
Interaction of the Escherichia coli HU Protein with Various Topological Forms of DNA


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Overproduction and Purification of HU Protein
2.2. Tritium Labeling of HU Protein
2.3. Topoisomers of Plasmid pRLM4
2.4. Gel Retardation Assays
2.5. Topological Assays
2.6. Nick Closure Assay
2.7. Chemical Crosslinking
3. Results
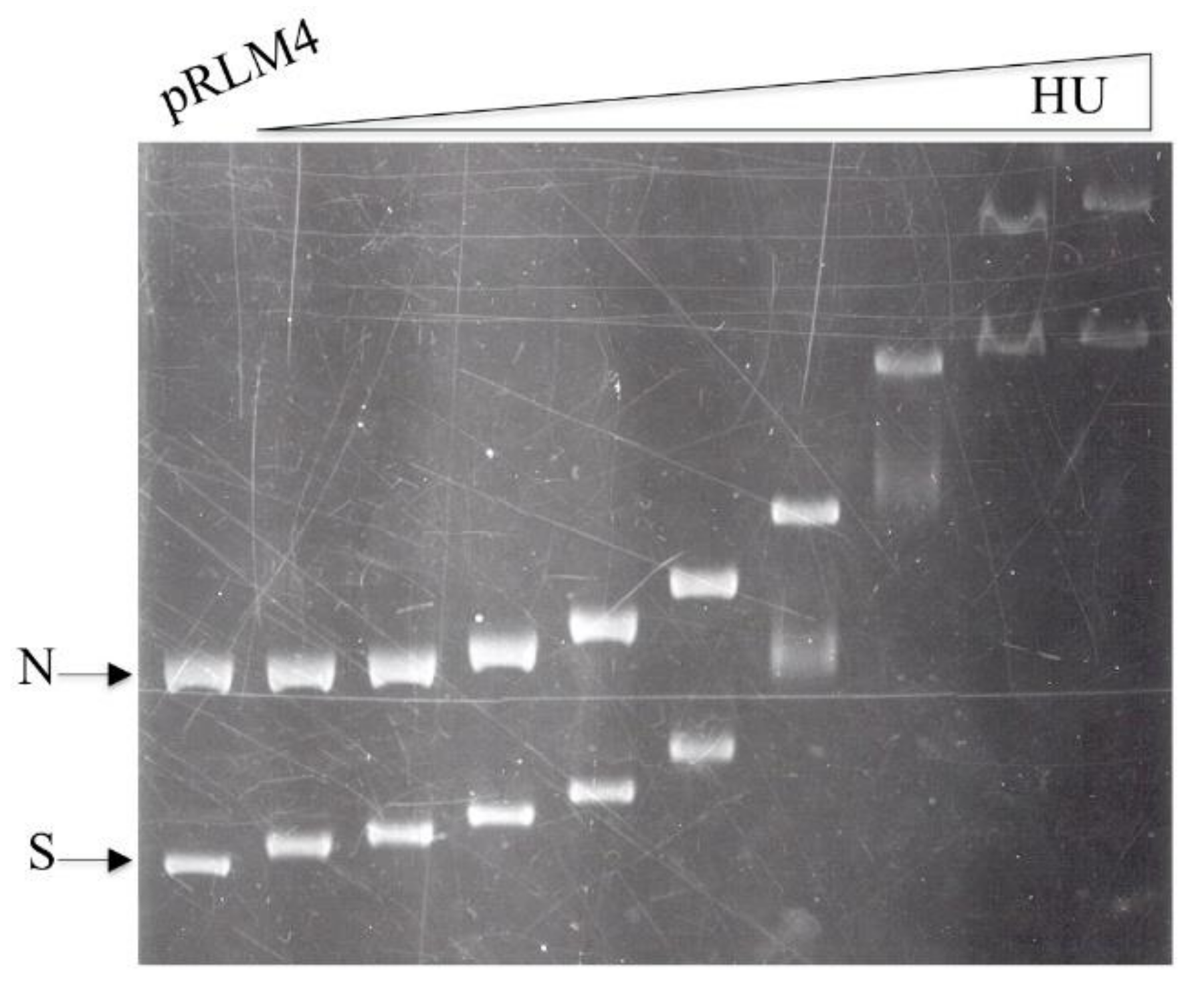
3.1. Binding of HU to Negatively Supercoiled and Nicked Circular DNA
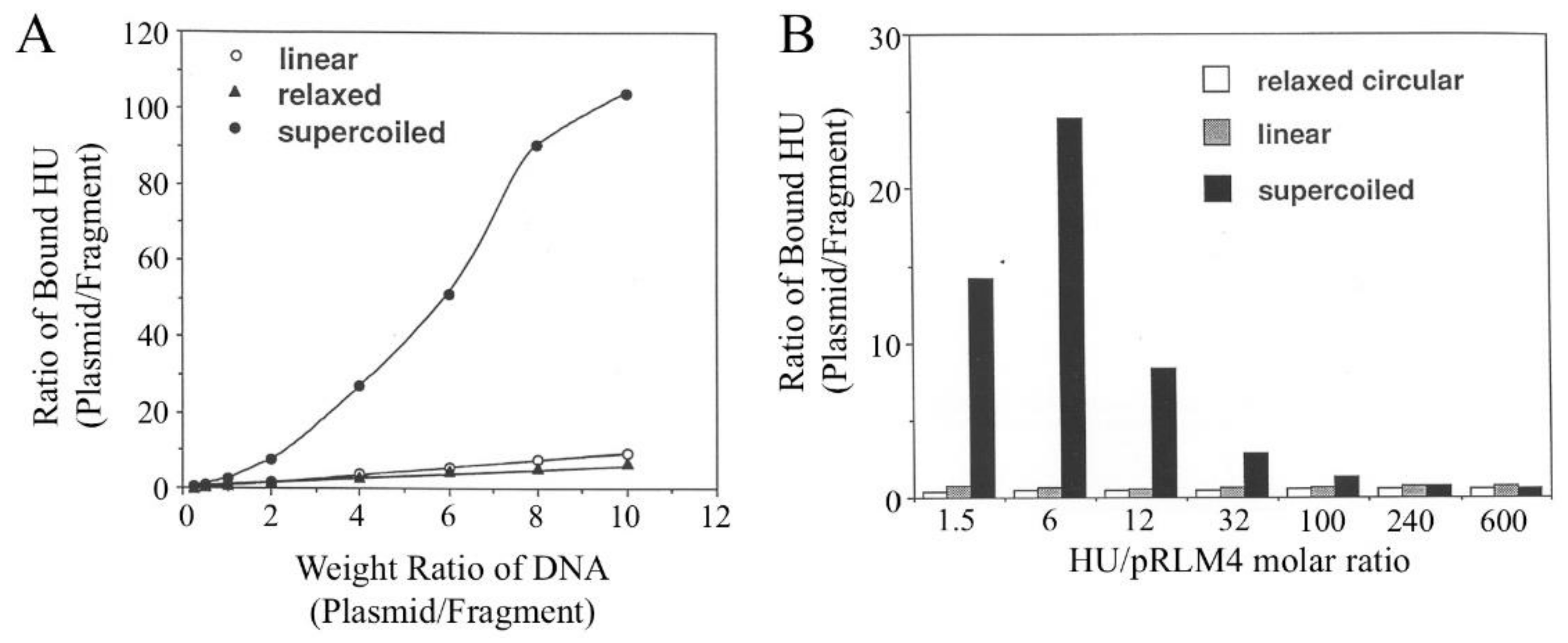
3.2. Affinities of HU for Different Forms of pRLM4
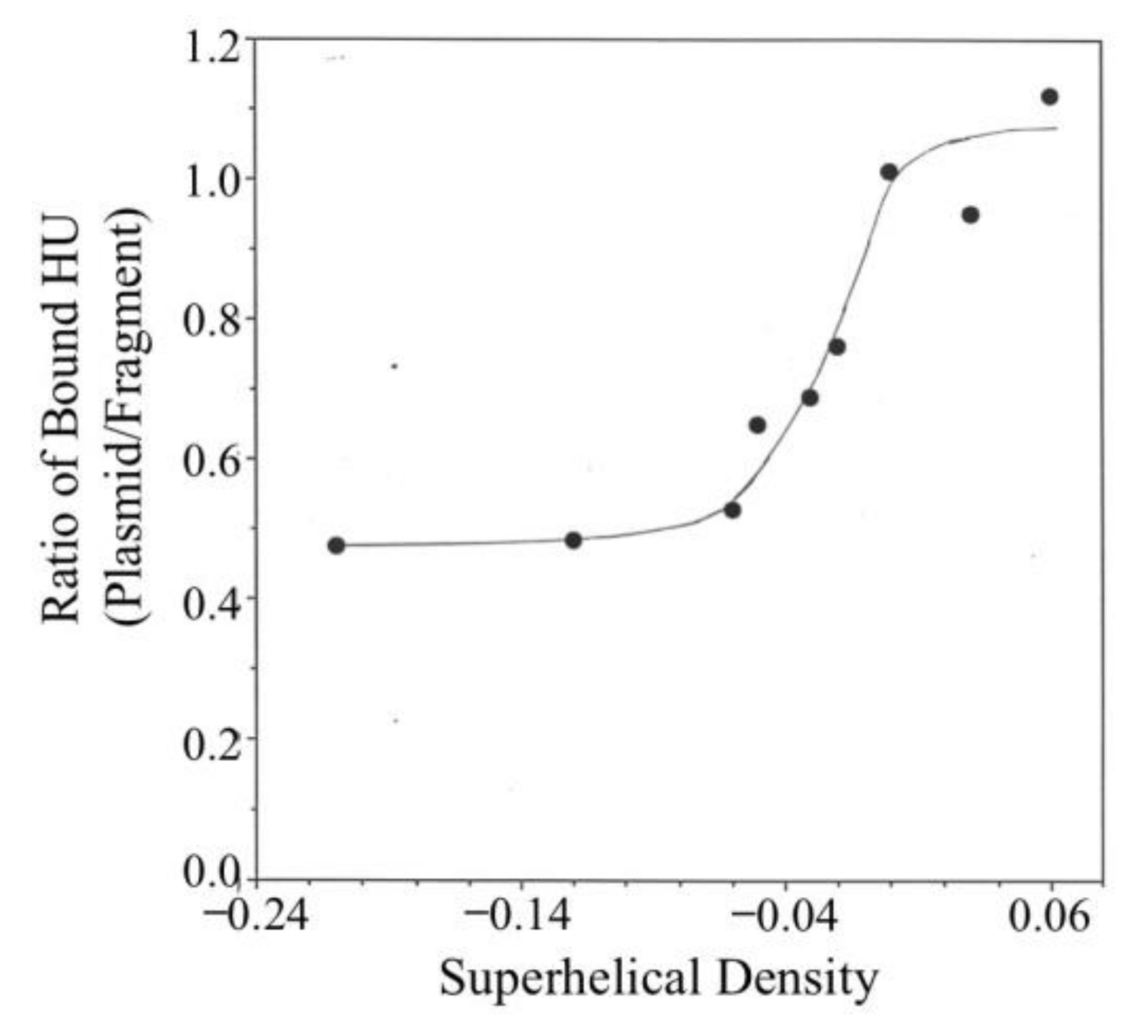
3.3. Binding of HU to pRLM4 Topoisomers
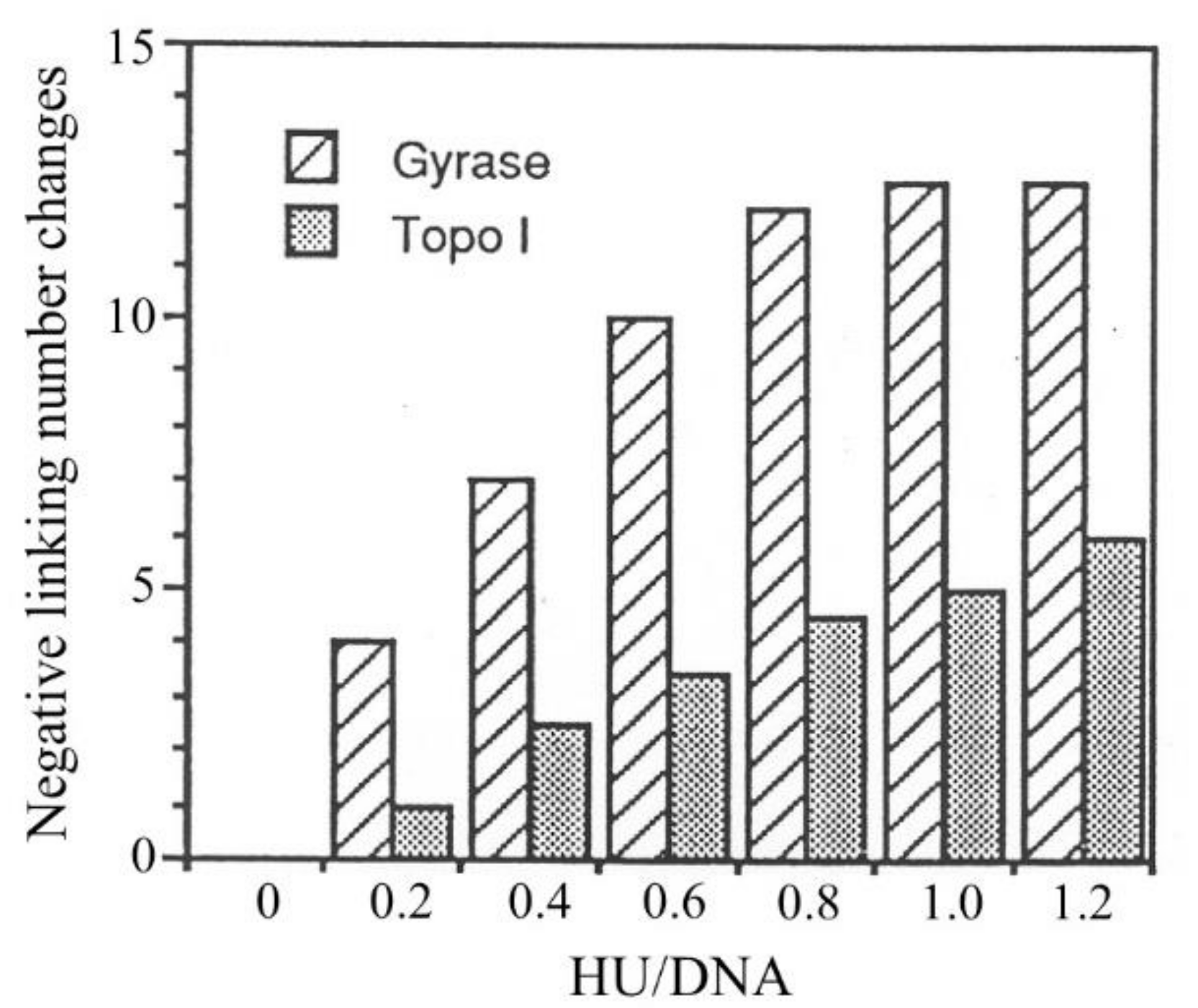
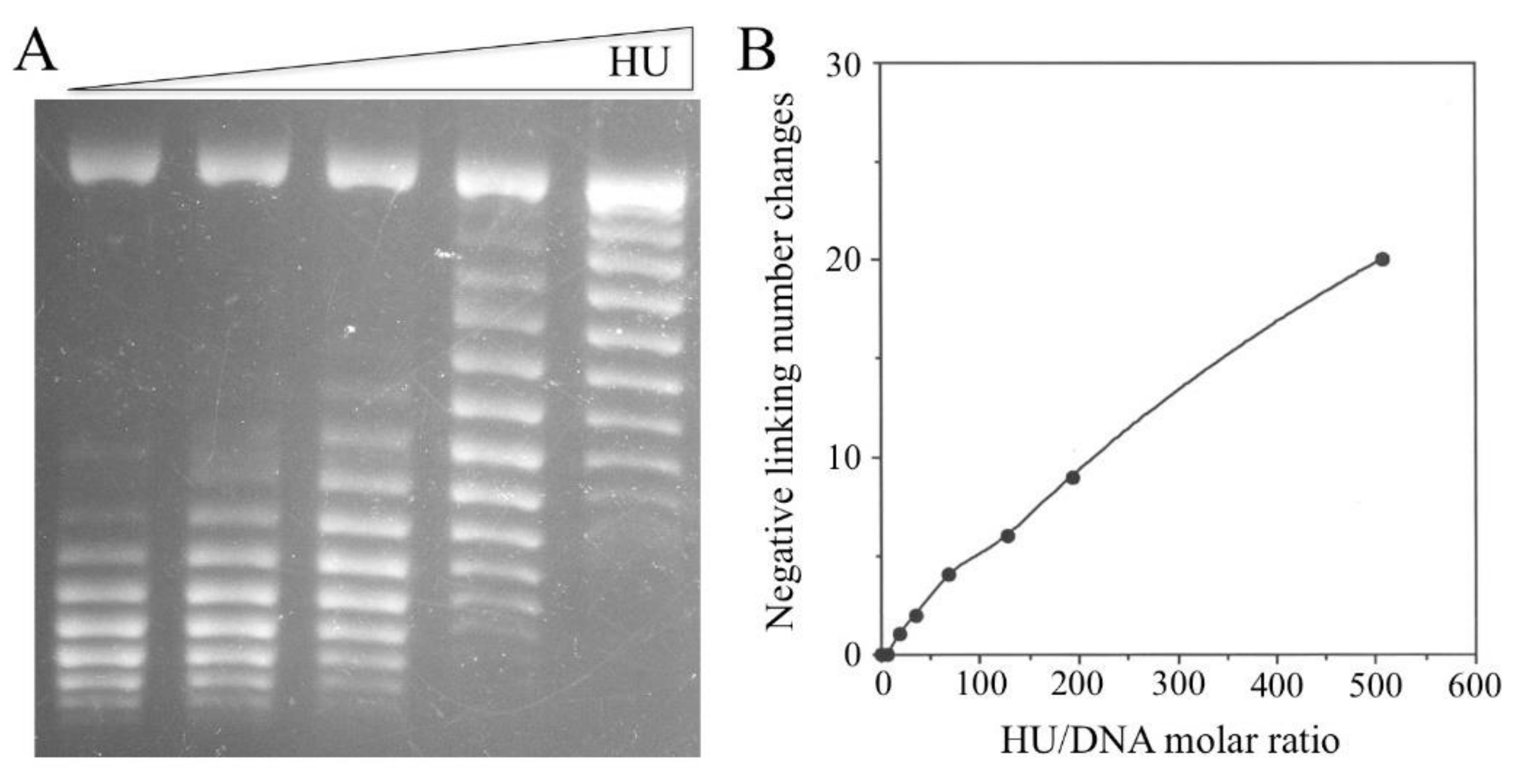
3.4. Constraining of Negative Supercoils by HU
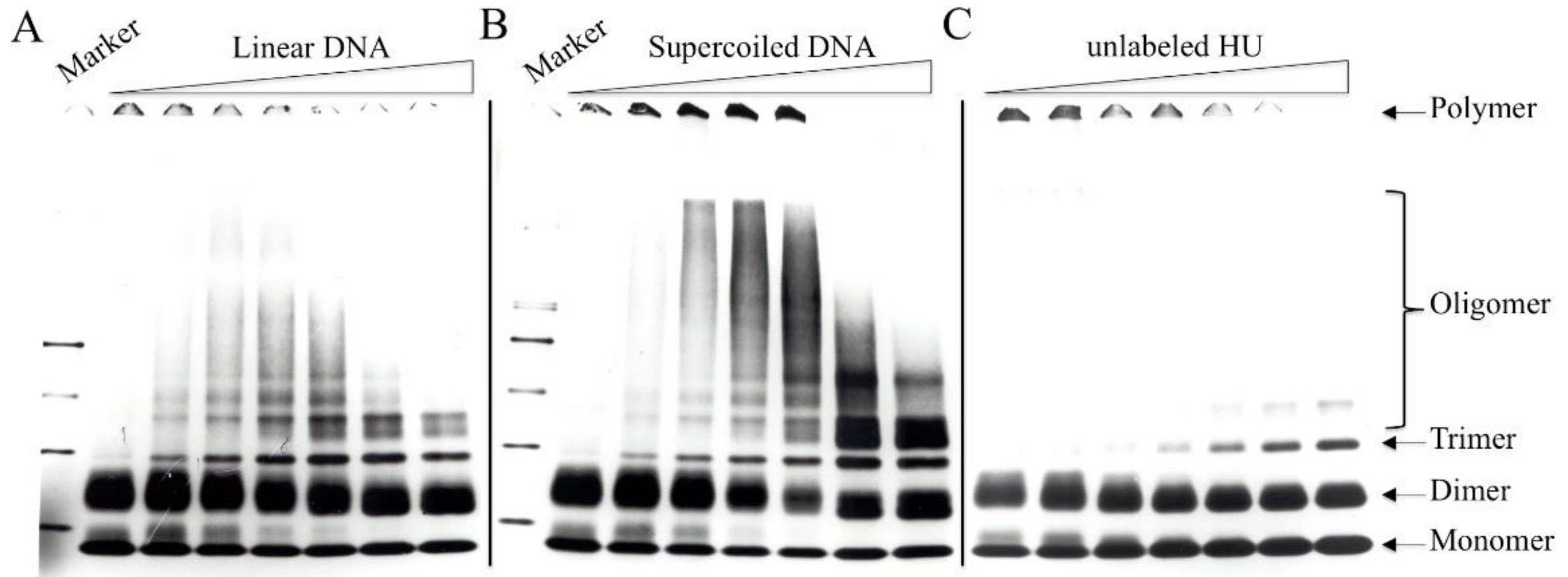
3.5. Chemical Crosslinking of DNA-Bound HU
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dame, R.T.; Rashid, F.Z.M.; Grainger, D.C. Chromosome organization in bacteria: Mechanistic insights into genome structure and function. Nat. Rev. Genet. 2020, 21, 227–242. [Google Scholar] [CrossRef]
- Qin, L.; Erkelens, A.M.; Ben Bdira, F.; Dame, R.T. The architects of bacterial DNA bridges: A structurally and functionally conserved family of proteins. Open Biol. 2019, 9, 190223. [Google Scholar] [CrossRef] [Green Version]
- Dorman, C.J. Genome architecture and global gene regulation in bacteria: Making progress towards a unified model? Nat. Rev. Microbiol. 2013, 11, 349–355. [Google Scholar] [CrossRef]
- Verma, S.C.; Qian, Z.; Adhya, S.L. Architecture of the Escherichia coli nucleoid. PLoS Genet. 2020, 16, e1008456. [Google Scholar] [CrossRef]
- Azam, T.A.; Ishihama, A. Twelve species of the nucleoid-associated protein from Escherichia coli—Sequence recognition specificity and DNA binding affinity. J. Biol. Chem. 1999, 274, 33105–33113. [Google Scholar] [CrossRef] [Green Version]
- Luijsterburg, M.S.; White, M.F.; van Driel, R.; Dame, R.T. The Major Architects of Chromatin: Architectural Proteins in Bacteria, Archaea and Eukaryotes. Crit. Rev. Biochem. Mol. 2008, 43, 393–418. [Google Scholar] [CrossRef]
- Megraw, T.L.; Chae, C.B. Functional Complementarity between the Hmg1-Like Yeast Mitochondrial Histone Hm and the Bacterial Histone-Like Protein Hu. J. Biol. Chem. 1993, 268, 12758–12763. [Google Scholar] [CrossRef]
- Paull, T.T.; Johnson, R.C. DNA Looping by Saccharomyces-Cerevisiae High-Mobility Group Proteins Nhp6a/B—Consequences for Nucleoprotein Complex Assembly and Chromatin Condensation. J. Biol. Chem. 1995, 270, 8744–8754. [Google Scholar] [CrossRef] [Green Version]
- Hodges-Garcia, Y.; Hagerman, P.J.; Pettijohn, D.E. DNA Ring-Closure Mediated by Protein Hu. J. Biol. Chem. 1989, 264, 14621–14623. [Google Scholar] [CrossRef]
- Paull, T.T.; Haykinson, M.J.; Johnson, R.C. The Nonspecific DNA-Binding and DNA-Bending Proteins Hmg1 and Hmg2 Promote the Assembly of Complex Nucleoprotein Structures. Gene Dev. 1993, 7, 1521–1534. [Google Scholar] [CrossRef] [Green Version]
- Pettijohn, D.E.; Pfenninger, O. Supercoils in prokaryotic DNA restrained in vivo. Proc. Natl. Acad. Sci. USA 1980, 77, 1331–1335. [Google Scholar] [CrossRef] [Green Version]
- Broyles, S.S.; Pettijohn, D.E. Interaction of the Escherichia coli HU protein with DNA: Evidence for formation of nucleosome-like structures with altered DNA helical pitch. J. Mol. Biol. 1986, 187, 47–60. [Google Scholar] [CrossRef]
- Rouvière-Yaniv, J.; Yaniv, M.; Germond, J.-E.E. coli DNA binding protein HU forms nucleosome-like structure with circular double-stranded DNA. Cell 1979, 17, 265–274. [Google Scholar] [CrossRef]
- Dame, R.T.; Goosen, N. HU: Promoting or counteracting DNA compaction? FEBS Lett. 2002, 529, 151–156. [Google Scholar] [CrossRef] [Green Version]
- van Noort, J.; Verbrugge, S.; Goosen, N.; Dekker, C.; Dame, R. Dual architectural roles of HU: Formation of flexible hinges and rigid filaments. Proc. Natl. Acad. Sci. USA 2004, 101, 6969–6974. [Google Scholar] [CrossRef] [Green Version]
- Pontiggia, A.; Negri, A.; Beltrame, M.; Bianchi, M.E. Protein Hu Binds Specifically to Kinked DNA. Mol. Microbiol. 1993, 7, 343–350. [Google Scholar] [CrossRef]
- Kamashev, D.; Rouviere-Yaniv, J. The histone-like protein HU binds specifically to DNA recombination and repair intermediates. EMBO J. 2000, 19, 6527–6535. [Google Scholar] [CrossRef] [Green Version]
- Bonnefoy, E.; Rouvière-Yaniv, J. Hu and Ihf, 2 Homologous Histone-Like Proteins of Escherichia coli, Form Different Protein DNA Complexes with Short DNA Fragments. EMBO J. 1991, 10, 687–696. [Google Scholar] [CrossRef]
- Bonnefoy, E.; Takahashi, M.; Yaniv, J.R. DNA-Binding Parameters of the Hu Protein of Escherichia coli to Cruciform DNA. J. Mol. Biol. 1994, 242, 116–129. [Google Scholar] [CrossRef]
- Lyubchenko, Y.L.; Shlyakhtenko, L.S.; Aki, T.; Adhya, S. Atomic force microscopic demonstration of DNA looping by GalR and HU. Nucleic Acids Res. 1997, 25, 873–876. [Google Scholar] [CrossRef]
- Castaing, B.; Zelwer, C.; Laval, J.; Boiteux, S. Hu Protein of Escherichia coli Binds Specifically to DNA That Contains Single-Strand Breaks or Gaps. J. Biol. Chem. 1995, 270, 10291–10296. [Google Scholar] [CrossRef] [Green Version]
- Lavoie, B.D.; Chaconas, G. A second high affinity HU binding site in the phage Mu transpososome. J. Biol. Chem. 1994, 269, 15571–15576. [Google Scholar] [CrossRef]
- Kano, Y.; Goshima, N.; Wada, M.; Imamoto, F. Participation of Hup Gene-Product in Replicative Transposition of Mu-Phage in Escherichia coli. Gene 1989, 76, 353–358. [Google Scholar] [CrossRef]
- Ogura, T.; Niki, H.; Kano, Y.; Imamoto, F.; Hiraga, S. Maintenance of Plasmids in Hu and Ihf Mutants of Escherichia coli. Mol. Gen. Genet. 1990, 220, 197–203. [Google Scholar] [CrossRef]
- Hwang, D.S.; Kornberg, A. Opening of the Replication Origin of Escherichia coli by Dnaa Protein with Protein Hu or Ihf. J. Biol. Chem. 1992, 267, 23083–23086. [Google Scholar] [CrossRef]
- Azam, T.A.; Iwata, A.; Nishimura, A.; Ueda, S.; Ishihama, A. Growth phase-dependent variation in protein composition of the Escherichia coli nucleoid. J. Bacteriol. 1999, 181, 6361–6370. [Google Scholar] [CrossRef] [Green Version]
- Prieto, A.I.; Kahramanoglou, C.; Ali, R.M.; Fraser, G.; Seshasayee, A.S.N.; Luscombe, N.M. Genomic analysis of DNA binding and gene regulation by homologous nucleoid-associated proteins IHF and HU in Escherichia coli K12. Nucleic Acids Res. 2012, 40, 3524–3537. [Google Scholar] [CrossRef]
- Macvanin, M.; Edgar, R.; Cui, F.; Trostel, A.; Zhurkin, V.; Adhya, S. Noncoding RNAs Binding to the Nucleoid Protein HU in Escherichia coli. J. Bacteriol. 2012, 194, 6046–6055. [Google Scholar] [CrossRef] [Green Version]
- Hammel, M.; Amlanjyoti, D.; Reyes, F.E.; Chen, J.-H.; Parpana, R.; Tang, H.Y.H.; Larabell, C.A.; Tainer, J.A.; Adhya, S. HU multimerization shift controls nucleoid compaction. Sci. Adv. 2016, 2, e1600650. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, H.; Yasuzawa, K.; Kohno, K.; Goshima, N.; Kano, Y.; Saiki, T.; Imamoto, F. Role of Hu Proteins in Forming and Constraining Supercoils of Chromosomal DNA in Escherichia coli. Mol. Gen. Genet. 1995, 248, 518–526. [Google Scholar] [CrossRef]
- Guo, F.S.; Adhya, S. Spiral structure of Escherichia coli HU alpha beta provides foundation for DNA supercoiling. Proc. Natl. Acad. Sci. USA 2007, 104, 4309–4314. [Google Scholar] [CrossRef] [Green Version]
- Kundukad, B.; Cong, P.; Van Der Maarel, J.R.C.; Doyle, P.S. Time-dependent bending rigidity and helical twist of DNA by rearrangement of bound HU protein. Nucleic Acids Res. 2013, 41, 8280–8288. [Google Scholar] [CrossRef]
- Wang, J.C. DNA supercoiling and its effects on the structure of DNA. J. Cell Sci. 1984, 1, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, N.; Droffner, M.L. Mechanisms Determining Aerobic or Anaerobic Growth in the Facultative Anaerobe Salmonella-Typhimurium. Proc. Natl. Acad. Sci. USA 1985, 82, 2077–2081. [Google Scholar] [CrossRef] [Green Version]
- Higgins, C.F.; Dorman, C.J.; Stirling, D.A.; Waddell, L.; Booth, I.R.; May, G.; Bremer, E. A physiological role for DNA supercoiling in the osmotic regulation of gene expression in S. typhimurium and E. coli. Cell 1988, 52, 569–584. [Google Scholar] [CrossRef]
- Wu, H.-Y.; Shyy, S.; Wang, J.C.; Liu, L. Transcription Generates Positively and Negatively Supercoiled Domains in the Template. Cell 1988, 53, 433–440. [Google Scholar] [CrossRef]
- McClellan, J.A.; Boublikova, P.; Palecek, E.; Lilley, D.M. Superhelical Torsion in Cellular DNA Responds Directly to Environmental and Genetic-Factors. Proc. Natl. Acad. Sci. USA 1990, 87, 8373–8377. [Google Scholar] [CrossRef] [Green Version]
- Rahmouni, A.; Wells, R. Stabilization of Z DNA in vivo by localized supercoiling. Science 1989, 246, 358–363. [Google Scholar] [CrossRef]
- Rahmouni, A.R.; Wells, R.D. Direct evidence for the effect of transcription on local DNA supercoiling in vivo. J. Mol. Biol. 1992, 223, 131–144. [Google Scholar] [CrossRef]
- Holck, A.; Kleppe, K. Affinity of Protein Hu for Different Nucleic-Acids. FEBS Lett. 1985, 185, 121–124. [Google Scholar] [CrossRef] [Green Version]
- Shindo, H.; Furubayashi, A.; Shimizu, M.; Miyake, M.; Lmamoto, F. Preferential Binding of Escherichia coli Histone-Like Protein Hu-Alpha to Negatively Supercoiled DNA. Nucleic Acids Res. 1992, 20, 1553–1558. [Google Scholar] [CrossRef]
- McMacken, R. A Bipartite Signaling Mechanism Involved in DnaJ-mediated Activation of the Escherichia coli DnaK Protein. J. Biol. Chem. 1996, 271, 11236. [Google Scholar]
- Wold, M.; Mallory, J.B.; Roberts, J.D.; LeBowitz, J.H.; McMacken, R. Initiation of bacteriophage lambda DNA replication in vitro with purified lambda replication proteins. Proc. Natl. Acad. Sci. USA 1982, 79, 6176–6180. [Google Scholar] [CrossRef] [Green Version]
- Green, M.R.; Sambrook, J. Molecular Cloning, a Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2012; pp. 11–14. [Google Scholar]
- Clark, D.J.; Felsenfeld, G. Formation of Nucleosomes on Positively Supercoiled DNA. EMBO J. 1991, 10, 387–395. [Google Scholar] [CrossRef]
- Keller, W. Determination of the number of superhelical turns in simian virus 40 DNA by gel electrophoresis. Proc. Natl. Acad. Sci. USA 1975, 72, 4876–4880. [Google Scholar] [CrossRef] [Green Version]
- Carey, J. Gel Retardation at Low Ph Resolves Trp Repressor-DNA Complexes for Quantitative Study. Proc. Natl. Acad. Sci. USA 1988, 85, 975–979. [Google Scholar] [CrossRef] [Green Version]
- Sinden, R.R.; Carlson, J.O.; Pettijohn, D.E. Torsional tension in the DNA double helix measured with trimethylpsoralen in living E. coli cells: Analogous measurements in insect and human cells. Cell 1980, 21, 773–783. [Google Scholar] [CrossRef]
- Bliska, J.B.; Cozzarelli, N.R. Use of site-specific recombination as a probe of DNA structure and metabolism in vivo. J. Mol. Biol. 1987, 194, 205–218. [Google Scholar] [CrossRef]
- Zheng, G.; Kochel, T.; Hoepfner, R.W.; Timmons, S.E.; Sinden, R.R. Torsionally Tuned Cruciform and Z-DNA Probes for Measuring Unrestrained Supercoiling at Specific Sites in DNA of Living Cells. J. Mol. Biol. 1991, 221, 107–129. [Google Scholar] [CrossRef]
- Drlica, K.; Rouviereyaniv, J. Histone-Like Proteins of Bacteria. Microbiol. Rev. 1987, 51, 301–319. [Google Scholar] [CrossRef]
- Anuchin, A.M.; Goncharenko, A.V.; Demidenok, O.I.; Kaprelyants, A.S. Histone-Like Proteins of Bacteria (Review). Appl. Biochem. Microbiol. 2011, 47, 580–585. [Google Scholar] [CrossRef]
- Dixon, N.E.; Kornberg, A. Protein-Hu in the Enzymatic Replication of the Chromosomal Origin of Escherichia coli. Proc. Natl. Acad. Sci. USA 1984, 81, 424–428. [Google Scholar] [CrossRef] [Green Version]
- Grabarek, Z.; Gergely, J. Zero-Length Crosslinking Procedure with the Use of Active Esters. Anal. Biochem. 1990, 185, 131–135. [Google Scholar] [CrossRef]
- Losso, M.A.; Pawlik, R.T.; Canonaco, M.A.; Gualerzi, C.O. Proteins from the prokaryotic nucleoid. A protein-protein cross-linking study on the quaternary structure of Escherichia coli DNA-binding protein NS (HU). FEBS J. 1986, 155, 27–32. [Google Scholar] [CrossRef]
- Kobryn, K.; Lavoie, B.D.; Chaconas, G. Supercoiling-dependent site-specific binding of HU to naked mu DNA. J. Mol. Biol. 1999, 289, 777–784. [Google Scholar] [CrossRef]
- Haniford, D.B.; Pulleyblank, D.E. Transition of a cloned d(AT)n-d(AT)n tract to a cruciform in vivo. Nucleic Acids Res. 1985, 13, 4343–4363. [Google Scholar] [CrossRef] [Green Version]
- Schneider, R.; Lurz, R.; Lüder, G.; Tolksdorf, C.; Travers, A.; Muskhelishvili, G. An architectural role of the Escherichia coli chromatin protein FIS in organising DNA. Nucleic Acids Res. 2001, 29, 5107–5114. [Google Scholar] [CrossRef] [Green Version]
- Tupper, A.; Owen-Hughes, T.; Ussery, D.; Santos, D.; Ferguson, D.; Sidebotham, J.; Hinton, J.; Higgins, C. The Chromatin-Associated Protein H-Ns Alters DNA Topology in-Vitro. EMBO J. 1994, 13, 258–268. [Google Scholar] [CrossRef]
- Malik, M.; Bensaid, A.; Rouviere-Yaniv, J.; Drlica, K. Histone-like protein HU and bacterial DNA topology: Suppression of an HU deficiency by gyrase mutations. J. Mol. Biol. 1996, 256, 66–76. [Google Scholar] [CrossRef]
- Bensaid, A.; Almeida, A.; Drlica, K.; Rouviere-Yaniv, J. Cross-talk between topoisomerase I and HU in Escherichia coli. J. Mol. Biol. 1996, 256, 292–300. [Google Scholar] [CrossRef]
- Voulgarakis, N.K. A stochastic reaction-diffusion model for protein aggregation on DNA. Int. J. Mod. Phys. C 2017, 28, 1750102. [Google Scholar] [CrossRef] [Green Version]
- Berthold, V.; Geider, K. Interaction of DNA with DNA-binding proteins. The characterization of protein HD from Escherichia coli and its nucleic acid complexes. FEBS J. 2010, 71, 443–449. [Google Scholar] [CrossRef]
- Paci, M.; Pon, C.L.; Losso, M.A.; Gualerzi, C. Proteins from the prokaryotic nucleoid. High-resolution 1H NMR spectroscopic study of Escherichia coli DNA-binding proteins NS1 and NS2. FEBS J. 2010, 138, 193–200. [Google Scholar] [CrossRef]
- Suryanarayana, T.; Subramanian, A.R. Specific association of two homologous DNA-binding proteins to the native 30-S ribosomal subunits of Escherichia coli. Biochim. Biophys. Acta (BBA)-Nucleic Acids Protein Synth. 1978, 520, 342–357. [Google Scholar] [CrossRef]
- White, S.W.; Appelt, K.; Wilson, K.S.; Tanaka, I. A protein structural motif that bends DNA. Proteins 1989, 5, 281–288. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, L.; Zhang, Z.; McMacken, R. Interaction of the Escherichia coli HU Protein with Various Topological Forms of DNA. Biomolecules 2021, 11, 1724. https://doi.org/10.3390/biom11111724
Huang L, Zhang Z, McMacken R. Interaction of the Escherichia coli HU Protein with Various Topological Forms of DNA. Biomolecules. 2021; 11(11):1724. https://doi.org/10.3390/biom11111724
Chicago/Turabian StyleHuang, Li, Zhenfeng Zhang, and Roger McMacken. 2021. "Interaction of the Escherichia coli HU Protein with Various Topological Forms of DNA" Biomolecules 11, no. 11: 1724. https://doi.org/10.3390/biom11111724