
Molecular and Genetic Mechanisms That Mediate Transmission of Yersinia pestis by Fleas

Abstract
:1. Introduction
2. The Insect Host Environment
3. Regurgitative Transmission of Y. pestis by Fleas
4. Y. pestis Transmission Factors
4.1. Biomolecules Required to Survive and Grow in the Flea Digestive Tract: Resistance to the Flea Antibacterial Response
4.2. Biomolecules Required to Survive and Grow in the Flea Digestive Tract: The Bacterial Autoaggregation Phenotype
4.3. Biomolecules Required to Survive and Grow in the Flea Digestive Tract: The Response to Nutrient Limitation and Other Stresses
4.4. Biomolecules Required for Mature Biofilm Formation in the Flea Digestive Tract
4.5. Biomolecules Left behind during the Evolution to Flea-Borne Transmission
5. Future Directions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Achtman, M.; Morelli, G.; Zhu, P.; Wirth, T.; Diehl, I.; Kusecek, B.; Vogler, A.J.; Wagner, D.M.; Allender, C.J.; Easterday, W.R.; et al. Microevolution and history of the plague bacillus, Yersinia pestis. Proc. Natl. Acad. Sci. USA 2004, 101, 17837–17842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Y.; Yu, C.; Yan, Y.; Li, D.; Li, Y.; Jombart, T.; Weinert, L.A.; Wang, Z.; Guo, Z.; Xu, L.; et al. Historical variations in mutation rate in an epidemic pathogen, Yersinia pestis. Proc. Natl. Acad. Sci. USA 2013, 110, 577–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, S.; Allentoft, M.E.; Nielsen, K.; Orlando, L.; Sikora, M.; Sjogren, K.G.; Pedersen, A.G.; Schubert, M.; Van Dam, A.; Kapel, C.M.; et al. Early divergent strains of Yersinia pestis in Eurasia 5000 years ago. Cell 2015, 163, 571–582. [Google Scholar] [CrossRef] [PubMed]
- Lorange, E.A.; Race, B.L.; Sebbane, F.; Hinnebusch, B.J. Poor vector competence of fleas and the evolution of hypervirulence in Yersinia pestis. J. Inf. Dis. 2005, 191, 1907–1912. [Google Scholar] [CrossRef] [Green Version]
- Vaschenok, V.S.; Solina, L.T.; Zhirnov, A.E. The characteristics of the digestion of the blood of various animals by the flea Xenopsylla cheopis. Parasitology 1976, 10, 544–549. (In Russian) [Google Scholar]
- Vaughan, J.A.; Azad, A.F. Patterns of erythrocyte digestion by bloodsucking insects: Constraints on vector competence. J. Med. Entomol. 1993, 30, 214–216. [Google Scholar] [CrossRef]
- Silverman, J.; Appel, A.G. Adult cat flea (Siphonaptera: Pulicidae) excretion of host blood proteins in relation to larval nutrition. J. Med. Entomol. 1994, 31, 265–271. [Google Scholar] [CrossRef]
- Bontemps-Gallo, S.; Fernandez, M.; Dewitte, A.; Raphael, E.; Gherardini, F.C.; Elizabeth, P.; Koch, L.; Biot, F.; Reboul, A.; Sebbane, F. Nutrient depletion may trigger the Yersinia pestis OmpR-EnvZ regulatory system to promote flea-borne plague transmission. Mol. Microbiol. 2019, 112, 1471–1482. [Google Scholar] [CrossRef]
- Wigglesworth, V.B. The regulation of respiration in the flea, Xenopsylla cheopis, Roths. (Pulicidae). Proc. R. Soc. Ser. B Biol. 1935, 118, 397–419. [Google Scholar]
- Bland, D.M.; Martens, C.A.; Virtaneva, K.; Kanakabandi, K.; Long, D.; Rosenke, R.; Saturday, G.A.; Hoyt, F.H.; Bruno, D.P.; Ribeiro, J.M.; et al. Transcriptomic profiling of the digestive tract of the rat flea, Xenopsylla cheopis, following blood feeding and infection with Yersinia pestis. PLoS Negl. Trop. Dis. 2020, 14, e0008688. [Google Scholar] [CrossRef]
- Capo, F.; Charroux, B.; Royet, J. Bacteria sensing mechanisms in Drosophila gut: Local and systemic consequences. Dev. Comp. Immunol. 2016, 64, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.D. Immunity of fleas (Order Siphonaptera). Dev. Comp. Immunol. 2019, 98, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Russell, C.W.; Johnson, K.L.; Mortensen, R.D.; Erickson, D.L. Gene expression analysis of Xenopsylla cheopis (Siphonaptera: Pulicidae) suggests a role for reactive oxygen species in response to Yersinia pestis infection. J. Med. Entomol. 2012, 49, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Graca-Souza, A.V.; Maya-Monteiro, C.; Paiva-Silva, G.O.; Braz, G.R.; Paes, M.C.; Sorgine, M.H.; Oliveira, M.F.; Oliveira, P.L. Adaptations against heme toxicity in blood-feeding arthropods. Insect Biochem. Mol. Biol. 2006, 36, 322–335. [Google Scholar] [CrossRef]
- Toh, S.Q.; Glanfield, A.; Gobert, G.N.; Jones, M.K. Heme and blood-feeding parasites: Friends or foes? Parasit. Vectors 2010, 3, 108. [Google Scholar] [CrossRef] [Green Version]
- Driscoll, T.P.; Verhoeve, V.I.; Gillespie, J.J.; Johnston, J.S.; Guillotte, M.L.; Rennoll-Bankert, K.E.; Rahman, M.S.; Hagen, D.; Elsik, C.G.; Macaluso, K.R.; et al. A chromosome-level assembly of the cat flea genome uncovers rampant gene duplication and genome size plasticity. BMC Biol. 2020, 18, 70. [Google Scholar] [CrossRef]
- Hinnebusch, B.J.; Jarrett, C.O.; Bland, D.M. “Fleaing” the plague: Adaptations of Yersinia pestis to its insect vector that lead to transmission. Annu. Rev. Microbiol. 2017, 71, 215–232. [Google Scholar] [CrossRef]
- Bland, D.M.; Jarrett, C.O.; Bosio, C.F.; Hinnebusch, B.J. Infectious blood source alters early foregut infection and regurgitative transmission of Yersinia pestis by rodent fleas. PLoS Pathog. 2018, 14, e1006859. [Google Scholar] [CrossRef] [Green Version]
- Dewitte, A.; Bouvenot, T.; Pierre, F.; Ricard, I.; Pradel, E.; Barois, N.; Hujeux, A.; Bontemps-Gallo, S.; Sebbane, F. A refined model of how Yersinia pestis produces a transmissible infection in its flea vector. PLoS Pathog. 2020, 16, e1008440. [Google Scholar] [CrossRef] [Green Version]
- Eisen, R.J.; Bearden, S.W.; Wilder, A.P.; Montenieri, J.A.; Antolin, M.F.; Gage, K.L. Early-phase transmission of Yersinia pestis by unblocked fleas as a mechanism explaining rapidly spreading plague epizootics. Proc. Natl. Acad. Sci. USA 2006, 103, 15380–15385. [Google Scholar] [CrossRef] [Green Version]
- Bosio, C.F.; Jarrett, C.O.; Scott, D.P.; Fintzi, J.; Hinnebusch, B.J. Comparison of the transmission efficiency and plague progression dynamics associated with two mechanisms by which fleas transmit Yersinia pestis. PLoS Pathog. 2020, 16, e1009092. [Google Scholar] [CrossRef] [PubMed]
- Bacot, A.W.; Martin, C.J. Observations on the mechanism of the transmission of plague by fleas. J. Hyg. Plague 1914, 13 (Suppl. 3), 423–439. [Google Scholar]
- Motin, V.L.; Georgescu, A.M.; Fitch, J.P.; Gu, P.P.; Nelson, D.O.; Mabery, S.L.; Garnham, J.B.; Sokhansanj, B.A.; Ott, L.L.; Coleman, M.A.; et al. Temporal global changes in gene expression during temperature transition in Yersinia pestis. J. Bacteriol. 2004, 186, 6298–6305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Zhou, D.; Pang, X.; Song, Y.; Zhang, L.; Bao, J.; Tong, Z.; Wang, J.; Guo, Z.; Zhai, J.; et al. Microarray analysis of temperature-induced transcriptome of Yersinia pestis. Microbiol. Immunol. 2004, 48, 791–805. [Google Scholar] [CrossRef] [PubMed]
- Hinnebusch, B.J.; Sebbane, F.; Vadyvaloo, V. Transcriptional profiling of the Yersinia pestis life cycle. In Yersinia: Systems Biology and Control; Carniel, E., Hinnebusch, B.J., Eds.; Horizon Scientific Press: Norwich, UK, 2012. [Google Scholar]
- Sebbane, F.; Lemaitre, N.; Sturdevant, D.E.; Rebeil, R.; Virtaneva, K.; Porcella, S.F.; Hinnebusch, B.J. Adaptive response of Yersinia pestis to extracellular effectors of innate immunity during bubonic plague. Proc. Natl. Acad. Sci. USA 2006, 103, 11766–11771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vadyvaloo, V.; Jarrett, C.; Sturdevant, D.E.; Sebbane, F.; Hinnebusch, B.J. Transit through the flea vector induces a pretransmission innate immunity resistance phenotype in Yersinia pestis. PLoS Pathog. 2010, 6, e10000783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vadyvaloo, V.; Viall, A.K.; Jarrett, C.O.; Hinz, A.K.; Sturdevant, D.E.; Hinnebusch, B.J. Role of the PhoP-PhoQ gene regulatory system in adaptation of Yersinia pestis to environmental stress in the flea digestive tract. Microbiology 2015, 161, 1198–1210. [Google Scholar] [CrossRef]
- Chouikha, I.; Sturdevant, D.E.; Jarrett, C.; Sun, Y.C.; Hinnebusch, B.J. Differential gene expression patterns of Yersinia pestis and Yersinia pseudotuberculosis during infection and biofilm formation in the flea digestive tract. mSystems 2019, 4, e00217-18. [Google Scholar] [CrossRef] [Green Version]
- Hinnebusch, B.J.; Rudolph, A.E.; Cherepanov, P.; Dixon, J.E.; Schwan, T.G.; Forsberg, Å. Role of Yersinia murine toxin in survival of Yersinia pestis in the midgut of the flea vector. Science 2002, 296, 733–735. [Google Scholar] [CrossRef]
- Vadyvaloo, V.; Hinz, A.K. A LysR-type transcriptional regulator, RovM, senses nutritional cues suggesting that it is involved in metabolic adaptation of Yersinia pestis to the flea gut. PLoS ONE 2015, 10, e0137508. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Fang, H.; Yang, H.; Zhang, Y.; Han, Y.; Zhou, D.; Yang, R. Reciprocal regulation of Yersinia pestis biofilm formation and virulence by RovM and RovA. Open Biol. 2016, 6, 150198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erickson, D.L.; Russell, C.W.; Johnson, K.L.; Hileman, T.; Stewart, R.M. PhoP and OxyR transcriptional regulators contribute to Yersinia pestis virulence and survival within Galleria mellonella. Microb. Pathog. 2011, 51, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Earl, S.C.; Rogers, M.T.; Keen, J.; Bland, D.M.; Houppert, A.S.; Miller, C.; Temple, I.; Anderson, D.M.; Marketon, M.M. Resistance to innate immunity contributes to colonization of the insect gut by Yersinia pestis. PLoS ONE 2015, 10, e0133318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoyagi, K.L.; Brooks, B.D.; Bearden, S.W.; Montenieri, J.A.; Gage, K.L.; Fisher, M.A. LPS modification promotes maintenance of Yersinia pestis in fleas. Microbiology 2015, 161, 628–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bobrov, A.G.; Kirillina, O.; Forman, S.; Mack, D.; Perry, R.D. Insights into Yersinia pestis biofilm development: Topology and co-interaction of Hms inner membrane proteins involved in exopolysaccharide production. Environ. Microbiol. 2008, 10, 1419–1432. [Google Scholar] [CrossRef]
- Hinnebusch, B.J.; Perry, R.D.; Schwan, T.G. Role of the Yersinia pestis hemin storage (hms) locus in the transmission of plague by fleas. Science 1996, 273, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Jarrett, C.O.; Deak, E.; Isherwood, K.E.; Oyston, P.C.; Fischer, E.R.; Whitney, A.R.; Kobayashi, S.D.; DeLeo, F.R.; Hinnebusch, B.J. Transmission of Yersinia pestis from an infectious biofilm in the flea vector. J. Inf. Dis. 2004, 190, 783–792. [Google Scholar] [CrossRef] [Green Version]
- Bobrov, A.G.; Kirillina, O.; Ryjenkov, D.A.; Waters, C.M.; Price, P.A.; Fetherston, J.D.; Mack, D.; Goldman, W.E.; Gomelsky, M.; Perry, R.D. Systematic analysis of cyclic di-GMP signalling enzymes and their role in biofilm formation and virulence in Yersinia pestis. Mol. Microbiol. 2011, 79, 533–551. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.-C.; Koumoutsi, A.; Jarrett, C.; Lawrence, K.; Gherardini, F.C.; Darby, C.; Hinnebusch, B.J. Differential control of Yersinia pestis biofilm formation in vitro and in the flea vector by two c-di-GMP diguanylate cyclases. PLoS ONE 2011, 6, e19267. [Google Scholar] [CrossRef] [Green Version]
- Ren, G.X.; Yan, H.Q.; Zhu, H.; Guo, X.P.; Sun, Y.C. HmsC, a periplasmic protein, controls biofilm formation via repression of HmsD, a diguanylate cyclase in Yersinia pestis. Environ. Microbiol. 2014, 16, 1202–1216. [Google Scholar] [CrossRef]
- Bobrov, A.G.; Kirillina, O.; Vadyvaloo, V.; Koestler, B.J.; Hinz, A.K.; Mack, D.; Waters, C.M.; Perry, R.D. The Yersinia pestis HmsCDE regulatory system is essential for blockage of the oriental rat flea (Xenopsylla cheopis), a classic plague vector. Environ. Microbiol. 2015, 17, 947–959. [Google Scholar] [CrossRef] [Green Version]
- Ren, G.X.; Fan, S.; Guo, X.P.; Chen, S.; Sun, Y.C. Differential regulation of c-di-GMP metabolic enzymes by environmental signals modulates biofilm formation in Yersinia pestis. Front. Microbiol. 2016, 7, 821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirillina, O.; Fetherston, J.D.; Bobrov, A.G.; Abney, J.; Perry, R.D. HmsP, a putative phosphodiesterase, and HmsT, a putative diguanylate cyclase, control Hms-dependent biofilm formation in Yersinia pestis. Mol. Microbiol. 2004, 54, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Rebeil, R.; Jarrett, C.O.; Driver, J.D.; Ernst, R.K.; Oyston, P.C.; Hinnebusch, B.J. Induction of the Yersinia pestis PhoP-PhoQ regulatory system in the flea and its role in producing a transmissible infection. J. Bacteriol. 2013, 195, 1920–1930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.-C.; Koumoutsi, A.; Darby, C. The response regulator PhoP negatively regulates Yersinia peseudotuberculosis and Yersinia pestis biofilms. FEMS Microbiol. Lett. 2009, 290, 85–90. [Google Scholar] [CrossRef]
- Hinnebusch, B.J.; Jarrett, C.O. NIH, NIAID Rocky Mountain Laboratories, Hamilton, MT, USA. Unpublished work. Unpublished work. 2017. [Google Scholar]
- Willias, S.P.; Chauhan, S.; Lo, C.C.; Chain, P.S.; Motin, V.L. CRP-mediated carbon catabolite regulation of Yersinia pestis biofilm formation is enhanced by the carbon storage regulator protein, CsrA. PLoS ONE 2015, 10, e0135481. [Google Scholar] [CrossRef]
- Liu, L.; Fang, H.; Yang, H.; Zhang, Y.; Han, Y.; Zhou, D.; Yang, R. CRP Is an activator of Yersinia pestis biofilm formation that operates via a mechanism involving gmhA and waaAE-coaD. Front. Microbiol. 2016, 7, 295. [Google Scholar] [CrossRef] [Green Version]
- Tam, C.; Demke, O.; Hermanas, T.; Mitchell, A.; Hendrickx, A.P.; Schneewind, O. YfbA, a Yersinia pestis regulator required for colonization and biofilm formation in the gut of cat fleas. J. Bacteriol. 2014, 196, 1165–1173. [Google Scholar] [CrossRef] [Green Version]
- Darby, C.; Ananth, S.L.; Tan, L.; Hinnebusch, B.J. Identification of gmhA, a Yersinia pestis gene required for flea blockage, by using a Caenorhabditis elegans biofilm system. Infect. Immun. 2005, 73, 7236–7242. [Google Scholar] [CrossRef] [Green Version]
- Rempe, K.A.; Hinz, A.K.; Vadyvaloo, V. Hfq regulates biofilm gut blockage that facilitates flea-borne transmission of Yersinia pestis. J. Bacteriol. 2012, 194, 2036–2040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellows, L.E.; Koestler, B.J.; Karaba, S.M.; Waters, C.M.; Lathem, W.W. Hfq-dependent, co-ordinate control of cyclic diguanylate synthesis and catabolism in the plague pathogen Yersinia pestis. Mol. Microbiol. 2012, 86, 661–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Gao, X.; Wang, H.; Fang, H.; Yan, Y.; Liu, L.; Chen, R.; Zhou, D.; Yang, R.; Han, Y. Plasmid pPCP1-derived sRNA HmsA promotes biofilm formation of Yersinia pestis. BMC Microbiol. 2016, 16, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, N.; Qu, S.; Yang, H.; Fang, H.; Liu, L.; Zhang, Y.; Wang, L.; Han, Y.; Zhou, D.; Yang, R. HmsB enhances biofilm formation in Yersinia pestis. Front. Microbiol. 2014, 5, 685. [Google Scholar] [CrossRef] [Green Version]
- Tan, L.; Darby, C. Yersinia pestis YrbH is a multifunctional protein required for both 3-deoxy-D-manno-oct-2-ulosonic acid biosynthesis and biofilm formation. Mol. Microbiol. 2006, 61, 861–870. [Google Scholar] [CrossRef]
- Fang, H.; Liu, L.; Zhang, Y.; Yang, H.; Yan, Y.; Ding, X.; Han, Y.; Zhou, D.; Yang, R. BfvR, an AraC-family regulator, controls biofilm formation and pH6 antigen production in opposite ways in Yersinia pestis Biovar Microtus. Front. Cell. Infect. Microbiol. 2018, 8, 347. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Schmid, B.V.; Cao, H.; Dai, X.; Du, Z.; Ryan Easterday, W.; Fang, H.; Guo, C.; Huang, S.; Liu, W.; et al. Evolutionary selection of biofilm-mediated extended phenotypes in Yersinia pestis in response to a fluctuating environment. Nat. Commun. 2020, 11, 281. [Google Scholar] [CrossRef] [Green Version]
- Gage, K.L.; Kosoy, M.Y. Natural history of plague: Perspectives from more than a century of research. Annu. Rev. Entomol. 2005, 50, 505–528. [Google Scholar] [CrossRef] [Green Version]
- Chouikha, I.; Hinnebusch, B.J. Silencing urease: A key evolutionary step that facilitated the adaptation of Yersinia pestis to the flea-borne transmission route. Proc. Natl. Acad. Sci. USA 2014, 111, 18709–18714. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.C.; Jarrett, C.O.; Bosio, C.F.; Hinnebusch, B.J. Retracing the evolutionary path that led to flea-borne transmission of Yersinia pestis. Cell Host Microbe 2014, 15, 578–586. [Google Scholar] [CrossRef] [Green Version]
- Fang, N.; Yang, H.; Fang, H.; Liu, L.; Zhang, Y.; Wang, L.; Han, Y.; Zhou, D.; Yang, R. RcsAB is a major repressor of Yersinia biofilm development through directly acting on hmsCDE, hmsT, and hmsHFRS. Sci. Rep. 2015, 5, 9566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.C.; Guo, X.P.; Hinnebusch, B.J.; Darby, C. The Yersinia pestis Rcs phosphorelay inhibits biofilm formation by repressing transcription of the diguanylate cyclase gene hmsT. J. Bacteriol. 2012, 194, 2020–2026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erickson, D.L.; Jarrett, C.O.; Callison, J.A.; Fischer, E.R.; Hinnebusch, B.J. Loss of a biofilm-inhibiting glycosyl hydrolase during the emergence of Yersinia pestis. J. Bacteriol. 2008, 190, 8163–8170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chain, P.S.G.; Carniel, E.; Larimer, F.W.; Lamerdin, J.; Stoutland, P.O.; Regala, W.M.; Georgescu, A.M.; Vergez, L.M.; Land, M.L.; Motin, V.L.; et al. Insights into the evolution of Yersinia pestis through whole-genome comparison with Yersinia pseudotuberculosis. Proc. Natl. Acad. Sci. USA 2004, 101, 13826–13831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trunk, T.; Khalil, H.S.; Leo, J.C. Bacterial autoaggregation. AIMS Microbiol. 2018, 4, 140–164. [Google Scholar] [CrossRef]
- Chauhan, N.; Wrobel, A.; Skurnik, M.; Leo, J.C. Yersinia adhesins: An arsenal for infection. Proteom. Clin. Appl. 2016, 10, 949–963. [Google Scholar] [CrossRef] [Green Version]
- Calder, J.T.; Christman, N.D.; Hawkins, J.M.; Erickson, D.L. A trimeric autotransporter enhances biofilm cohesiveness in Yersinia pseudotuberculosis but not in Yersinia pestis. J. Bacteriol. 2020, 202. [Google Scholar] [CrossRef]
- Hatkoff, M.; Runco, L.M.; Pujol, C.; Jayatilaka, I.; Furie, M.B.; Bliska, J.B.; Thanassi, D.G. Roles of chaperone/usher pathways of Yersinia pestis in a murine model of plague and adhesion to host cells. Infect. Immun. 2012, 80, 3490–3500. [Google Scholar] [CrossRef] [Green Version]
- Kolodziejek, A.M.; Hovde, C.J.; Minnich, S.A. Yersinia pestis Ail: Multiple roles of a single protein. Front. Cell. Infect. Microbiol. 2012, 2. [Google Scholar] [CrossRef] [Green Version]
- Bartra, S.S.; Styer, K.L.; O’Bryant, D.M.; Nilles, M.L.; Hinnebusch, B.J.; Aballay, A.; Plano, G.V. Resistance of Yersinia pestis to complement-dependent killing is mediated by the Ail outer membrane protein. Infect. Immun. 2008, 76, 612–622. [Google Scholar] [CrossRef] [Green Version]
- Eboigbodin, K.E.; Newton, J.R.; Routh, A.F.; Biggs, C.A. Role of nonadsorbing polymers in bacterial aggregation. Langmuir 2005, 21, 12315–12319. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, P.; Mondal, J.; Ben-Jacob, E.; Levine, H. Mechanically-driven phase separation in a growing bacterial colony. Proc. Natl. Acad. Sci. USA 2015, 112, E2166–E2173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Chavarria, L.C.; Sagawa, J.; Irons, J.; Hinz, A.K.; Lemon, A.; Graca, T.; Downs, D.M.; Vadyvaloo, V. Putative horizontally acquired genes, highly transcribed during Yersinia pestis flea infection, are induced by hyperosmotic stress and function in aromatic amino acid metabolism. J. Bacteriol. 2020, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelthaler, D.M.; Hinnebusch, B.J.; Rittner, C.M.; Gage, K.L. Quantitative competitive PCR as a technique for exploring flea-Yersina pestis dynamics. Am. J. Trop. Med. Hyg. 2000, 62, 552–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinnebusch, B.J.; Bland, D.M.; Bosio, C.F.; Jarrett, C.O. Comparative ability of Oropsylla montana and Xenopsylla cheopis fleas to transmit Yersinia pestis by two different mechanisms. PLoS Negl. Trop. Dis. 2017, 11, e0005276. [Google Scholar] [CrossRef] [PubMed]
- Hengge, R. Linking bacterial growth, survival, and multicellularity—Small signaling molecules as triggers and drivers. Curr. Opin. Microbiol. 2020, 55, 57–66. [Google Scholar] [CrossRef]
- Liu, L.; Zheng, S. Transcriptional regulation of Yersinia pestis biofilm formation. Microb. Pathog. 2019, 131, 212–217. [Google Scholar] [CrossRef]
- Hengge, R. Principles of c-di-GMP signalling in bacteria. Nat. Rev. Microbiol. 2009, 7, 263–273. [Google Scholar] [CrossRef]
- Jenal, U.; Reinders, A.; Lori, C. Cyclic di-GMP: Second messenger extraordinaire. Nat. Rev. Microbiol. 2017, 15, 271–284. [Google Scholar] [CrossRef] [Green Version]
- Erickson, D.L.; Jarrett, C.O.; Wren, B.W.; Hinnebusch, B.J. Serotype differences and lack of biofilm formation characterize Yersinia pseudotuberculosis infection of the Xenopsylla cheopis flea vector of Yersinia pestis. J. Bacteriol. 2006, 188, 1113–1119. [Google Scholar] [CrossRef] [Green Version]
- Wall, E.; Majdalani, N.; Gottesman, S. The complex Rcs regulatory cascade. Annu. Rev. Microbiol. 2018, 72, 111–139. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.P.; Ren, G.X.; Zhu, H.; Mao, X.J.; Sun, Y.C. Differential regulation of the hmsCDE operon in Yersinia pestis and Yersinia pseudotuberculosis by the Rcs phosphorelay system. Sci. Rep. 2015, 5, 8412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.-C.; Hinnebusch, B.J.; Darby, C. Experimental evidence for negative selection in the evolution of a Yersinia pestis pseudogene. Proc. Natl. Acad. Sci. USA 2008, 105, 8097–8101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, R.; Song, Y.; Dai, Q.; Kang, Y.; Pan, J.; Zhu, L.; Zhang, L.; Wang, Y.; Shen, X. A starvation-induced regulator, RovM, acts as a switch for planktonic/biofilm state transition in Yersinia pseudotuberculosis. Sci. Rep. 2017, 7, 639. [Google Scholar] [CrossRef]



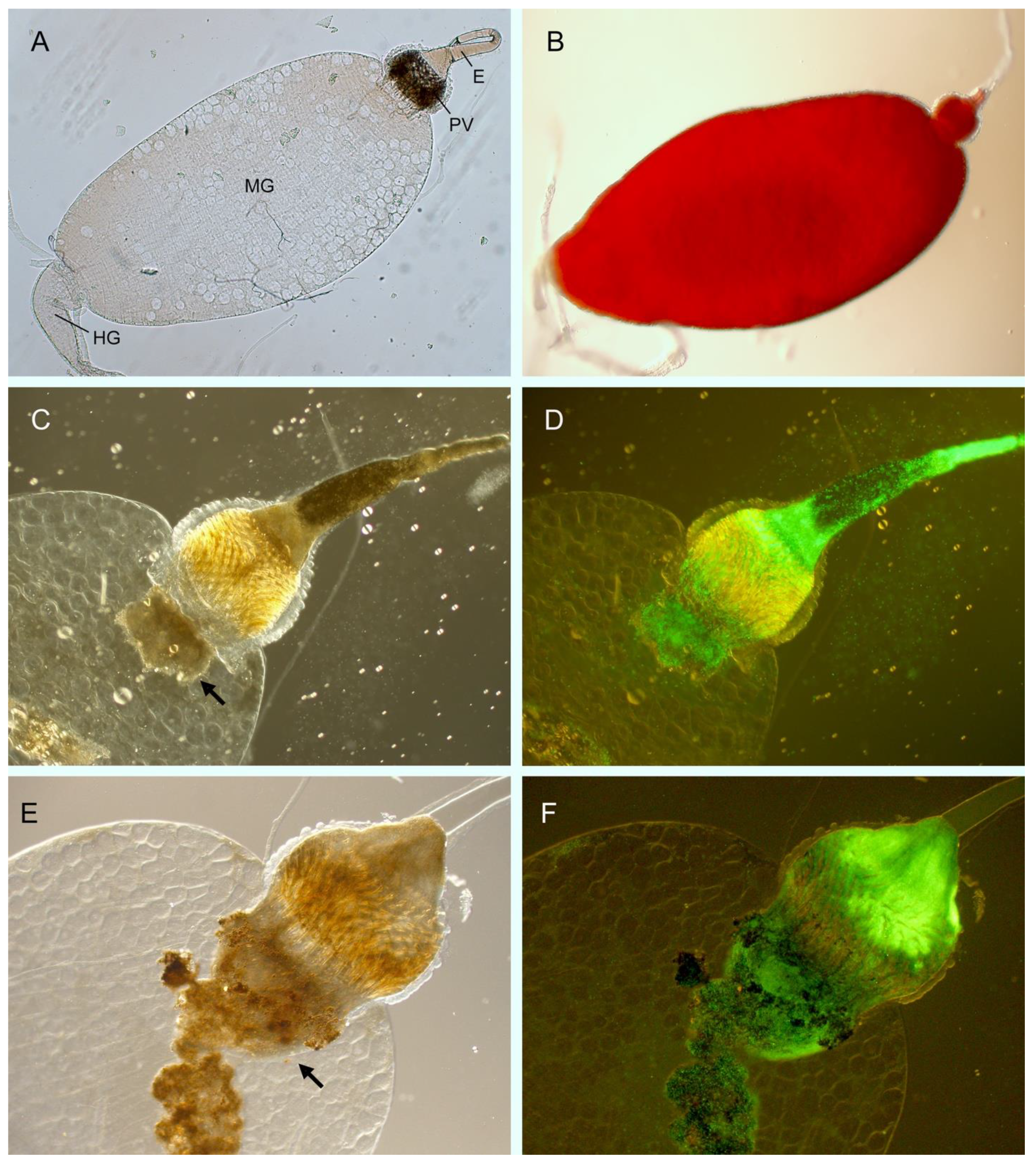{kind=link}
{kind=link}
| Name | Molecular Function | Role in the Flea | Reference |
|---|---|---|---|
| Ymt | phospholipase D | protection against toxic byproduct of blood digestion | [30] |
| RovM | transcriptional regulator | (i) metabolic adaptation to the flea gut; (ii) upregulates hmsCDE, hmsT; increases intracellular c-di-GMP and biofilm production | [31,32] |
| OxyR | transcriptional regulator | protection against reactive oxygen species generated in the flea | [13,33,34] |
| GalU, ArnB | lipid A modification enzymes | protection against antimicrobial peptides of flea immune response | [35] |
| HmsHFRS | glycolsyl transferase, polysaccharide deacetylase, transport porin | synthesis of extracellular β-1,6-GlcNAc polysaccharide biofilm matrix component | [36,37,38] |
| HmsCDE | diguanylate cyclase and its regulators | c-di-GMP synthesis | [39,40,41,42,43] |
| HmsT | diguanylate cyclase | c-di-GMP synthesis | [39,40] |
| HmsP | phosphodiesterase | c-di-GMP degradation | [44] |
| PhoP | transcriptional regulator | (i) protection against antimicrobial peptides of flea immune response; (ii) enhances stability of biofilm; mechanism unknown | [27,45,46] |
| Crp | cAMP receptor protein | (i) carbon catabolite regulation enabling metabolism of alternate carbon sources; (ii) required for biofilm production, activates gmhA and waaAE-coaD | [47,48,49] |
| YfbA | transcriptional regulator | enhances biofilm production; mechanism unknown | [50] |
| GmhA | phosphoheptose isomerase | heptose synthesis; LPS production | [51] |
| RpiA | ribose-5-phosphate isomerase | enhances stability of biofilm; mechanism unknown | [19] |
| Hfq | RNA chaperone | regulation of hmsT and hmsP | [52,53] |
| HmsA * | sRNA | upregulates hmsCDE, hmsT, hmsHFRS; increases intracellular c-di-GMP | [54] |
| HmsB * | sRNA | upregulates hmsCDE, hmsT, hmsHFRS; represses hmsP; increases intracellular c-di-GMP | [55] |
| CsrA * | carbon storage regulator protein | carbon catabolite regulation; enhances biofilm production; mechanism unknown | [48] |
| YrbH * | arabinose 5-phosphate isomerase | Kdo synthesis; LPS modification | [56] |
| WaaA * | Kdo transferase | Kdo synthesis; LPS modification | [56] |
| BfvR * | transcriptional regulator | upregulates hmsHFRS, waaAE-coaD, and hmsCDE expression | [57] |
| RpoZ * | RNA polymerase subunit | different variants affect rate of biofilm production; mechanism unknown | [58] |
| Name | Molecular Function | Original Role or Phenotype | Reference |
|---|---|---|---|
| PDE2 (Rtn) | phosphodiesterase | c-di-GMP degradation | [39,40,61] |
| PDE3 | phosphodiesterase | c-di-GMP degradation | [39,40,61] |
| RcsA | transcriptional regulator | represses hmsT, hmsCDE, hmsHFRS; upregulates hmsP | [61,62,63] |
| UreD | urease enzyme subunit | urease generates toxic amounts of ammonia in the flea gut | [60] |
| NghA | glycosyl hydrolase | degradation of Hms exopolysaccharide | [64] |
| FlhD | regulator of flagellum synthesis | motility | [65] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hinnebusch, B.J.; Jarrett, C.O.; Bland, D.M. Molecular and Genetic Mechanisms That Mediate Transmission of Yersinia pestis by Fleas. Biomolecules 2021, 11, 210. https://doi.org/10.3390/biom11020210
Hinnebusch BJ, Jarrett CO, Bland DM. Molecular and Genetic Mechanisms That Mediate Transmission of Yersinia pestis by Fleas. Biomolecules. 2021; 11(2):210. https://doi.org/10.3390/biom11020210
Chicago/Turabian StyleHinnebusch, B. Joseph, Clayton O. Jarrett, and David M. Bland. 2021. "Molecular and Genetic Mechanisms That Mediate Transmission of Yersinia pestis by Fleas" Biomolecules 11, no. 2: 210. https://doi.org/10.3390/biom11020210