
SNCA 3′ UTR Genetic Variants in Patients with Parkinson’s Disease



Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Control Subjects
2.2. Sample and Data Collection
2.3. Next Generation Sequencing
2.4. Statistical Analysis
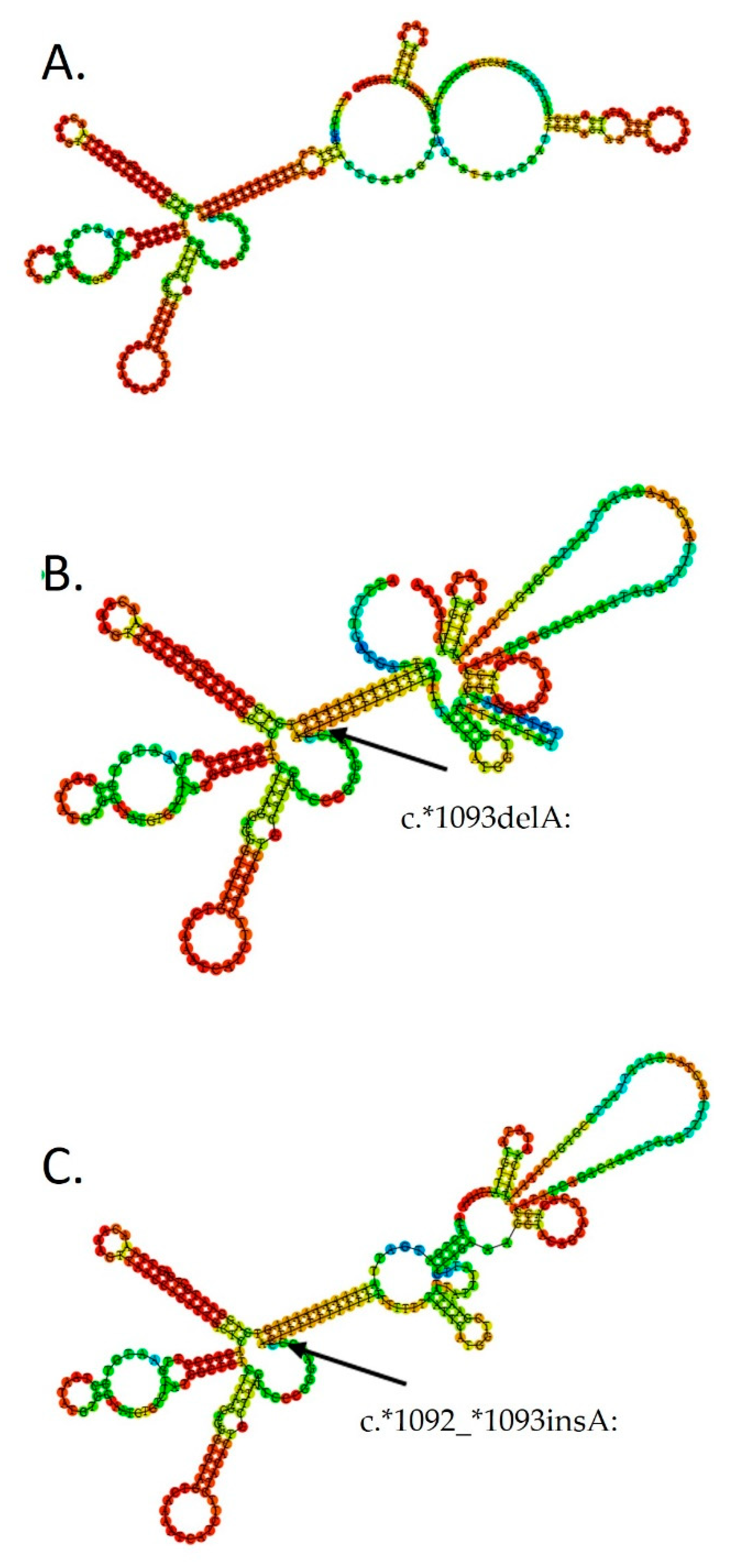
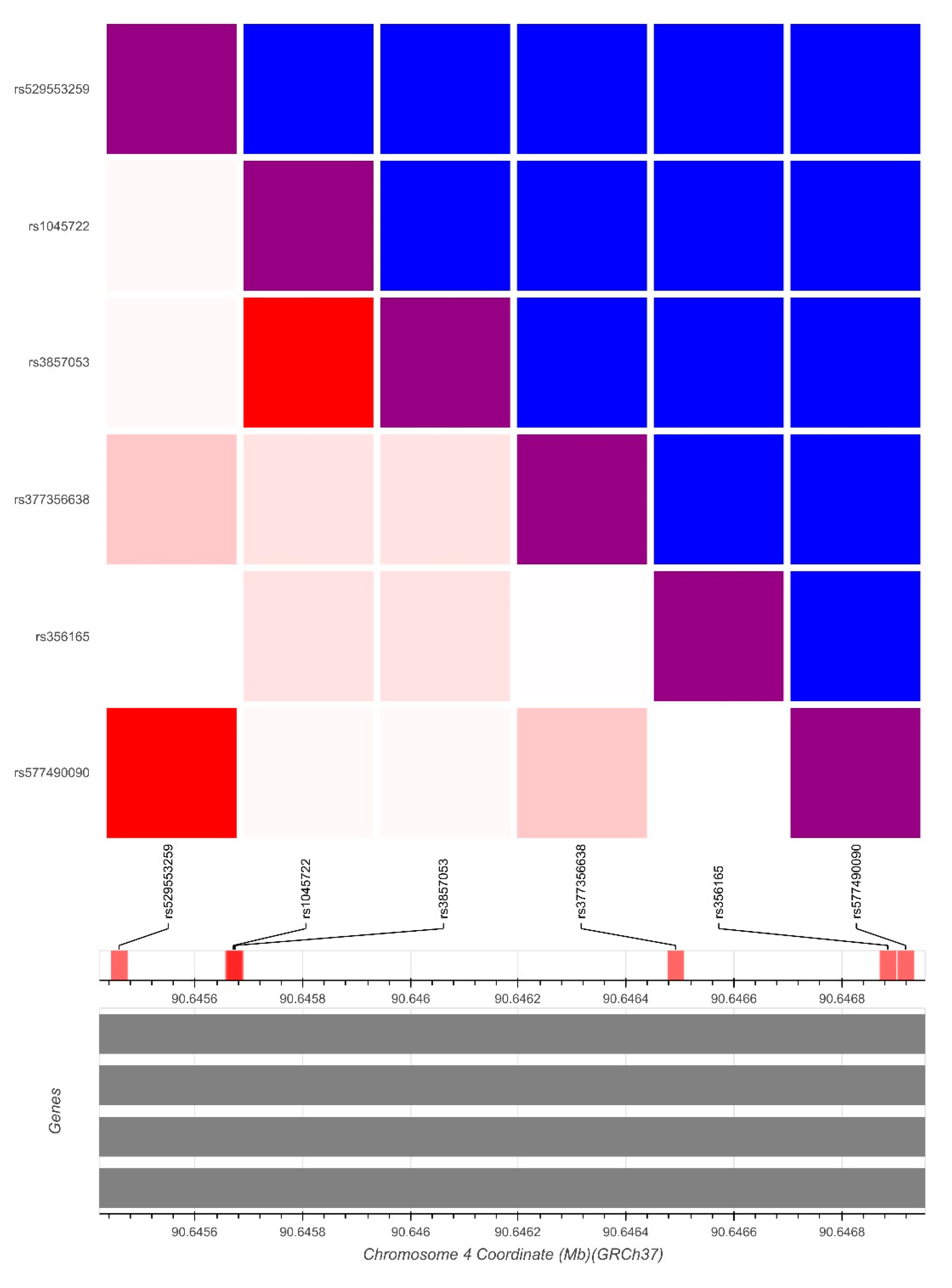
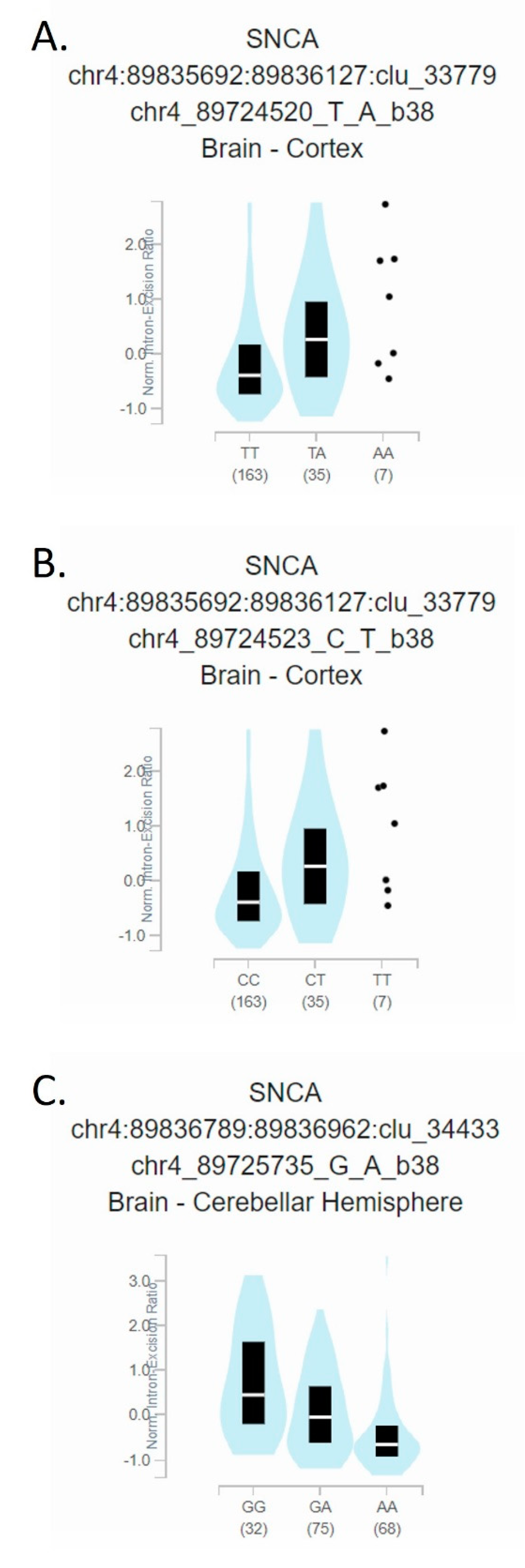
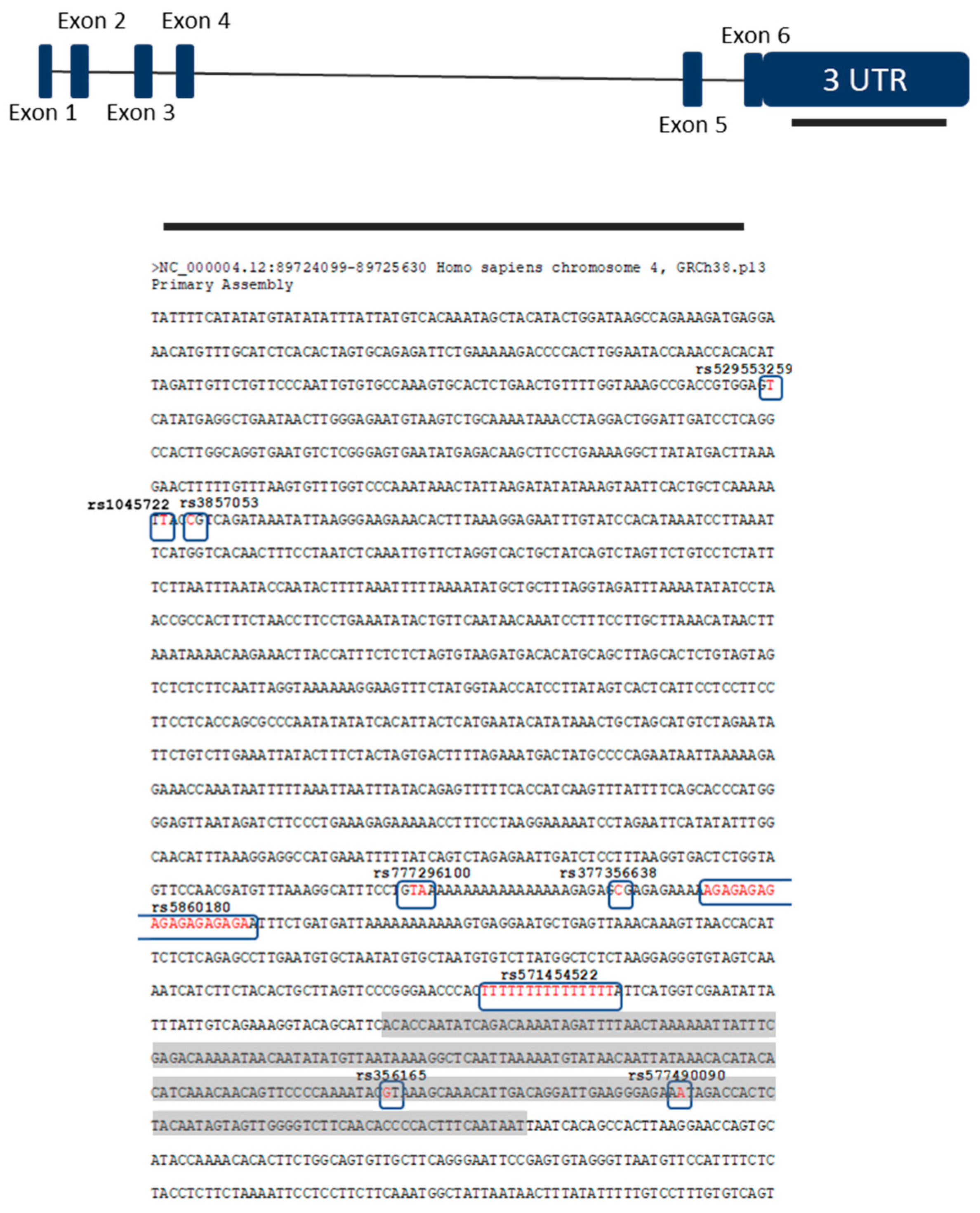
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barrett, L.W.; Fletcher, S.; Wilton, S.D. Regulation of eukaryotic gene expression by the untranslated gene regions and other non-coding elements. Cell. Mol. Life Sci. 2012, 69, 3613–3634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junn, E.; Lee, K.W.; Jeong, B.S.; Chan, T.W.; Im, J.Y.; Mouradian, M.M. Repression of alpha-synuclein expression and toxicity by microRNA-7. Proc. Natl. Acad. Sci. USA 2009, 106, 13052–13057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Mena, L.; Samaranch, L.L.; Coto, E.; Cardo, L.F.; Ribacoba, R.; Lorenzo-Betancor, O.; Pastor, P.; Wang, L.; Irigoyen, J.; Mata, I.F.; et al. Mutational screening of PARKIN identified a 3′ UTR variant (rs62637702) associated with Parkinson’s disease. J. Mol. Neurosci. 2013, 50, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Rhinn, H.; Qiang, L.; Yamashita, T.; Rhee, D.; Zolin, A.; Vanti, W.; Abeliovich, A. Alternative alpha-synuclein transcript usage as a convergent mechanism in Parkinson’s disease pathology. Nat. Commun. 2012, 3, 1084. [Google Scholar] [CrossRef]
- Simón-Sánchez, J.; Schulte, C.; Bras, J.M.; Sharma, M.; Gibbs, J.R.; Berg, D.; Paisan-Ruiz, C.; Lichtner, P.; Scholz, S.W.; Hernandez, D.G.; et al. Genome-wide association study reveals genetic risk underlying Parkinson’s disease. Nat. Genet. 2009, 41, 1308–1312. [Google Scholar] [CrossRef]
- Soldner, F.; Stelzer, Y.; Shivalila, C.S.; Abraham, B.; Latourelle, J.C.; Barrasa, M.I.; Goldmann, J.; Myers, R.H.; Young, R.A.; Jaenisch, R. Parkinson-associated risk variant in distal enhancer of α-synuclein modulates target gene expression. Nature 2016, 533, 95–99. [Google Scholar] [CrossRef] [Green Version]
- Nalls, M.A.; Pankratz, N.; Lill, C.M.; Do, C.B.; Hernandez, D.G.; Saad, M.; DeStefano, A.L.; Kara, E.; Bras, J.; Sharma, M.; et al. Large-scale meta-analysis of genome-wide association data identifies six new risk loci for Parkinson’s disease. Nat. Genet. 2014, 46, 989–993. [Google Scholar] [CrossRef] [PubMed]
- Krohn, L.; Wu, R.Y.J.; Heilbron, K.; Ruskey, J.A.; Laurent, S.B.; Blauwendraat, C.; Alam, A.; Arnulf, I.; Hu, M.T.; Dauvilliers, Y.; et al. Fine-Mapping of SNCA in Rapid Eye Movement Sleep Behavior Disorder and Overt Synucleinopathies. Ann. Neurol. 2020, 87, 584–598. [Google Scholar] [CrossRef] [PubMed]
- Rudakou, U.; Yu, E.; Krohn, L.; A Ruskey, J.; Asayesh, F.; Dauvilliers, Y.; Spiegelman, D.; Greenbaum, L.; Fahn, S.; Waters, C.H.; et al. Targeted sequencing of Parkinson’s disease loci genes highlights SYT11, FGF20 and other associations. Brain 2021, 144, 462–472. [Google Scholar] [CrossRef]
- Marchese, D.; Botta-Orfila, T.; Cirillo, D.; Rodriguez, J.A.; Livi, C.M.; Fernández-Santiago, R.; Ezquerra, M.; Martí, M.J.; Bechara, E.; Tartaglia, G.G.; et al. Discovering the 3′ UTR-mediated regulation of alpha-synuclein. Nucleic Acids Res. 2017, 45, 12888–12903. [Google Scholar] [CrossRef]
- Sotiriou, S.; Gibney, G.; Baxevanis, A.D.; Nussbaum, R.L. A single nucleotide polymorphism in the 3′UTR of the SNCA gene encoding alpha-synuclein is a new potential susceptibility locus for Parkinson disease. Neurosci Lett. 2009, 461, 196–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardo, L.F.; Coto, E.; de Mena, L.; Ribacoba, R.; Lorenzo-Betancor, O.; Pastor, P.; Samaranch, L.; Mata, I.F.; Díaz, M.; Morís, G.; et al. A search for SNCA 3′ UTR variants identified SNP rs356165 as a determinant of disease risk and onset age in Parkinson’s disease. J. Mol. Neurosci. 2012, 47, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Je, G.; Guhathakurta, S.; Yun, S.P.; Ko, H.S.; Kim, Y.S. A novel extended form of alpha-synuclein 3′UTR in the human brain. Mol. Brain 2018, 11, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miura, P.; Shenker, S.; Andreu-Agullo, C.; Westholm, J.O.; Lai, E.C. Widespread and extensive lengthening of 3′ UTRs in the mammalian brain. Genome Res. 2013, 23, 812–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouradian, M.M. MicroRNAs in Parkinson’s disease. Neurobiol. Dis. 2012, 46, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Yu, J.T.; Tan, L. Causes and consequences of microRNA dysregulation in neurodegenerative diseases. Mol. Neurobiol. 2015, 51, 1249–1262. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.J.; Daniel, S.E.; Kilford, L.; Lees, A.J. Accuracy of clinical diagnosis of idiopathic Parkinson’s disease: A clinico-pathological study of 100 cases. J. Neurol. Neurosurg. Psychiatry 1992, 55, 181–184. [Google Scholar] [CrossRef] [Green Version]
- Tatura, R.; Kraus, T.; Giese, A.; Arzberger, T.; Buchholz, M.; Höglinger, G.; Müller, U. Parkinson’s disease: SNCA-, PARK2-, and LRRK2- targeting microRNAs elevated in cingulate gyrus. Park. Relat. Disord. 2016, 33, 115–121. [Google Scholar] [CrossRef]
- Kabaria, S.; Choi, D.C.; Chaudhuri, A.D.; Mouradian, M.M.; Junn, E. Inhibition of miR-34b and miR-34c enhances α-synuclein expression in Parkinson’s disease. FEBS Lett. 2015, 589, 319–325. [Google Scholar] [CrossRef] [Green Version]
- Ghanbari, M.; Darweesh, S.K.; de Looper, H.W.; van Luijn, M.M.; Hofman, A.; Ikram, M.A.; Franco, O.; Erkeland, S.J.; Dehghan, A. Genetic Variants in MicroRNAs and Their Binding Sites Are Associated with the Risk of Parkinson Disease. Hum. Mutat. 2016, 37, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Toffoli, M.; Dreussi, E.; Cecchin, E.; Valente, M.; Sanvilli, N.; Montico, M.; Gagno, S.; Garziera, M.; Polano, M.; Savarese, M.; et al. SNCA 3′UTR genetic variants in patients with Parkinson’s disease and REM sleep behavior disorder. Neurol. Sci. 2017, 38, 1233–1240. [Google Scholar] [CrossRef]
- Emelyanov, A.K.; Andoskin, P.A.; Miliukhina, I.V.; Timofeeva, A.A.; Yakimovskii, A.F.; Senkevich, K.A.; Nikolaev, M.A.; Pchelina, S.N. SNCA alleles rs356219 and rs356165 are associated with Parkinson’s disease and increased α-synuclein gene expression in CD45+ blood cells. Cell Tissue Biol. 2016, 10, 277–283. [Google Scholar] [CrossRef]
- Hu, F.Y.; Hu, W.B.; Liu, L.; Yu, L.H.; Xi, J.; He, X.H.; Zhu, M.-R.; Liu, Z.-L.; Xu, Y.-M. Lack of replication of a previously reported association between polymorphism in the 3′UTR of the alpha-synuclein gene and Parkinson’s disease in Chinese subjects. Neurosci. Lett. 2010, 479, 31–33. [Google Scholar] [CrossRef]
- Tagliafierro, L.; Glenn, O.C.; Zamora, M.E.; Beach, T.G.; Woltjer, R.L.; Lutz, M.W.; Chiba-Falek, O. Genetic analysis of α-synuclein 3′ untranslated region and its corresponding microRNAs in relation to Parkinson’s disease compared to dementia with Lewy bodies. Alzheimers Dement. 2017, 13, 1237–1250. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.F.; Zhu, Z.Y.; Qian, S.X.; Xu, C.Y.; Wang, Y.P. miR-30b protects nigrostriatal dopaminergic neurons from MPP(+)-induced neurotoxicity via SNCA. Brain Behav. 2020, 10, e01567. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, I.; Wüllner, U.; van Rooyen, J.P.; Khazneh, H.; Becker, J.; Volk, A.; Kubisch, C.; Becker, T.; Kostic, V.S.; Klein, C.; et al. Variants in the 3′UTR of SNCA do not affect miRNA-433 binding and alpha-synuclein expression. Eur. J. Hum. Genet. 2012, 20, 1265–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Locascio, J.J.; Eberly, S.; Liao, Z.; Liu, G.; Hoesing, A.N.; Duong, K.; Trisini-Lipsanopoulos, A.; Dhima, K.; Hung, A.Y.; Flaherty, A.W. Association between α-synuclein blood transcripts and early, neuroimaging-supported Parkinson’s disease. Brain 2015, 138, 2659–2671. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PD | M | F | Control | M | F | |
|---|---|---|---|---|---|---|
| Number of subjects | 53 | 29 (54%) | 24 (46%) | 20 | 10 (50%) | 10 (50%) |
| Mean age (SD) | 66 | 63 | 70 | 63 | 64 | 63 |
| Median age ± SD | 66.33 ± 8.63 | 63.68 ± 8.07 | 69.86 ± 8.23 | 64.56 ± 9.19 | 62.43 ± 9.18 | 64.5 ± 8.37 |
| ID | PD | CTRL | p Value * | HGVS (NM_000345.3) | Position GRCh38.p13 | Alleles | Type | Allele Frequency (1000 G) | |
|---|---|---|---|---|---|---|---|---|---|
| rs1045722 | 4 | 3 | 0.6687 | c.*2108A > T | chr4:89724520 | T > A/T > G | [15] | SNP | 0.38 |
| rs3857053 | 4 | 3 | 0.6687 | c.*2105G > A | chr4:89724523 | C > T | [15] | SNP | 0.377 |
| rs777296100 | 31 | 13 | 0.8052 | c.*1309_*1310insTTT | chr4:89725318-89725319 | insTA/insTAA/ | [16] | Insertion | |
| rs35270750 (rs5860180) | 37 | 16 | 1.000 | c.*1274_*1277delCTCT | chr4:89725351-89725369 | delGAGA/ | NR | Deletion | 0.331 |
| rs356165 | 44 | 20 | 1.000 | c.*893C > T | chr4:89725735 | G > A | [15] | SNP | 0.484 |
| rs529553259 | 1 | 0 | c.*2320A > T | chr4:89724308 | T > A | NR | SNP | 0.001 | |
| rs377356638 | 1 | 0 | c.*1287G > A | chr4:89725341 | C > T | NR/LB | SNP | 0.046 | |
| rs577490090 | 1 | 0 | c.*860T > A | chr4:89725768 | A > T | NR/VUS | SNP | 0.001 | |
| rs571454522 | 36 | 9 | 0.0211 | c.*1092delA/insA | chr4:89725536-89725550 | NR | Deletion/insertion | ||
| rs571454522 (loss) | 28 | 9 | 0.318 | c.*1092delA | chr4:89725536-89725550 | delT | NR | Deletion | |
| rs750347645 (rs571454522) (gain) | 8 | 0 | 0.0974 | c.*1092_*1093insA | chr4:89725536-89725550 | NR | Insertion |
Publisher′s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blažeković, A.; Jerčić, K.G.; Borovečki, F. SNCA 3′ UTR Genetic Variants in Patients with Parkinson’s Disease. Biomolecules 2021, 11, 1799. https://doi.org/10.3390/biom11121799
Blažeković A, Jerčić KG, Borovečki F. SNCA 3′ UTR Genetic Variants in Patients with Parkinson’s Disease. Biomolecules. 2021; 11(12):1799. https://doi.org/10.3390/biom11121799
Chicago/Turabian StyleBlažeković, Antonela, Kristina Gotovac Jerčić, and Fran Borovečki. 2021. "SNCA 3′ UTR Genetic Variants in Patients with Parkinson’s Disease" Biomolecules 11, no. 12: 1799. https://doi.org/10.3390/biom11121799