen. Comp. Endocrinol. 2020, 296, 113546. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrAIC. pl. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Abascal, F.; Zardoya, R.; Posada, D. ProtTest: Selection of best-fit models of protein evolution. Bioinformatics 2005, 21, 2104–2105. [Google Scholar] [CrossRef] [Green Version]
- Qin, C.J.; Wen, Z.Y.; Wang, J.; He, Y.; Yuan, D.Y.; Li, R. Uncoupling protein 1 in snakehead (Channa argus): Cloning, tissue distribution, and its expression in response to fasting and refeeding. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2018, 225, 1–6. [Google Scholar] [CrossRef]
- Chen, C.; Okayama, H. High-efficiency transformation of mammalian cells by plasmid DNA. Mol. Cell Biol. 1987, 7, 2745–2752. [Google Scholar] [CrossRef]
- Wang, S.X.; Fan, Z.C.; Tao, Y.X. Functions of acidic transmembrane residues in human melanocortin-3 receptor binding and activation. Biochem. Pharmacol. 2008, 76, 520–530. [Google Scholar] [CrossRef] [Green Version]
- Hughes, L.C.; Orti, G.; Huang, Y.; Sun, Y.; Baldwin, C.C.; Thompson, A.W.; Arcila, D.; Betancur, R.R.; Li, C.; Becker, L.; et al. Comprehensive phylogeny of ray-finned fishes (Actinopterygii) based on transcriptomic and genomic data. Proc. Natl. Acad. Sci. USA 2018, 115, 6249–6254. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.; Zhang, Y.; Ji, P.; Li, Y.; Xu, P.; Sun, X. Molecular characterization of CART, AgRP, and MC4R genes and their expression with fasting and re-feeding in common carp (Cyprinus carpio). Mol. Biol. Rep. 2012, 39, 2215–2223. [Google Scholar] [CrossRef]
- Lampert, K.P.; Schmidt, C.; Fischer, P.; Volff, J.N.; Hoffmann, C.; Muck, J.; Lohse, M.J.; Ryan, M.J.; Schartl, M. Determination of onset of sexual maturation and mating behavior by melanocortin receptor 4 polymorphisms. Curr. Biol. 2010, 20, 1729–1734. [Google Scholar] [CrossRef]
- Jangprai, A.; Boonanuntanasarn, S.; Yoshizaki, G. Characterization of melanocortin 4 receptor in Snakeskin Gourami and its expression in relation to daily feed intake and short-term fasting. Gen. Comp. Endocrinol. 2011, 173, 27–37. [Google Scholar] [CrossRef]
- Zhang, J.; Li, X.; Zhou, Y.; Cui, L.; Li, J.; Wu, C.; Wan, Y.; Li, J.; Wang, Y. The interaction of MC3R and MC4R with MRAP2, ACTH, α-MSH and AgRP in chickens. J. Endocrinol. 2017, 234, 155–174. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.Q.; Hou, Z.S.; Wen, H.S.; Li, Y.; Qi, X.; Li, W.J.; Tao, Y.X. Melanocortin-4 receptor in spotted sea bass, Lateolabrax maculatus: Cloning, tissue distribution, physiology, and pharmacology. Front. Endocrinol. 2019, 10, 705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Y.X. Functional characterization of novel melanocortin-3 receptor mutations identified from obese subjects. Biochim. Biophys. Acta 2007, 1772, 1167–1174. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, S.; Lubrano-Berthelier, C.; Govaerts, C.; Picard, F.; Santiago, P.; Conklin, B.R.; Vaisse, C. Constitutive activity of the melanocortin-4 receptor is maintained by its N-terminal domain and plays a role in energy homeostasis in humans. J. Clin. Investig. 2004, 114, 1158–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Y.X. Constitutive activation of G protein-coupled receptors and diseases: Insights into mechanism of activation and therapeutics. Pharmacol. Ther. 2008, 120, 129–148. [Google Scholar] [CrossRef] [Green Version]
- Sebag, J.A.; Hinkle, P.M. Regions of melanocortin 2 (MC2) receptor accessory protein necessary for dual topology and MC2 receptor trafficking and signaling. J. Biol. Chem. 2009, 284, 610–618. [Google Scholar] [CrossRef] [Green Version]
- Soletto, L.; Hernandez-Balfago, S.; Rocha, A.; Scheerer, P.; Kleinau, G.; Cerdá-Reverter, J.M. Melanocortin receptor accessory protein 2-induced adrenocorticotropic hormone response of human melanocortin 4 receptor. J. Endocr. Soc. 2019, 3, 314–323. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.N.; Li, J.T.; Tao, Y.X.; Chen, H.P.; Deng, S.P.; Zhu, C.H.; Li, G.L. Effects of melanocortin-4 receptor agonists and antagonists on expression of genes related to reproduction in spotted scat, Scatophagus argus. J. Comp. Physiol. B 2017, 187, 603–612. [Google Scholar] [CrossRef]
- Zhang, Y.; Wen, H.S.; Li, Y.; Lyu, L.K.; Zhang, Z.X.; Wang, X.J.; Li, J.S.; Tao, Y.X.; Qi, X. Melanocortin-4 receptor regulation of reproductive function in black rockfish (Sebastes schlegelii). Gene 2020, 741, 144541. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MC4R | Bmax (%) | NDP-MSH | ACTH (1–24) | α-MSH | β-MSH |
|---|---|---|---|---|---|
| IC50 (nM) | IC50 (nM) | IC50 (nM) | IC50 (nM) | ||
| hMC4R | 100 | 27.72 ± 4.29 | 324.6 ± 88.97 | 488.2 ± 2.55 | 749.4 ± 93.98 |
| caMC4R | 16.32 ± 2.46 c | 4.50 ± 1.07 a | 415.1 ± 163.7 | 981.0 ± 45.52 b | 328.7 ± 119.6 |
| MC4R | hMC4R | caMC4R | |
|---|---|---|---|
| Basal (%) | 100 | 739.2 ± 63.82 b | |
| NDP-MSH | EC50 (nM) | 0.59 ± 0.18 | 0.04 ± 0.02 |
| Rmax (%) | 100 | 75.18 ± 27.83 | |
| ACTH (1–24) | EC50 (nM) | 2.68 ± 0.50 | 0.81 ± 0.18 a |
| Rmax (%) | 100 | 119.0 ± 3.73 a | |
| α-MSH | EC50 (nM) | 2.82 ± 0.40 | 2.32 ± 0.57 |
| Rmax (%) | 100 | 198.1 ± 55.95 | |
| β-MSH | EC50 (nM) | 6.67 ± 3.41 | 2.49 ± 0.81 |
| Rmax (%) | 100 | 163.9 ± 89.36 |
| caMC4R/caMRAP2 | Bmax (%) | α-MSH | ACTH (1–24) |
|---|---|---|---|
| IC50 (nM) | IC50 (nM) | ||
| 1:0 | 100 | 383.80 ± 68.82 | 239.90 ± 23.61 |
| 1:1 | 100.70 ± 6.24 | 461.00 ± 65.51 | 144.80 ± 74.73 |
| 1:3 | 102.80 ± 9.56 | 497.10 ± 147.4 | 49.48 ± 19.47 a |
| 1:5 | 83.62 ± 7.17 | 238.10 ± 60.77 | 24.73 ± 11.73 a |
| caMC4R/caMRAP2 | α-MSH | ACTH (1–24) | ||
|---|---|---|---|---|
| EC50 (nM) | Rmax (%) | EC50 (nM) | Rmax (%) | |
| 1:0 | 0.48 ± 0.12 | 100 | 0.45 ± 0.20 | 100 |
| 1:5 | 0.61 ± 0.15 | 44.70 ± 7.96 c | 0.42 ± 0.13 | 54.81 ± 5.19 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, Z.-Y.; Liu, T.; Qin, C.-J.; Zou, Y.-C.; Wang, J.; Li, R.; Tao, Y.-X. MRAP2 Interaction with Melanocortin-4 Receptor in SnakeHead (Channa argus). Biomolecules 2021, 11, 481. https://doi.org/10.3390/biom11030481
Wen Z-Y, Liu T, Qin C-J, Zou Y-C, Wang J, Li R, Tao Y-X. MRAP2 Interaction with Melanocortin-4 Receptor in SnakeHead (Channa argus). Biomolecules. 2021; 11(3):481. https://doi.org/10.3390/biom11030481
Chicago/Turabian StyleWen, Zheng-Yong, Ting Liu, Chuan-Jie Qin, Yuan-Chao Zou, Jun Wang, Rui Li, and Ya-Xiong Tao. 2021. "MRAP2 Interaction with Melanocortin-4 Receptor in SnakeHead (Channa argus)" Biomolecules 11, no. 3: 481. https://doi.org/10.3390/biom11030481