Human Saliva-Mediated Hydrolysis of Eugenyl-β-D-Glucoside and Fluorescein-di-β-D-Glucoside in In Vivo and In Vitro Models


Abstract
:1. Introduction
2. Materials and Methods
2.1. A New In Vivo Rapid Method for the Detection of β-D-Glucosidase Activity in the Oral Cavity
2.2. Two Kinetics of In Vitro Hydrolysis of FbD with A. niger β-D-Glucosidase and Human Saliva
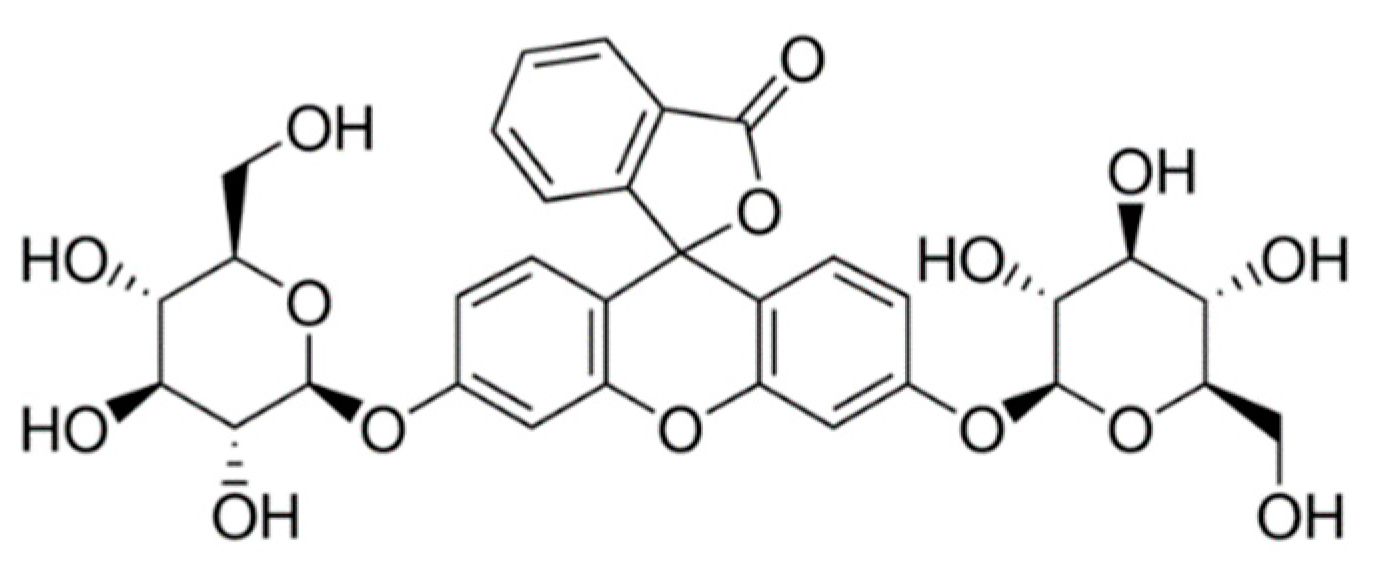
2.3. Incubation of Selected Pathogens Strains with Fluorescein-Di-β-D-Glucoside (FbD)
Microbial Species Isolation from Kefir Product and Preparation of Microbial Protein Extract for MALDI-TOF
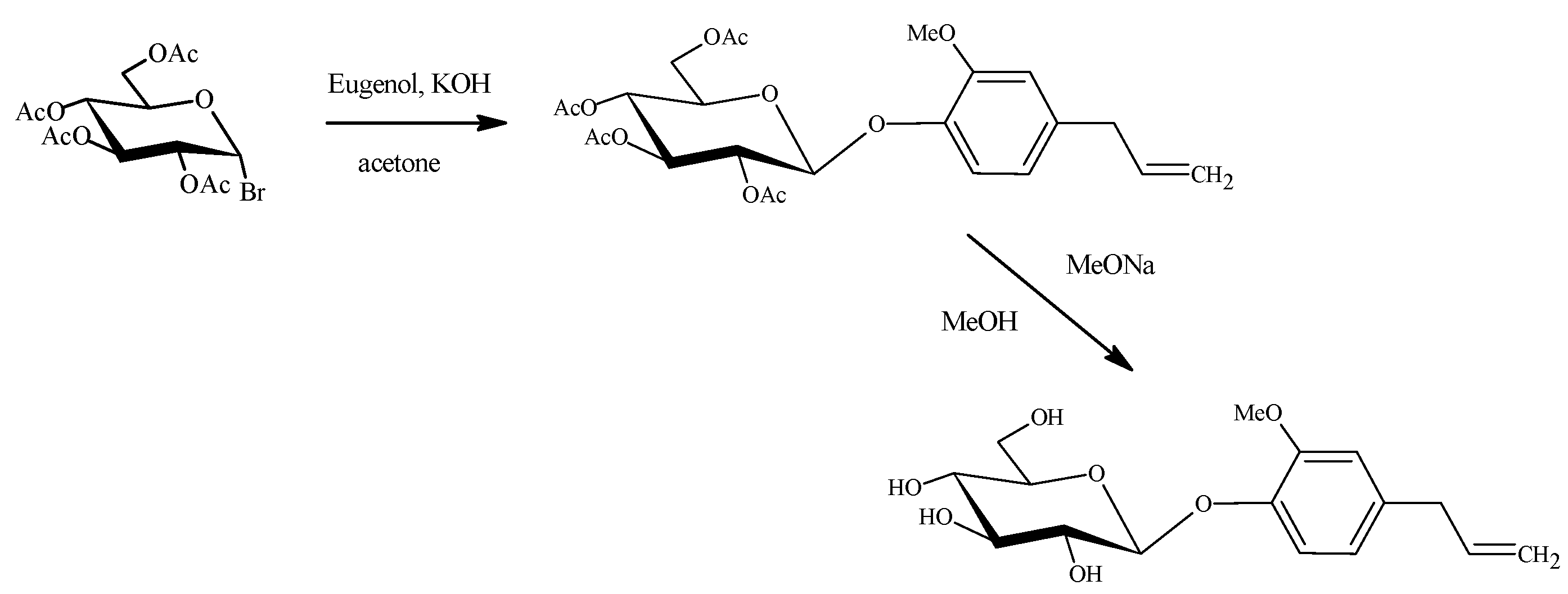
2.4. Organic Synthesis of Eugenyl β-D-Glucoside
2.5. Experiment of Hydrolysis Eugenyl β-D-Glucoside In Vitro with Antibiotics
2.5.1. Testing of Biological Activity of Substrate
2.5.2. Preparation of Reactions
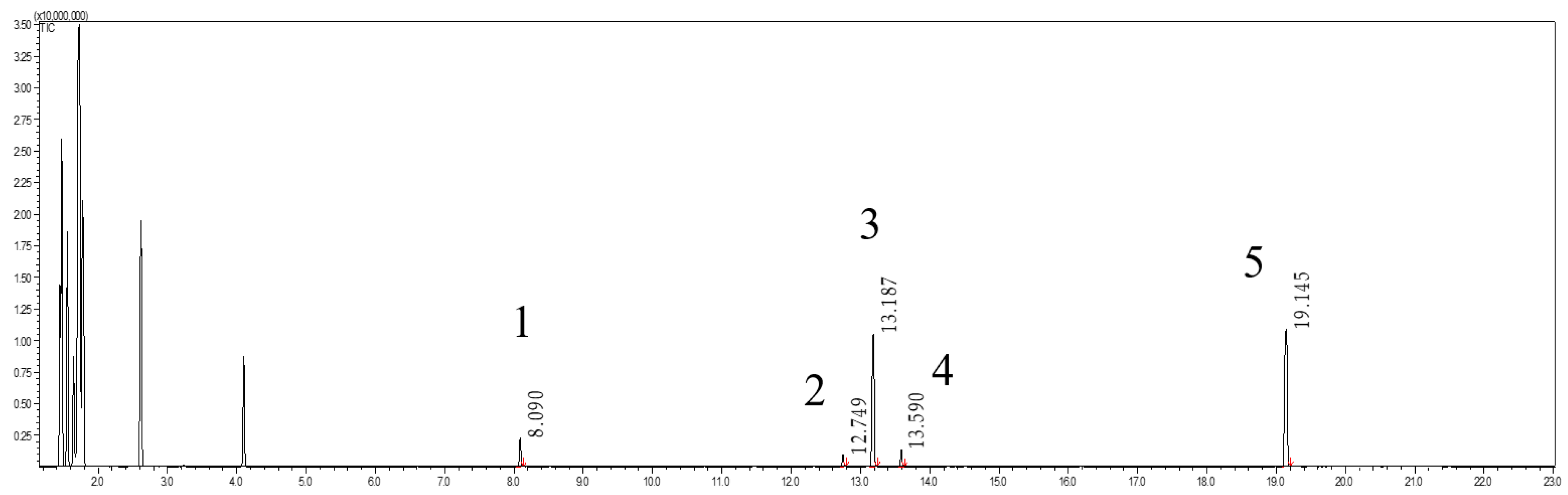
2.5.3. Sample Preparation and LC-MS/MS Analysis of Citrusin D
3. Results
3.1. A New In Vivo Rapid Method for the Detection of β-D-Glucosidase Activity in the Oral Cavity on the Tounge
3.2. Kinetics In Vitro Hydrolysis of FbD with Enzyme and Saliva
3.3. Incubation of Selected Pathogens Strains with FbD
3.4. Organic Synthesis of Eugenyl β-D-Glucoside
Synthetic Glycoside Biological Activity Test
3.5. Experiment of Hydrolysis Eugenyl β-D-Glucoside In Vitro with Human Saliva and Antibiotics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Day, A.J.; Cañada, F.J.; Díaz, J.C.; Kroon, P.A.; Mclauchlan, R.; Faulds, C.B.; Plumb, G.W.; Morgan, M.R.A.; Williamson, G. Dietary flavonoid and isoflavone glycosides are hydrolysed by the lactase site of lactase phlorizin hydrolase. FEBS Lett. 2000, 468, 166–170. [Google Scholar] [CrossRef] [Green Version]
- Hollman, P.C.H.; Van Trijp, J.M.P.; Buysman, M.N.C.P.; Gaag, M.S.; Mengelers, M.J.B.; de Vries, J.H.M.; Katan, M.B. Relative bioavailability of the antioxidant flavonoid quercetin from various foods in man. FEBS Lett. 1997, 418, 152–156. [Google Scholar] [CrossRef] [Green Version]
- King, R.A; Bursill, D.B. Plasma and urinary kinetics of the isoflavones daidzein and genistein after a single soy meal in humans. Am. J. Clin. Nutr. 1998, 67, 867–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, A.J.; DuPont, M.S.; Ridley, S.; Rhodes, M.; Rhodes, M.J.C.; Morgan, M.R.A.; Williamson, G. Deglycosylation of £avonoid and iso£avonoid glycosides by human small intestine and liver L-glucosidase activity. FEBS Lett. 1998, 436, 71–75. [Google Scholar] [CrossRef] [Green Version]
- Bokkenheuser, V.D.; Shackelton, C.H.L.; Winter, J. Hydrolysis of dietary flavonoid glycosides by strains of intestinal Bacteroides from humans. Biochem. J. 1987, 248, 953–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walle, T.; Browning, A.M.; Steed, L.L.; Reed, S.G.; Walle, U.K. Flavonoid glucosides are hydrolyzed and thus activated in the oral cavity in humans. J. Nutr. 2005, 135, 48–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthiller, F.; Krska, R.; Domig, K.J.; Kneifeld, W.; Jugee, N.; Schuhmacherb, R.; Adam, G. Hydrolytic fate of deoxynivalenol-3-glucoside during digestion. Toxicol. Lett. 2011, 206, 264–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemingway, K.M.; Alston, M.J.; Chappell, C.G.; Taylor, A.J. Carbohydrate-flavour conjugates in wine. Carbohydr. Polym. 1999, 38, 283–286. [Google Scholar] [CrossRef]
- Muńoz-González, C.; Cueva, C.; Angeles Pozo-Bayón, M.; Moreno-Arribas, V. Ability of human oral microbiota to produce wine odorant aglycones from odourless grape glycosidic aroma precursors. Food Chem. 2015, 187, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Hirota, S.; Nishioka, T.; Shimoda, T.; Miura, K.; Ansai, T.; Takahama, U. Quercetin Glucosides are Hydrolyzed to Quercetin in Human Oral Cavity to Participate in Peroxidase-Dependent Scavenging of Hydrogen Peroxide. Food Sci. Technol. Res. 2001, 7, 239–245. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Water Solutions | V1 | V2 | V3 | V4 * | V5 | V6 | V7 |
|---|---|---|---|---|---|---|---|---|
| 1 | Saliva with water (1:10, v/v) | 8 | 8 | 8 | 8 | |||
| 2 | Citrusin D (2.5 mg/mL) | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 3 | β-D-glucosidase (2 mg/mL) | 1 | 0.1 | |||||
| 4 | all antibiotics (2 mg/mL) | 1 | 1 | 1 | ||||
| 5 | sucrose (2%) | 1 | ||||||
| 6 | sterile deionized water | 9 | 1 | 7 | 9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dziadas, M.; Junka, A.; Jeleń, H. Human Saliva-Mediated Hydrolysis of Eugenyl-β-D-Glucoside and Fluorescein-di-β-D-Glucoside in In Vivo and In Vitro Models. Biomolecules 2021, 11, 172. https://doi.org/10.3390/biom11020172
Dziadas M, Junka A, Jeleń H. Human Saliva-Mediated Hydrolysis of Eugenyl-β-D-Glucoside and Fluorescein-di-β-D-Glucoside in In Vivo and In Vitro Models. Biomolecules. 2021; 11(2):172. https://doi.org/10.3390/biom11020172
Chicago/Turabian StyleDziadas, Mariusz, Adam Junka, and Henryk Jeleń. 2021. "Human Saliva-Mediated Hydrolysis of Eugenyl-β-D-Glucoside and Fluorescein-di-β-D-Glucoside in In Vivo and In Vitro Models" Biomolecules 11, no. 2: 172. https://doi.org/10.3390/biom11020172