
Optimization of Exopolysaccharides Production by Porphyridium sordidum and Their Potential to Induce Defense Responses in Arabidopsis thaliana against Fusarium oxysporum

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain Isolation and Growth Conditions
2.2. Genomic DNA Extraction and Molecular Identification of the Isolated Strain
2.3. Extraction of Exopolysaccharides (EPS) from P. sordidum
2.4. Experimental Design and Data Analysis
2.5. Structural and Physic-Chemical Characterizations of EPS
2.5.1. HPAEC-PAD Analysis of EPS
2.5.2. HPSEC-MALLS Analysis of EPS
2.6. Plant and Fungal Growth Conditions
2.7. Elicitor Treatment and Infestation
2.8. Determination of Hydrogen Peroxide (H2O2) Content
2.9. Extraction and Measurement of Phenylalanine Ammonia Lyase (PAL) Activity
2.10. Extraction of RNA, Synthesis of cDNA and Real-Time Quantitative PCR
3. Results
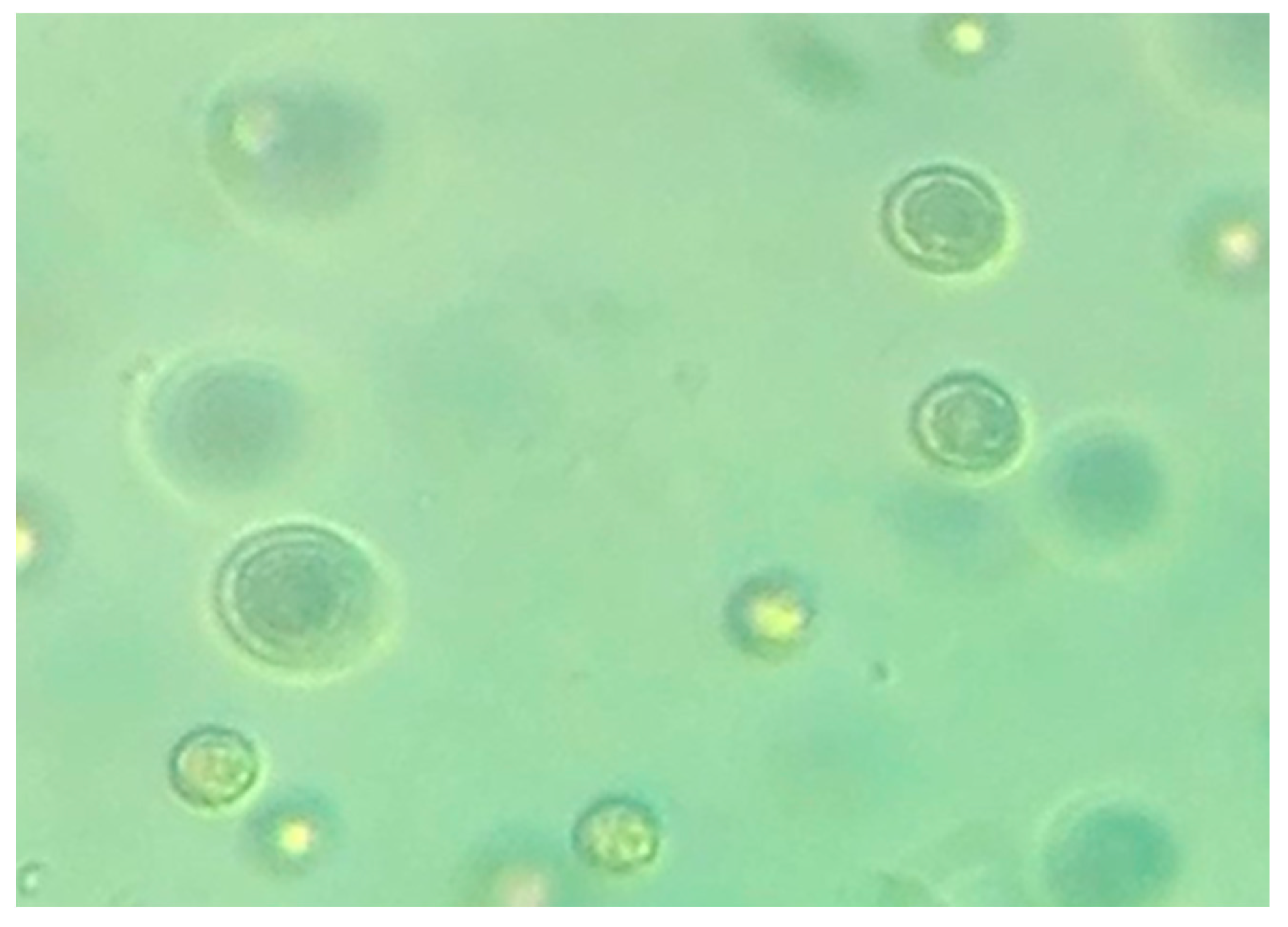
3.1. Isolation and Identification of the New Porphyridium Strain
3.2. Kinetics of Growth and EPS Production of P. sordidum under Standard Conditions
3.3. Structural Characterization of EPS
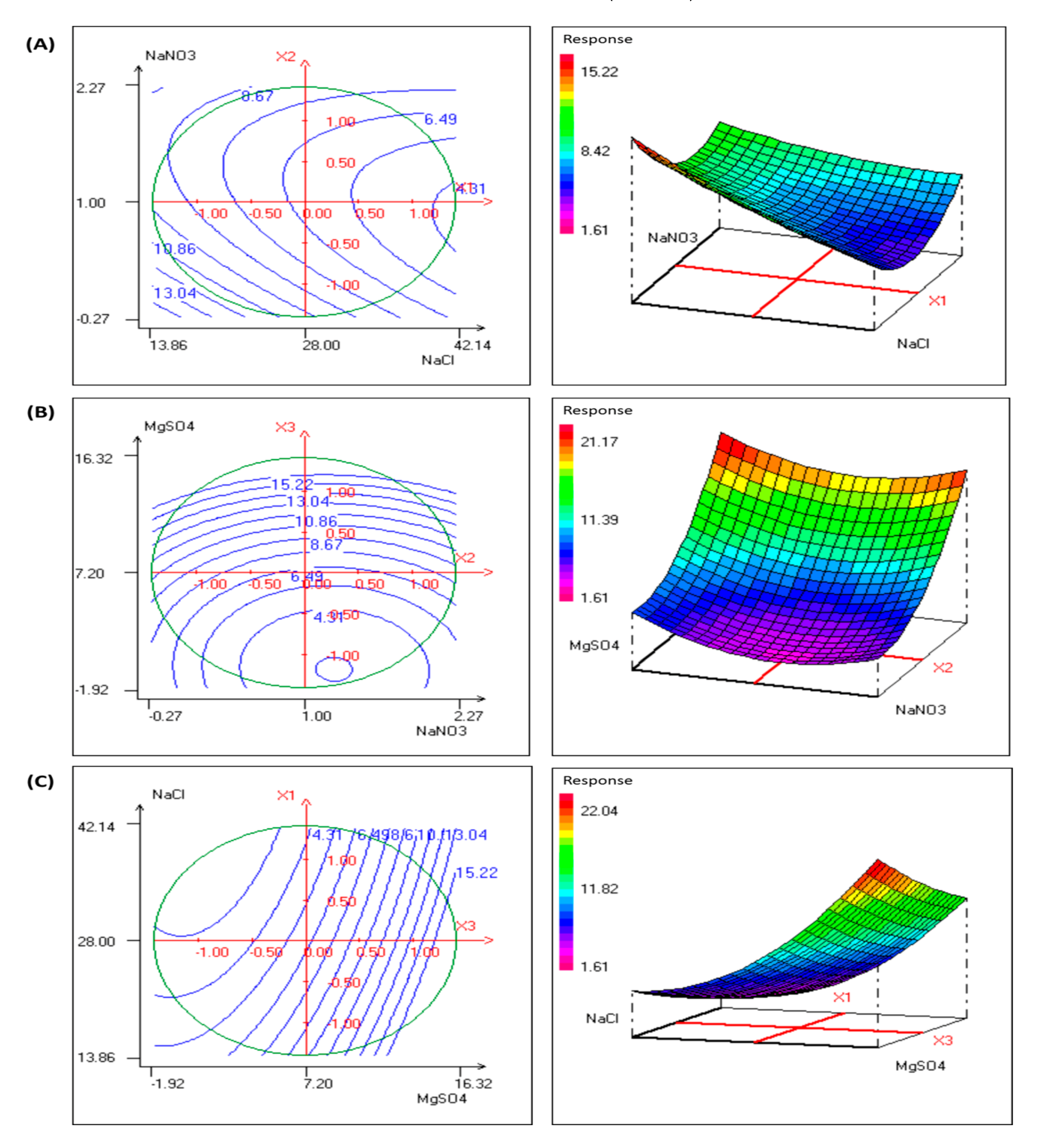
3.4. Optimization of EPS Production by P. sordidum
3.5. Effect of EPS Pretreatments on Subsequent F. oxysporum Infection in A. thaliana Plants
3.6. Effect of EPS Treatment on H2O2 Accumulation and PAL Activity
3.7. Effect of EPS Pretreatment on the Induction of Plant Defense Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hamuel, J.D. An overview of plant immunity. J. Plant Pathol. Microbiol. 2015, 6. [Google Scholar] [CrossRef]
- Wiesel, L.; Newton, A.C.; Elliott, I.; Booty, D.; Gilroy, E.M.; Birch, P.R.J.; Hein, I. Molecular effects of resistance elicitors from biological origin and their potential for crop protection. Front. Plant Sci. 2014, 5, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, A.K.; Sharma, K.; Misra, R.S. Elicitor recognition, signal transduction and induced resistance in plants. J. Plant Interact. 2012, 7, 95–120. [Google Scholar] [CrossRef]
- García-Mier, L.; Guevara-González, R.G.; Mondragón-Olguín, V.M.; Verduzco-Cuellar, B.D.R.; Torres-Pacheco, I. Agriculture and bioactives: Achieving both crop yield and phytochemicals. Int. J. Mol. Sci. 2013, 14, 4203–4222. [Google Scholar] [CrossRef] [PubMed]
- Stadnik, M.J.; de Freitas, M.B. Algal polysaccharides as source of plant resistance inducers. Trop. Plant Pathol. 2014, 39, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Vera, J.; Castro, J.; Gonzalez, A.; Moenne, A. Seaweed polysaccharides and derived oligosaccharides stimulate defense responses and protection against pathogens in plants. Mar. Drugs 2011, 9, 2514–2525. [Google Scholar] [CrossRef]
- Aitouguinane, M.; Bouissil, S.; Mouhoub, A.; Rchid, H.; Fendri, I.; Abdelkafi, S.; Ould El-Hadj, M.D.; Boual, Z.; Dubessay, P.; Gardarin, C.; et al. Induction of natural defenses in tomato seedlings by using alginate and oligoalginates derivatives extracted from Moroccan brown algae. Mar. Drugs 2020, 18, 521. [Google Scholar] [CrossRef]
- Ghannam, A.; Abbas, A.; Alek, H.; Al-Waari, Z.; Al-Ktaifani, M. Enhancement of local plant immunity against tobacco mosaic virus infection after treatment with sulphated-carrageenan from red alga (Hypnea musciformis). Physiol. Mol. Plant Pathol. 2013, 84, 19–27. [Google Scholar] [CrossRef]
- Abouraïcha, E.; El Alaoui-Talibi, Z.; El Boutachfaiti, R.; Petit, E.; Courtois, B.; Courtois, J.; El Modafar, C. Induction of natural defense and protection against Penicillium expansum and Botrytis cinerea in apple fruit in response to bioelicitors isolated from green algae. Sci. Hortic. 2015, 181, 121–128. [Google Scholar] [CrossRef]
- Chanda, M.J.; Merghoub, N.; Arroussi, H.E. Microalgae polysaccharides: The new sustainable bioactive products for the development of plant bio-stimulants? World J. Microbiol. Biotechnol. 2019, 35, 1–10. [Google Scholar] [CrossRef]
- Ben Hlima, H.; Dammak, M.; Karkouch, N.; Hentati, F.; Laroche, C.; Michaud, P.; Fendri, I.; Abdelkafi, S. Optimal cultivation towards enhanced biomass and floridean starch production by Porphyridium marinum. Int. J. Biol. Macromol. 2019, 129, 152–161. [Google Scholar] [CrossRef]
- Ben Amor, F.; Elleuch, F.; Ben Hlima, H.; Garnier, M.; Saint-Jean, B.; Barkallah, M.; Pichon, C.; Abdelkafi, S.; Fendri, I. Proteomic analysis of the chlorophyta Dunaliella new strain AL-1 revealed global changes of metabolism during high carotenoid production. Mar. Drugs 2017, 15, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dammak, M.; Haase, S.M.; Miladi, R.; Ben Amor, F.; Barkallah, M.; Gosset, D.; Pichon, C.; Huchzermeyer, B.; Fendri, I.; Denis, M.; et al. Enhanced lipid and biomass production by a newly isolated and identified marine microalga. Lipids Health Dis. 2016, 15, 209. [Google Scholar] [CrossRef] [Green Version]
- Abdelkafi, S.; Chamkha, M.; Casalot, L.; Sayadi, S.; Labat, M. Isolation and characterization of a novel Bacillus sp., strain YAS1, capable of transforming tyrosol under hypersaline conditions. FEMS Microbiol. Lett. 2005, 252, 79–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fendri, I.; Chaari, A.; Dhouib, A.; Jlassi, B.; Abousalham, A.; Carriere, F.; Sayadi, S.; Abdelkafi, S. Isolation, identification and characterization of a new lipolytic Pseudomonas sp., strain AHD-1, from Tunisian soil. Environ. Technol. 2010, 31, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Hentati, F.; Delattre, C.; Gardarin, C.; Desbrières, J.; Le Cerf, D.; Rihouey, C.; Michaud, P.; Abdelkafi, S.; Pierre, G. Structural features and rheological properties of a sulphated xylogalactan-rich fraction isolated from Tunisian red seaweed Jania ahaerens. Appl. Sci. 2020, 10, 1655. [Google Scholar] [CrossRef] [Green Version]
- Hentati, F.; Delattre, C.; Ursu, A.V.; Desbrières, J.; Le Cerf, D.; Gardarin, C.; Abdelkafi, S.; Michaud, P.; Pierre, G. Structural characterization and antioxidant activity of water-soluble polysaccharides from the Tunisian brown seaweed Cystoseira compressa. Carbohydr. Polym. 2018, 198, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Planatrum 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- El-Shora, H.M. Properties of phenylalanine ammonia-lyase from marrow cotyledons. Plant Sci. 2002, 162, 1–7. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ott, F.D. A second record of Porphyridium sordidum Geitler 1. J. Phycol. 1967, 3, 34–36. [Google Scholar] [CrossRef]
- Medina-Cabrera, E.V.; Rühmann, B.; Schmid, J.; Sieber, V. Characterization and comparison of Porphyridium Sordidum and Porphyridium Purpureum concerning growth characteristics and polysaccharide production. Algal Res. 2020, 49, 101931. [Google Scholar] [CrossRef]
- Kroen, W.K.; Rayburn, W.R. Influence of growth status and nutrients on extracellular polysaccharide synthesis by the soil alga Chlamydomonas mexicana (Chlorophyceae). J. Phycol. 1984, 20, 253–257. [Google Scholar] [CrossRef]
- Díaz Bayona, K.C.; Garcés, L.A. Effect of different media on exopolysaccharide and biomass production by the green microalga Botryococcus Braunii. J. Appl. Phycol. 2014, 26, 2087–2095. [Google Scholar] [CrossRef]
- Patel, A.K.; Laroche, C.; Marcati, A.; Ursu, A.V.; Jubeau, S.; Marchal, L.; Petit, E.; Djelveh, G.; Michaud, P. Separation and fractionation of exopolysaccharides from Porphyridium cruentum. Bioresour. Technol. 2013, 145, 345–350. [Google Scholar] [CrossRef]
- Gaignard, C.; Gargouch, N.; Dubessay, P.; Delattre, C.; Pierre, G.; Laroche, C.; Fendri, I.; Abdelkafi, S.; Michaud, P. New horizons in culture and valorization of red microalgae. Biotechnol. Adv. 2019, 37, 193–222. [Google Scholar] [CrossRef]
- Han, S.I.I.; Jeon, M.S.; Heo, Y.M.; Kim, S.; Choi, Y.E. Effect of Pseudoalteromonas sp. MEBiC 03485 on biomass production and sulfated polysaccharide biosynthesis in Porphyridium cruentum UTEX 161. Bioresour. Technol. 2020, 302, 122791. [Google Scholar] [CrossRef]
- Arad, S.; Levy-Ontman, O. Red microalgal cell-wall polysaccharides: Biotechnological aspects. Curr. Opin. Biotechnol. 2010, 21, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Delattre, C.; Pierre, G.; Laroche, C.; Michaud, P. Production, extraction and characterization of microalgal and cyanobacterial exopolysaccharides. Biotechnol. Adv. 2016, 34, 1159–1179. [Google Scholar] [CrossRef] [PubMed]
- Arad, S.; Lerental, Y.B.; Dubinsky, O. Effect of nitrate and sulfate starvation on polysaccharide formation in Rhodella reticulata. Bioresour. Technol. 1992, 42, 141–148. [Google Scholar] [CrossRef]
- Michielse, C.B.; Rep, M. Pathogen profile update: Fusarium oxysporum. Mol. Plant Pathol. 2009, 10, 311. [Google Scholar] [CrossRef]
- Torres, M.A.; Jones, J.D.G.; Dangl, J.L. Reactive oxygen species signaling in response to pathogens. Plant Physiol. 2006, 141, 373–378. [Google Scholar] [CrossRef] [Green Version]
- Kong, J.Q. Phenylalanine ammonia-lyase, a key component used for phenylpropanoids production by metabolic engineering. RSC Adv. 2015, 5, 62587–62603. [Google Scholar] [CrossRef]
- Zheng, F.; Chen, L.; Zhang, P.; Zhou, J.; Lu, X.; Tian, W. Carbohydrate polymers exhibit great potential as effective elicitors in organic agriculture: A review. Carbohydr. Polym. 2020, 230, 115637. [Google Scholar] [CrossRef]
- Barkallah, M.; Ben Atitallah, A.; Hentati, F.; Dammak, M.; Hadrich, B.; Fendri, I.; Ayadi, M.-A.; Michaud, P.; Abdelkafi, S. Effect of Spirulina platensis biomass with high polysaccharides content on quality attributes of common carp (Cyprinus carpio) and common barbel (Barbus barbus) fish burgers. Appl. Sci. 2019, 9, 2197. [Google Scholar] [CrossRef] [Green Version]
- Cruz, D.; Vasconcelos, V.; Pierre, G.; Michaud, P.; Delattre, C. Exopolysaccharides from cyanobacteria: Strategies for bioprocess development. Appl. Sci. 2020, 10, 3763. [Google Scholar] [CrossRef]
- Ozturk, S.; Aslim, B. Modification of exopolysaccharide composition and production by three cyanobacterial isolates under salt stress. Environ. Sci. Pollut. Res. 2010, 17, 595–602. [Google Scholar] [CrossRef]
- Jindal, N.; Singh, D.P.; Khattar, J.I.S. Kinetics and physico-chemical characterization of exopolysaccharides produced by the cyanobacterium Oscillatoria formosa. World J. Microbiol. Biotechnol. 2011, 27, 2139–2146. [Google Scholar] [CrossRef]
- Borah, D.; Nainamalai, S.; Gopalakrishnan, S.; Rout, J.; Alharbi, N.S.; Alharbi, S.A.; Nooruddin, T. Biolubricant potential of exopolysaccharides from the cyanobacterium Cyanothece epiphytica. Appl. Microbiol. Biotechnol. 2018, 102, 3635–3647. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Jha, B. Isolation and characterization of extracellular polymeric substances from micro-algae Dunaliella salina under salt stress. Bioresour. Technol. 2009, 100, 3382–3386. [Google Scholar] [CrossRef]
- Li, T.; Xu, J.; Wu, H.; Jiang, P.; Chen, Z.; Xiang, W. Growth and biochemical composition of Porphyridium purpureum SCS-02 under different nitrogen concentrations. Mar. Drugs 2019, 17, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, M.B.F.; Azero, E.G.; Teixeira, C.M.L.L.; Andrade, C.T. Influence of culture conditions on the production of Extracellular Polymeric Substances (EPS) by Arthrospira Platensis. Bioresour. Bioprocess. 2020, 7. [Google Scholar] [CrossRef]
- Cepák, V.; Přibyl, P. Light intensity and nitrogen effectively control exopolysaccharide production by the green microalga botryococcus braunii (trebouxiophyceae). Genet. Plant Physiol. 2018, 8, 24–37. [Google Scholar]
- Aziz, A.; Poinssot, B.; Daire, X.; Adrian, M.; Bézier, A.; Lambert, B.; Joubert, J.M.; Pugin, A. Laminarin elicits defense responses in grapevine and induces protection against Botrytis cinerea and Plasmopara viticola. Mol. Plant Microbe Interact. 2003, 16, 1118–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, P.; Ramanujam, R.; Venkatesan, G.; Nagarathnam, R. Sodium alginate potentiates antioxidant defense and PR proteins against early blight disease caused by Alternaria solani in Solanum lycopersicum Linn. PLoS ONE 2019, 14, e223216. [Google Scholar] [CrossRef] [Green Version]
- Sangha, J.S.; Ravichandran, S.; Prithiviraj, K.; Critchley, A.T.; Prithiviraj, B. Sulfated macroalgal polysaccharides λ-carrageenan and ι-carrageenan differentially alter Arabidopsis thaliana resistance to Sclerotinia sclerotiorum. Physiol. Mol. Plant Pathol. 2010, 75, 38–45. [Google Scholar] [CrossRef]
- Ceron-Garcia, A.; Vargas-Arispuro, I.; Aispuro-Hernandez, E.; Angel, M. Oligoglucan elicitor effects during plant oxidative stress. Cell Metab. 2012, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Gunupuru, L.R.; Patel, J.S.; Sumarah, M.W.; Renaud, J.B.; Mantin, E.G.; Prithiviraj, B. A plant biostimulant made from the marine brown algae Ascophyllum nodosum and chitosan reduce fusarium head blight and mycotoxin contamination in wheat. PLoS ONE 2019, e220562. [Google Scholar] [CrossRef] [PubMed]
- Jayaraj, J.; Wan, A.; Rahman, M.; Punja, Z.K. Seaweed extract reduces foliar fungal diseases on carrot. Crop Prot. 2008, 27, 1360–1366. [Google Scholar] [CrossRef]
- Rachidi, F.; Chanda, M.J.; Benhima, R.; El Mernissi, N.; Asfar, A.; Sbabou, L.; El Arroussi, H. Effect of microalgae polysaccharides on biochemical and metabolomics pathways related to plant defense in Solanum lycopersicum. Appl. Biochem. Biotechnol. 2019, 188, 225–240. [Google Scholar] [CrossRef]
- Trouvelot, S.; Héloir, M.C.; Poinssot, B.; Gauthier, A.; Paris, F.; Guillier, C.; Combier, M.; Trdá, L.; Daire, X.; Adrian, M. Carbohydrates in plant immunity and plant protection: Roles and potential application as foliar sprays. Front. Plant Sci. 2014, 5, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Rachidi, F.; Benhima, R.; Kasmi, Y.; Sbabou, L.; Arroussi, H.E. Evaluation of microalgae polysaccharides as biostimulants of tomato plant defense using metabolomics and biochemical approaches. Sci. Rep. 2021, 1–16. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Symbol | Levels | ||
|---|---|---|---|---|
| −1 | 0 | +1 | ||
| NaCl (g/L) | X1 | 18 | 28 | 38 |
| NaNO₃ (g/L) | X2 | 0.1 | 1 | 1.9 |
| MgSO₄ (g/L) | X3 | 0.75 | 7.2 | 13.68 |
| Gene | Description | Primer (F = forward; R = reverse) |
|---|---|---|
| SOD | Superoxide dismutase | F 5′-CGCATGATCCTTTGGCTTCG-3′ R 5′-TCCTGGTTGGCTGTGGTTTC-3′ |
| CAT | Catalase | F 5′-TCCTGTTATCGTTCGTTTCTCA-3′ R 5′-CAAAGTTCCCCTCTCTGGTGTA-3′ |
| POD | Peroxidase | F 5′-CCAAACTCTTCGTGGACTATGC-3′ R 5′-AACTCTTGGTCGCTCTGGAT-3′ |
| PR1 | Pathogenesis-related protein 1 | F 5’-ACGGGGAAAACTTAGCCTGG-3’ R 5’-GCCTTCTCGCTAACCCACAT-3’ |
| PDF1.2 | Plant defensin 1.2 | F 5’- TGCTGGGAAGACATAGTTGC-3’ R 5’- TGGTGGAAGCACAGAAGTTG-3’ |
| CYP | Cytochrome P450 monooxygenase | F 5’-GCTGGACCAAATGGGGATCA-3’ R 5’-TCGGCAAACATCGAGACCAA-3’ |
| ACT | Actin | F 5’-CATCAGGAAGGACTTGTACGG-3’ R 5’-GATGGACCTGACTCGTCATAC-3’ |
| GAPC2 | Glyceraldehyde-3-phosphate dehydrogenase C2 | F 5’-AGAATTGGACGTTTGGTTG-3′ R 5’-ACTTTGACAGGCTTCTCAC-3′ |
| Monosaccharides | Fuc | Rha | Ara | Gal | Glc | Xyl | GlcA |
|---|---|---|---|---|---|---|---|
| Molar% | 1.93 | 0.36 | 0.36 | 48.28 | 19.01 | 28.2 | 0.76 |
| SulfateContent (w/w) | Macromolecular Parameters Determined by HPSEC-MALLS | ||||||
|---|---|---|---|---|---|---|---|
| 6.7% | Mn (g/mol) | Mw (g/mol) | Đ | Rg (nm) | Rh (nm) | [η] (mL/g) | C* (g/L) |
| 12 × 10⁵ | 14 × 10⁵ | 1.1 | 120 | 66 | 1360 | 2.94 | |
| Runing Order | Factors | EPS Concentration (g/L) | |||
|---|---|---|---|---|---|
| X1 | X2 | X3 | Y exp | Y pre | |
| 1 | −1 | −1 | 0 | 0.93 | 0.95 |
| 2 | 1 | −1 | 0 | 2.42 | 2.23 |
| 3 | −1 | 1 | 0 | 0.7 | 0.89 |
| 4 | 1 | 1 | 0 | 1.42 | 1.40 |
| 5 | −1 | 0 | −1 | 0.88 | 0.73 |
| 6 | 1 | 0 | −1 | 1.71 | 1.77 |
| 7 | −1 | 0 | 1 | 0.73 | 0.67 |
| 8 | 1 | 0 | 1 | 1.26 | 1.41 |
| 9 | 0 | −1 | −1 | 0.94 | 1.07 |
| 10 | 0 | 1 | −1 | 0.84 | 0.80 |
| 11 | 0 | −1 | 1 | 0.99 | 1.03 |
| 12 | 0 | 1 | 1 | 0.54 | 0.41 |
| 13 | 0 | 0 | 0 | 0.76 | 0.76 |
| 14 | 0 | 0 | 0 | 0.74 | 0.76 |
| 15 | 0 | 0 | 0 | 0.77 | 0.76 |
| Source of Variation | Sum of Squares | Degrees of Freedom | Mean Square | Ratio | Significance * |
|---|---|---|---|---|---|
| Regression | 0.31753 | 9 | 0.3528 | 1512.0706 | Significant |
| Residual | 0.1639 | 5 | 0.0328 | ||
| Lack of fit | 0.1639 | 3 | 0.0545 | 233.4643 | Not significant |
| Error | 0.0005 | 2 | 0.0002 | ||
| Total | 3.3392 | 14 |
| Coefficients | Coefficient Values | Student Test | p-Value |
|---|---|---|---|
| b0 | 0.757 | 85.80 | <0.0001 |
| b1 | 0.446 | 82.63 | <0.0001 |
| b2 | −0.222 | −41.20 | <0.0001 |
| b3 | −0.106 | −19.67 | <0.001 |
| b11 | 0.464 | 58.39 | <0.0001 |
| b22 | 0.147 | 18.45 | <0.001 |
| b33 | −0.076 | −9.54 | <0.001 |
| b12 | −0.192 | −25.20 | <0.0001 |
| b13 | −0.075 | −9.82 | <0.001 |
| b23 | −0.087 | −11.46 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drira, M.; Elleuch, J.; Ben Hlima, H.; Hentati, F.; Gardarin, C.; Rihouey, C.; Le Cerf, D.; Michaud, P.; Abdelkafi, S.; Fendri, I. Optimization of Exopolysaccharides Production by Porphyridium sordidum and Their Potential to Induce Defense Responses in Arabidopsis thaliana against Fusarium oxysporum. Biomolecules 2021, 11, 282. https://doi.org/10.3390/biom11020282
Drira M, Elleuch J, Ben Hlima H, Hentati F, Gardarin C, Rihouey C, Le Cerf D, Michaud P, Abdelkafi S, Fendri I. Optimization of Exopolysaccharides Production by Porphyridium sordidum and Their Potential to Induce Defense Responses in Arabidopsis thaliana against Fusarium oxysporum. Biomolecules. 2021; 11(2):282. https://doi.org/10.3390/biom11020282
Chicago/Turabian StyleDrira, Marwa, Jihen Elleuch, Hajer Ben Hlima, Faiez Hentati, Christine Gardarin, Christophe Rihouey, Didier Le Cerf, Philippe Michaud, Slim Abdelkafi, and Imen Fendri. 2021. "Optimization of Exopolysaccharides Production by Porphyridium sordidum and Their Potential to Induce Defense Responses in Arabidopsis thaliana against Fusarium oxysporum" Biomolecules 11, no. 2: 282. https://doi.org/10.3390/biom11020282