
Dynamic DNA Methylation Changes in the COMT Gene Promoter Region in Response to Mental Stress and Its Modulation by Transcranial Direct Current Stimulation

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Adaptive 2-Back Paced Auditory Serial Addition Task (PASAT)
2.3. Positive and Negative Affect Schedule (PANAS)
2.4. Transcranial Direct Current Stimulation (tDCS)
2.5. Experimental Procedure
2.6. DNAm Analysis
2.7. Saliva Cortisol Concentration
2.8. Statistical Analysis
3. Results
3.1. Study Sample
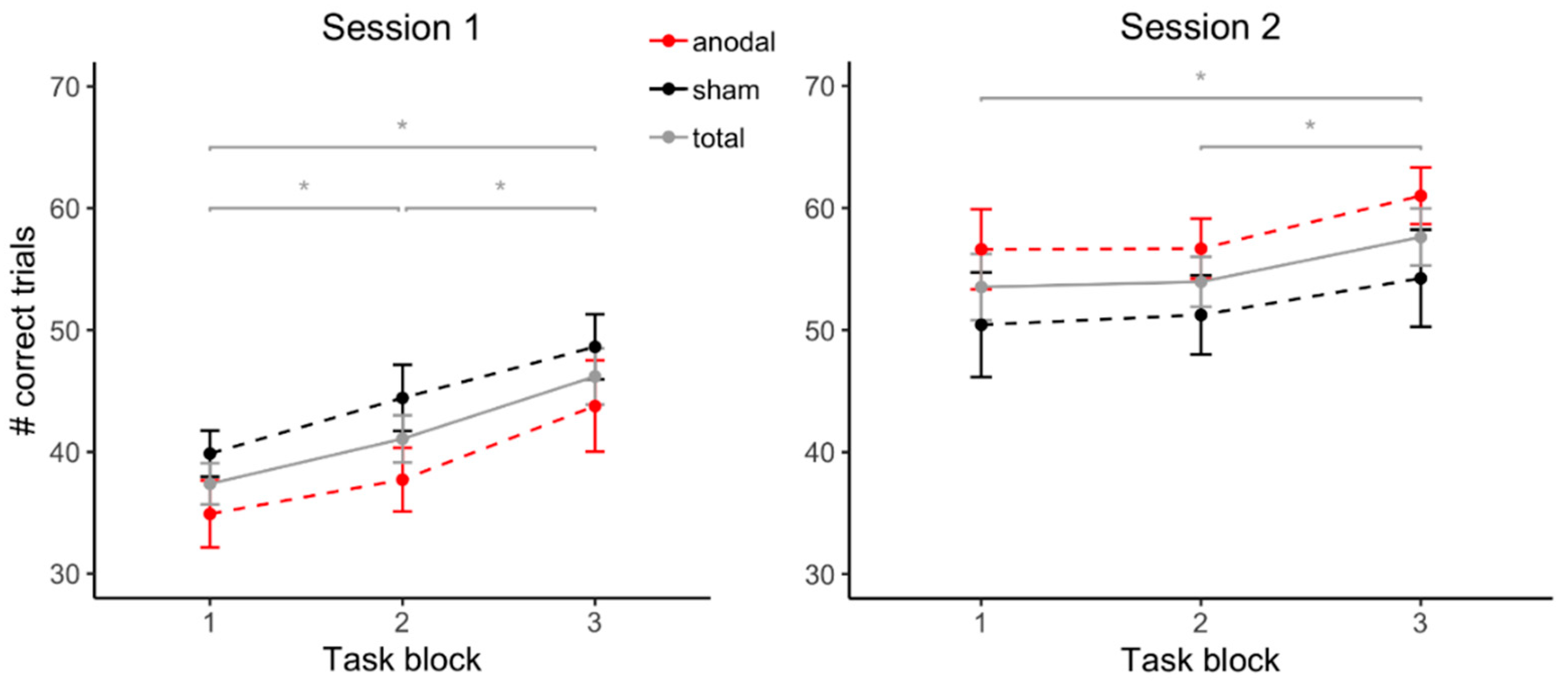
3.2. Task Performance
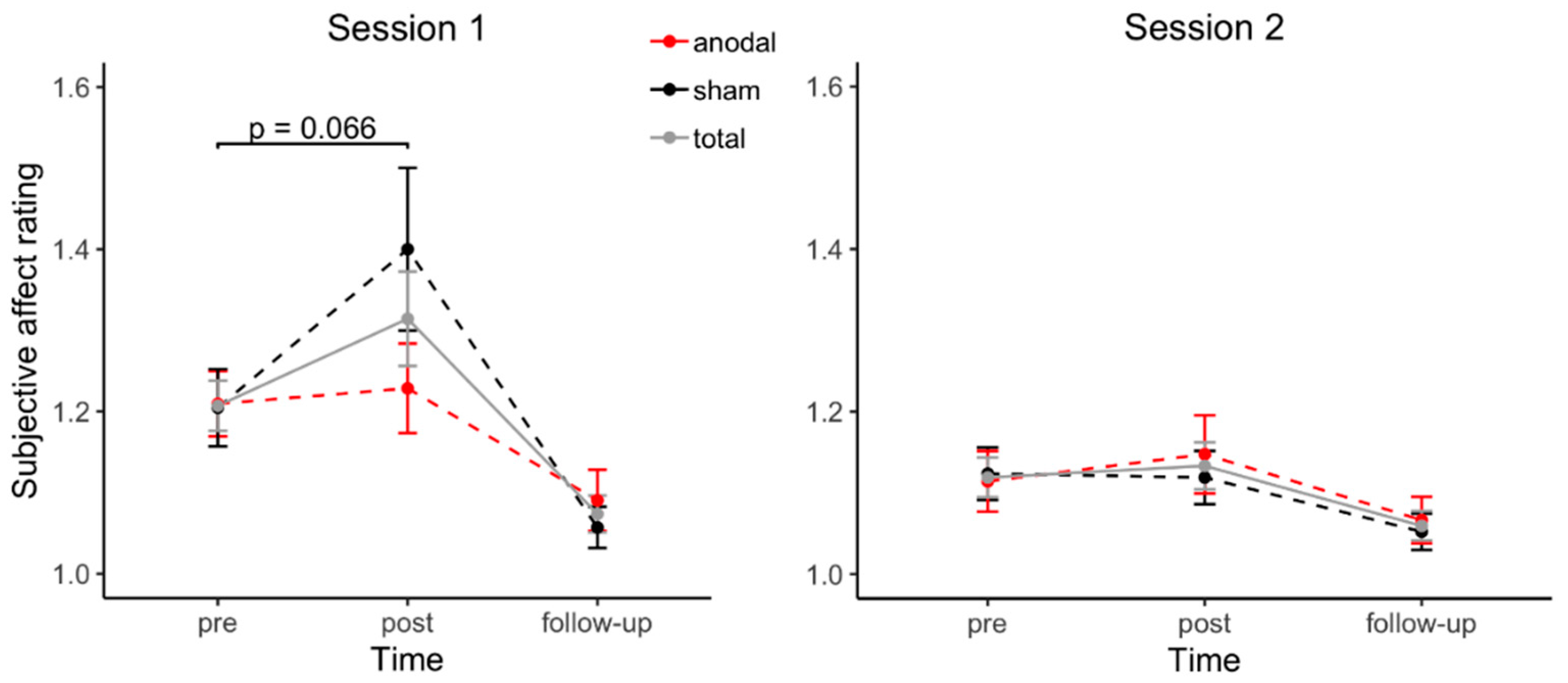
3.3. Affective Changes
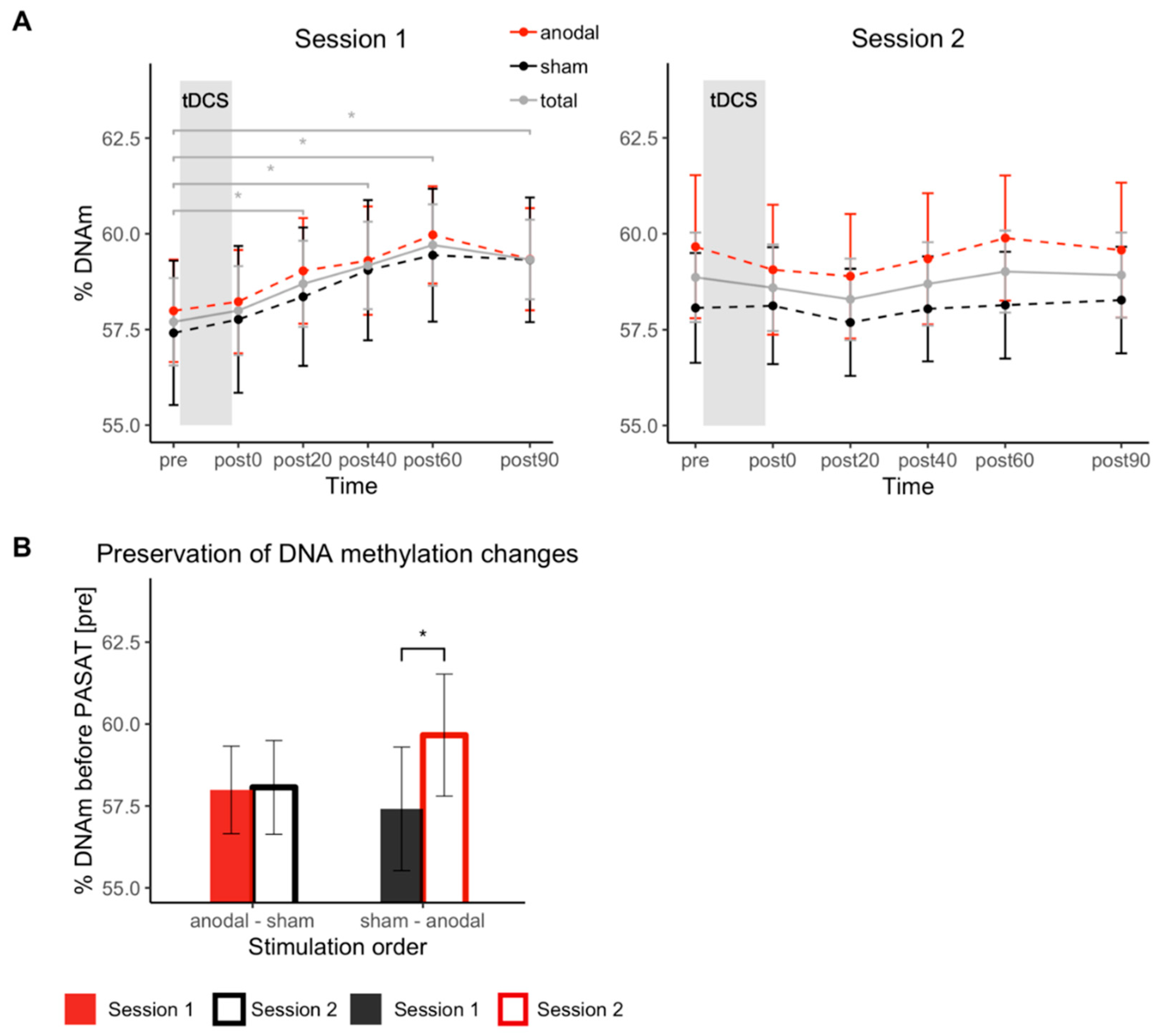
3.4. DNAm Changes in COMT Gene Promoter Region
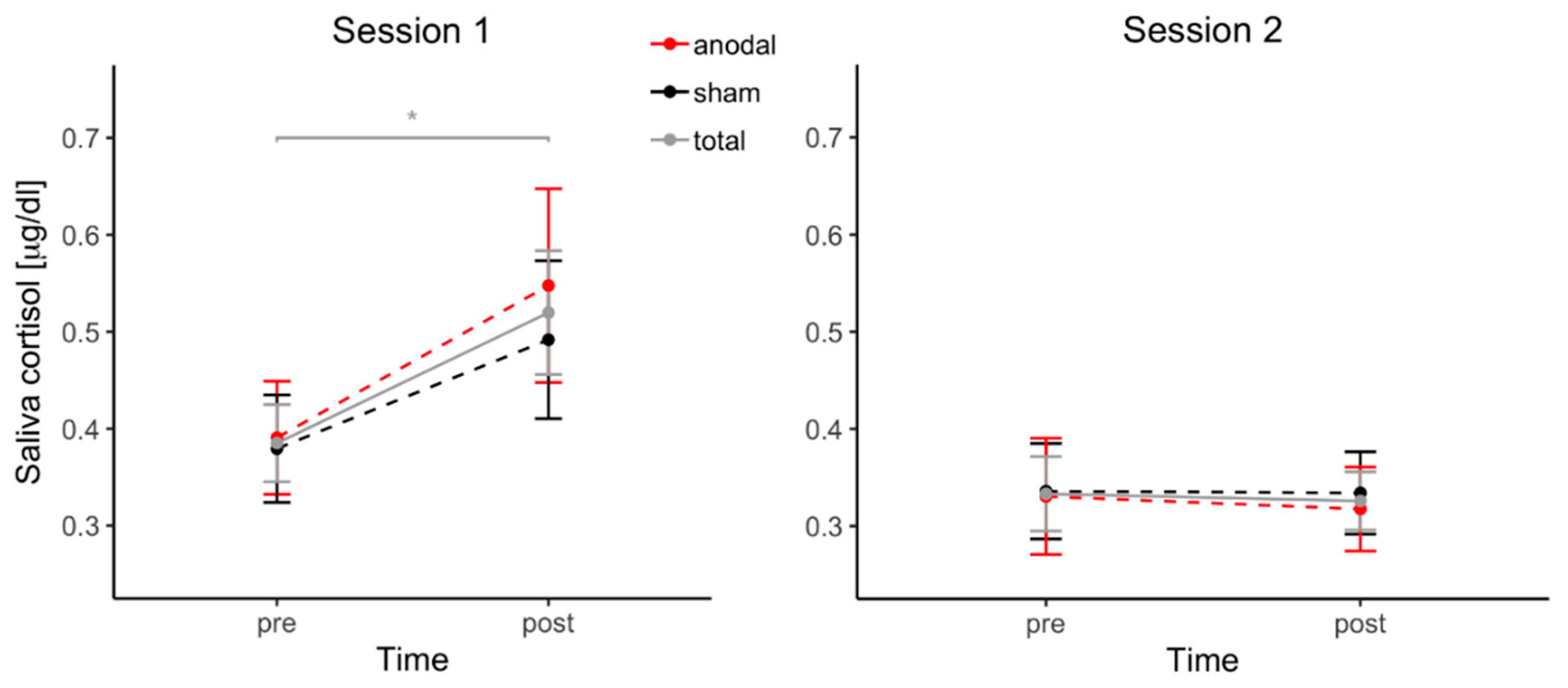
3.5. Cortisol Changes
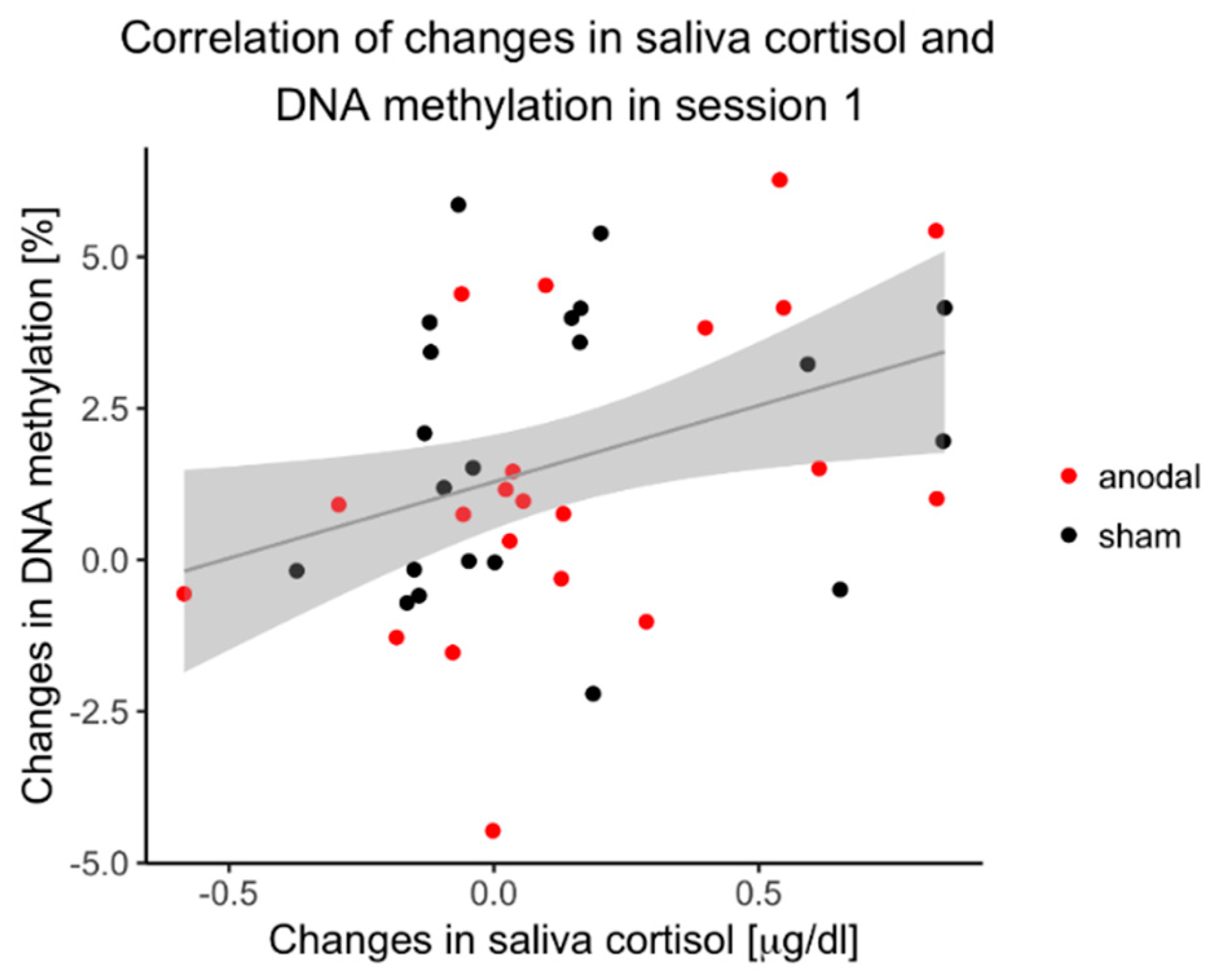
3.6. Correlation of DNAm Changes and Cortisol Changes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klose, R.J.; Bird, A.P. Genomic DNA methylation: The mark and its mediators. Trends Biochem. Sci. 2006, 31, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Vinkers, C.H.; Kalafateli, A.L.; Rutten, B.P.; Kas, M.J.; Kaminsky, Z.; Turner, J.D.; Boks, M.P. Traumatic stress and human DNA methylation: A critical review. Epigenomics 2015, 7, 593–608. [Google Scholar] [CrossRef]
- Klengel, T.; Pape, J.; Binder, E.B.; Mehta, D. The role of DNA methylation in stress-related psychiatric disorders. Neuropharmacology 2014, 80, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Provençal, N.; Binder, E.B. The effects of early life stress on the epigenome: From the womb to adulthood and even before. Exp. Neurol. 2015, 268, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Weaver, I.C.; Cervoni, N.; Champagne, F.A.; D’Alessio, A.C.; Sharma, S.; Seckl, J.R.; Dymov, S.; Szyf, M.; Meaney, M.J. Epigenetic programming by maternal behavior. Nat. Neurosci. 2004, 7, 847. [Google Scholar] [CrossRef] [PubMed]
- Zannas, A.S.; Provençal, N.; Binder, E.B. Epigenetics of posttraumatic stress disorder: Current evidence, challenges, and future directions. Biol. Psychiatry 2015, 78, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Thaler, R.; Spitzer, S.; Karlic, H.; Klaushofer, K.; Varga, F. DMSO is a strong inducer of DNA hydroxymethylation in pre-osteoblastic MC3T3-E1 cells. Epigenetics 2012, 7, 635–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, K.; Li, Y.-S.; Song, M.-F.; Kasai, H. DNA methylation by dimethyl sulfoxide and methionine sulfoxide triggered by hydroxyl radical and implications for epigenetic modifications. Bioorganic Med. Chem. Lett. 2010, 20, 260–265. [Google Scholar] [CrossRef]
- Iwatani, M.; Ikegami, K.; Kremenska, Y.; Hattori, N.; Tanaka, S.; Yagi, S.; Shiota, K. Dimethyl sulfoxide has an impact on epigenetic profile in mouse embryoid body. Stem Cells 2006, 24, 2549–2556. [Google Scholar] [CrossRef]
- Bruniquel, D.; Schwartz, R.H. Selective, stable demethylation of the interleukin-2 gene enhances transcription by an active process. J. Nat. Immunol. 2003, 4, 235. [Google Scholar] [CrossRef] [PubMed]
- Unternaehrer, E.; Luers, P.; Mill, J.; Dempster, E.; Meyer, A.H.; Staehli, S.; Lieb, R.; Hellhammer, D.H.; Meinlschmidt, G. Dynamic changes in DNA methylation of stress-associated genes (OXTR, BDNF) after acute psychosocial stress. Transl. Psychiatry 2012, 2, e150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitsche, M.A.; Cohen, L.G.; Wassermann, E.M.; Priori, A.; Lang, N.; Antal, A.; Paulus, W.; Hummel, F.; Boggio, P.S.; Fregni, F. Transcranial direct current stimulation: State of the art 2008. Brain Stimul. 2008, 1, 206–223. [Google Scholar] [CrossRef] [PubMed]
- Weller, S.; Nitsche, M.A.; Plewnia, C. Cognitive control training and transcranial direct current stimulation: A systematic approach to optimisation. Brain Stimul. 2020, 13, 1358–1369. [Google Scholar] [CrossRef] [PubMed]
- Kuo, M.F.; Nitsche, M.A. Effects of transcranial electrical stimulation on cognition. Clin. EEG Neurosci. 2012, 43, 192–199. [Google Scholar] [CrossRef]
- Moffa, A.H.; Brunoni, A.R.; Nikolin, S.; Loo, C.K. Transcranial direct current stimulation in psychiatric disorders: A comprehensive review. Psychiatr. Clin. 2018, 41, 447–463. [Google Scholar]
- Wiegand, A.; Nieratschker, V.; Plewnia, C. Genetic Modulation of Transcranial Direct Current Stimulation Effects on Cognition. Front. Hum Neurosci. 2016, 10, 651. [Google Scholar] [CrossRef] [Green Version]
- Lachman, H.M.; Papolos, D.F.; Saito, T.; Yu, Y.-M.; Szumlanski, C.L.; Weinshilboum, R.M. Human catechol-O-methyltransferase pharmacogenetics: Description of a functional polymorphism and its potential application to neuropsychiatric disorders. Pharmacogenetics 1996, 6, 243–250. [Google Scholar] [CrossRef]
- Stephens, J.A.; Jones, K.T.; Berryhill, M.E. Task demands, tDCS intensity, and the COMT val 158 met polymorphism impact tDCS-linked working memory training gains. Sci. Rep. 2017, 7, 13463. [Google Scholar] [CrossRef] [Green Version]
- Schacht, J.P. COMT val158met moderation of dopaminergic drug effects on cognitive function: A critical review. Pharm. J. 2016, 16, 430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geller, S.; Wilhelm, O.; Wacker, J.; Hamm, A.; Hildebrandt, A. Associations of the COMT Val158Met polymorphism with working memory and intelligence–A review and meta-analysis. Intelligence 2017, 65, 75–92. [Google Scholar] [CrossRef]
- Cools, R.; D’Esposito, M. Inverted-U–shaped dopamine actions on human working memory and cognitive control. Biol. Psychiatry 2011, 69, e113–e125. [Google Scholar] [CrossRef] [Green Version]
- Grace, A.A. Dysregulation of the dopamine system in the pathophysiology of schizophrenia and depression. Nat. Rev. Neurosci. 2016, 17, 524. [Google Scholar] [CrossRef]
- Abercrombie, E.D.; Keefe, K.A.; DiFrischia, D.S.; Zigmond, M.J. Differential effect of stress on in vivo dopamine release in striatum, nucleus accumbens, and medial frontal cortex. J. Neurochem. 1989, 52, 1655–1658. [Google Scholar] [CrossRef]
- Nagano-Saito, A.; Dagher, A.; Booij, L.; Gravel, P.; Welfeld, K.; Casey, K.F.; Leyton, M.; Benkelfat, C. Stress-induced dopamine release in human medial prefrontal cortex—18F-Fallypride/PET study in healthy volunteers. Synapse 2013, 67, 821–830. [Google Scholar] [CrossRef]
- Desbonnet, L.; Tighe, O.; Karayiorgou, M.; Gogos, J.A.; Waddington, J.L.; O’Tuathaigh, C.M. Physiological and behavioural responsivity to stress and anxiogenic stimuli in COMT-deficient mice. Behav. Brain Res. 2012, 228, 351–358. [Google Scholar] [CrossRef]
- Armbruster, D.; Mueller, A.; Strobel, A.; Lesch, K.-P.; Brocke, B.; Kirschbaum, C. Children under stress–COMT genotype and stressful life events predict cortisol increase in an acute social stress paradigm. Int. J. Neuropsychopharmacol. 2012, 15, 1229–1239. [Google Scholar] [CrossRef] [Green Version]
- Walder, D.J.; Trotman, H.D.; Cubells, J.F.; Brasfield, J.; Tang, Y.; Walker, E.F. Catechol-O-Methyltransferase (COMT) modulation of cortisol secretion in psychiatrically at-risk and healthy adolescents. Psychiatr. Genet. 2010, 20, 166. [Google Scholar] [CrossRef] [Green Version]
- Wiegand, A.; Sommer, A.; Nieratschker, V.; Plewnia, C. Improvement of cognitive control and stabilization of affect by prefrontal transcranial direct current stimulation (tDCS). Sci. Rep. 2019, 9, 6797. [Google Scholar] [CrossRef] [PubMed]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Watson, D.; Clark, L.A.; Tellegen, A. Development and validation of brief measures of positive and negative affect: The PANAS scales. J. Pers. Soc. Psychol. 1988, 54, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Krohne, H.W.; Egloff, B.; Kohlmann, C.-W.; Tausch, A. Untersuchungen mit einer deutschen Version der “Positive and Negative Affect Schedule” (PANAS). Diagn. Gott. 1996, 42, 139–156. [Google Scholar]
- Jasper, H.H. The ten twenty electrode system of the international federation. Electroencephalogr. Clin. Neurophysiol. 1958, 10, 371–375. [Google Scholar]
- Priori, A.; Mameli, F.; Cogiamanian, F.; Marceglia, S.; Tiriticco, M.; Mrakic-Sposta, S.; Ferrucci, R.; Zago, S.; Polezzi, D.; Sartori, G. Lie-specific involvement of dorsolateral prefrontal cortex in deception. Cereb. Cortex 2008, 18, 451–455. [Google Scholar] [CrossRef] [Green Version]
- Derogatis, L.R. Symptom Checklist-90-R [SCL-90-R] Administration, Scoring, and Procedures Manual, 3rd ed.; National Computer Systems: Minneapolis, MN, USA, 1994. [Google Scholar]
- Mill, J.; Dempster, E.; Caspi, A.; Williams, B.; Moffitt, T.; Craig, I. Evidence for monozygotic twin (MZ) discordance in methylation level at two CpG sites in the promoter region of the catechol-O-methyltransferase (COMT) gene. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2006, 141, 421–425. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 27 November 2018).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects Models, R Package Version 3.1-137. 2018. Available online: https://CRAN.R-project.org/package=nlme (accessed on 27 November 2018).
- Menard, S. An introduction to logistic regression diagnostics. In Applied Logistic Regression Analysis; Sage: Washington, DC, USA, 1995; pp. 58–79. [Google Scholar]
- Alin, A. Multicollinearity. Wiley Interdiscip. Rev. Comput. Stat. 2010, 2, 370–374. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D. Mixed-Effects Models in S and S-PLUS; Springer Science & Business Media: Luxembourg, 2006. [Google Scholar]
- Reik, W. Stability and flexibility of epigenetic gene regulation in mammalian development. Nature 2007, 447, 425. [Google Scholar] [CrossRef] [PubMed]
- Yehuda, R.; Bierer, L.M. The relevance of epigenetics to PTSD: Implications for the DSM-V. J. Trauma. Stress 2009, 22, 427–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Höhne, N.; Poidinger, M.; Merz, F.; Pfister, H.; Brückl, T.; Zimmermann, P.; Uhr, M.; Holsboer, F.; Ising, M. FKBP5 genotype-dependent DNA methylation and mRNA regulation after psychosocial stress in remitted depression and healthy controls. Int. J. Neuropsychopharmacol. 2015, 18, pyu087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klengel, T.; Mehta, D.; Anacker, C.; Rex-Haffner, M.; Pruessner, J.C.; Pariante, C.M.; Pace, T.W.; Mercer, K.B.; Mayberg, H.S.; Bradley, B. Allele-specific FKBP5 DNA demethylation mediates gene–childhood trauma interactions. Nat. Neurosci. 2013, 16, 33. [Google Scholar] [CrossRef] [Green Version]
- Braun, P.R.; Tanaka-Sahker, M.; Chan, A.C.; Jellison, S.S.; Klisares, M.J.; Hing, B.W.; Shabbir, Y.; Gaul, L.N.; Nagahama, Y.; Robles, J. Genome-wide DNA methylation investigation of glucocorticoid exposure within buccal samples. Psychiatry Clin. Neurosci. 2019, 73, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Winkler, E.A.; Yue, J.K.; Ferguson, A.R.; Temkin, N.R.; Stein, M.B.; Barber, J.; Yuh, E.L.; Sharma, S.; Satris, G.G.; McAllister, T.W. COMT Val158Met polymorphism is associated with post-traumatic stress disorder and functional outcome following mild traumatic brain injury. J. Clin. Neurosci. 2017, 35, 109–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano, J.M.; Banks, J.B.; Fagan, T.J.; Tartar, J.L. The influence of Val158Met COMT on physiological stress responsivity. Stress 2019, 22, 276–279. [Google Scholar] [CrossRef]
- Ursini, G.; Bollati, V.; Fazio, L.; Porcelli, A.; Iacovelli, L.; Catalani, A.; Sinibaldi, L.; Gelao, B.; Romano, R.; Rampino, A. Stress-related methylation of the catechol-O-methyltransferase Val158 allele predicts human prefrontal cognition and activity. J. Neurosci. 2011, 31, 6692–6698. [Google Scholar] [CrossRef] [Green Version]
- Hosak, L. Role of the COMT gene Val158Met polymorphism in mental disorders: A review. Eur. Psychiatry 2007, 22, 276–281. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.C.; O’Reilly, R.L.; Singh, S.M. Site-specific cytosine methylation in S-COMT promoter in 31 brain regions with implications for studies involving schizophrenia. Am. J. Med Genet. Part B Neuropsychiatr. Genet. 2005, 133, 37–42. [Google Scholar] [CrossRef]
- Teruhisa, U.; Ryoji, H.; Taisuke, I.; Tatsuya, S.; Fumihiro, M.; Tatsuo, S. Use of saliva for monitoring unbound free cortisol levels in serum. Clin. Chim. Acta 1981, 110, 245–253. [Google Scholar] [CrossRef]
- Tenhunen, J.; Salminen, M.; Lundström, K.; Kiviluoto, T.; Savolainen, R.; Ulmanen, I. Genomic organization of the human catechol O-methyltransferase gene and its expression from two distinct promoters. Eur. J. Biochem. 1994, 223, 1049–1059. [Google Scholar] [CrossRef]
- Jjingo, D.; Conley, A.B.; Soojin, V.Y.; Lunyak, V.V.; Jordan, I.K. On the presence and role of human gene-body DNA methylation. Oncotarget 2012, 3, 462. [Google Scholar] [CrossRef] [Green Version]
- Gronwall, D. Paced auditory serial-addition task: A measure of recovery from concussion. Percept. Mot. Ski. 1977, 44, 367–373. [Google Scholar] [CrossRef]
- Lejuez, C.; Kahler, C.W.; Brown, R.A. A modified computer version of the Paced Auditory Serial Addition Task (PASAT) as a laboratory-based stressor. Behav. Ther. 2003, 26, 290–293. [Google Scholar]
- Plewnia, C.; Schroeder, P.A.; Kunze, R.; Faehling, F.; Wolkenstein, L. Keep calm and carry on: Improved frustration tolerance and processing speed by transcranial direct current stimulation (tDCS). PLoS ONE 2015, 10, e0122578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolkenstein, L.; Plewnia, C. Amelioration of cognitive control in depression by transcranial direct current stimulation. Biol. Psychiatry 2013, 73, 646–651. [Google Scholar] [CrossRef]
- Brunoni, A.R.; Vanderhasselt, M.-A.; Boggio, P.S.; Fregni, F.; Dantas, E.M.; Mill, J.G.; Lotufo, P.A.; Benseñor, I.M. Polarity-and valence-dependent effects of prefrontal transcranial direct current stimulation on heart rate variability and salivary cortisol. Psychoneuroendocrinology 2013, 38, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, A.; Dowker, A.; Cohen Kadosh, R. Cognitive enhancement or cognitive cost: Trait-specific outcomes of brain stimulation in the case of mathematics anxiety. J. Neurosci. 2014, 34, 16605–16610. [Google Scholar] [CrossRef]
- Podda, M.V.; Cocco, S.; Mastrodonato, A.; Fusco, S.; Leone, L.; Barbati, S.A.; Colussi, C.; Ripoli, C.; Grassi, C. Anodal transcranial direct current stimulation boosts synaptic plasticity and memory in mice via epigenetic regulation of Bdnf expression. Sci. Rep. 2016, 6, 22180. [Google Scholar] [CrossRef]
- Komaki, S.; Shiwa, Y.; Furukawa, R.; Hachiya, T.; Ohmomo, H.; Otomo, R.; Satoh, M.; Hitomi, J.; Sobue, K.; Sasaki, M. iMETHYL: An integrative database of human DNA methylation, gene expression, and genomic variation. Hum. Genome Var. 2018, 5, 18008. [Google Scholar] [CrossRef] [Green Version]
- Hachiya, T.; Furukawa, R.; Shiwa, Y.; Ohmomo, H.; Ono, K.; Katsuoka, F.; Nagasaki, M.; Yasuda, J.; Fuse, N.; Kinoshita, K. Genome-wide identification of inter-individually variable DNA methylation sites improves the efficacy of epigenetic association studies. NPJ Genom. Med. 2017, 2, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tohoku, I. Medical Megabank Organization, iMETHYL Database; over 100 Japanese whole Genome DNA Methylation Database from Monocytes, CD4+ T Cells, and Neutrophils. Available online: http://imethyl.iwate-megabank.org/ (accessed on 21 March 2019).
- Koch, A.J. Immune response to exercise. Braz. J. Biomotricity 2010, 4, 92–103. [Google Scholar]
- Davis, A.; Maney, D.; Maerz, J. The use of leukocyte profiles to measure stress in vertebrates: A review for ecologists. Funct. Ecol. 2008, 22, 760–772. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Hoffman-Goetz, L. Exercise and the immune system: Regulation, integration, and adaptation. Physiol. Rev. 2000, 80, 1055–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tønnesen, E.; Christensen, N.; Brinkløv, M. Natural killer cell activity during cortisol and adrenaline infusion in healthy volunteers. Eur. J. Clin. Investig. 1987, 17, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Schiele, M.A.; Gottschalk, M.G.; Domschke, K. The applied implications of epigenetics in anxiety, affective and stress-related disorders-A review and synthesis on psychosocial stress, psychotherapy and prevention. Clin. Psychol. Rev. 2020, 77, 101830. [Google Scholar] [CrossRef] [PubMed]
- Minarik, T.; Berger, B.; Althaus, L.; Bader, V.; Biebl, B.; Brotzeller, F.; Fusban, T.; Hegemann, J.; Jesteadt, L.; Kalweit, L. The importance of sample size for reproducibility of tDCS effects. Front. Hum. Neurosci. 2016, 10, 453. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wiegand, A.; Blickle, A.; Brückmann, C.; Weller, S.; Nieratschker, V.; Plewnia, C. Dynamic DNA Methylation Changes in the COMT Gene Promoter Region in Response to Mental Stress and Its Modulation by Transcranial Direct Current Stimulation. Biomolecules 2021, 11, 1726. https://doi.org/10.3390/biom11111726
Wiegand A, Blickle A, Brückmann C, Weller S, Nieratschker V, Plewnia C. Dynamic DNA Methylation Changes in the COMT Gene Promoter Region in Response to Mental Stress and Its Modulation by Transcranial Direct Current Stimulation. Biomolecules. 2021; 11(11):1726. https://doi.org/10.3390/biom11111726
Chicago/Turabian StyleWiegand, Ariane, Arne Blickle, Christof Brückmann, Simone Weller, Vanessa Nieratschker, and Christian Plewnia. 2021. "Dynamic DNA Methylation Changes in the COMT Gene Promoter Region in Response to Mental Stress and Its Modulation by Transcranial Direct Current Stimulation" Biomolecules 11, no. 11: 1726. https://doi.org/10.3390/biom11111726