
Novel Methylation Biomarkers for Colorectal Cancer Prognosis



Abstract
:1. Introduction
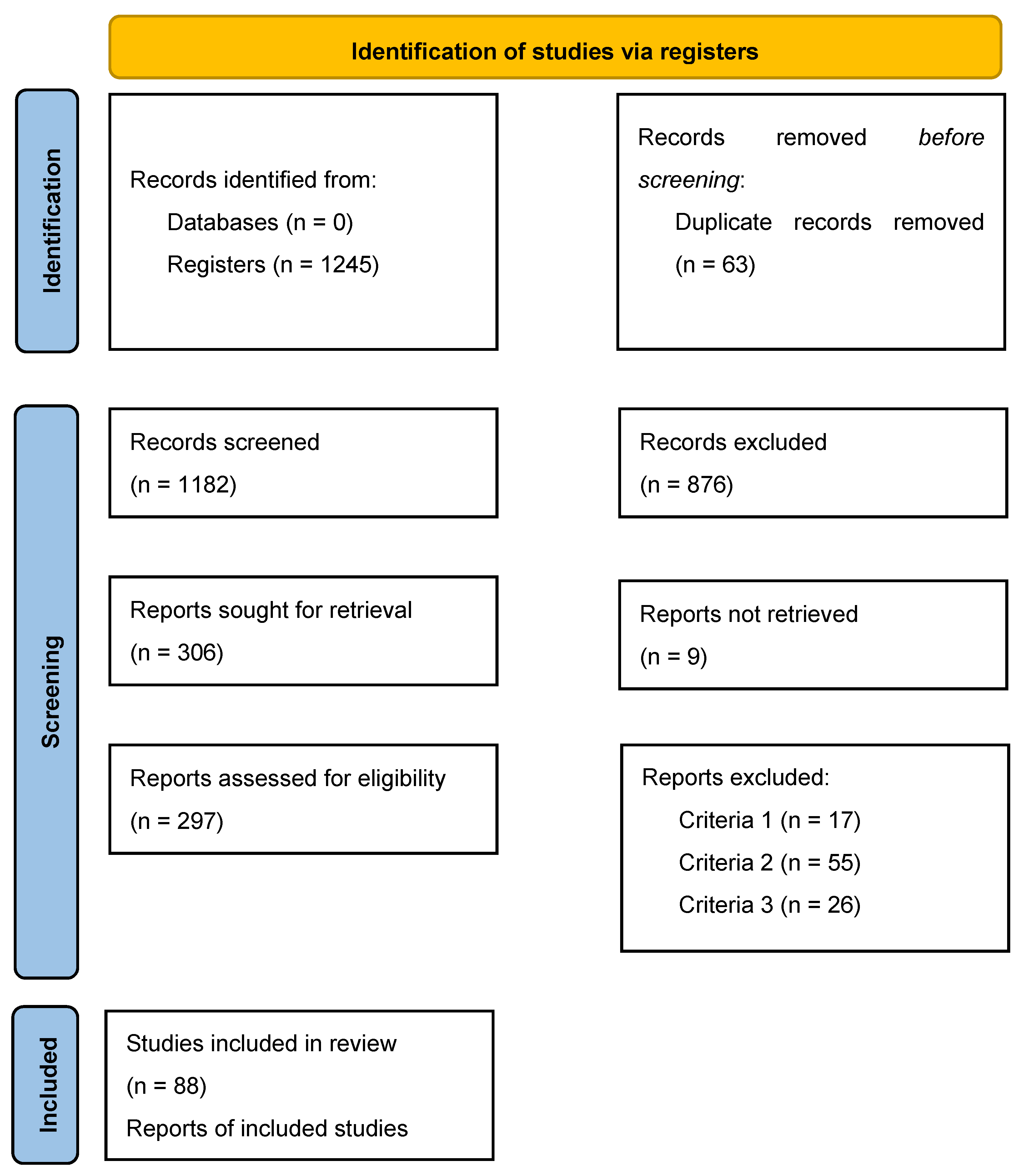
2. Search Strategy
3. Insights into Colorectal Carcinogenesis
4. From CRC to Metastasis: A Complex Journey
5. Chromosome Stability and DNA Methylation Profiles: Can Epigenetic Changes Drive Metastasis?
6. Novel Methylation Biomarkers in CRC and Metastasis Prognosis
6.1. Microbiome and DNA Methylation as Biomarkers of Colorectal Cancer
6.2. Inhibin Subunit Beta B (INHBB)
6.3. SPARC-Related Modular Calcium-Binding Protein 2 (SMOC2)
6.4. Helicase-like Transcription Factor (HLTF)
6.5. Eye Absent Homolog 4 (EYA4)
6.6. GATA-Binding Protein 5 (GATA5)
6.7. Brain-Derived Neurotrophic Factor (BDNF)
6.8. Transforming Growth Factor Beta Regulator 4 (TBRG4)
6.9. Open Issues Regarding the Use of Methylation Biomarkers
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA. Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Borgna, V.; Villegas, J.; Burzio, V.A.; Belmar, S.; Araya, M.; Jeldes, E.; Lobos-González, L.; Silva, V.; Villota, C.; Oliveira-Cruz, L.; et al. Mitochondrial ASncmtRNA-1 and ASncmtRNA-2 as Potent Targets to Inhibit tumor Growth and Metastasis in the RenCa murine Renal Adenocarcinoma Model. Oncotarget 2017, 8, 43692–43708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanley, M.P.; Hahn, M.A.; Li, A.X.; Wu, X.; Lin, J.; Wang, J.; Choi, A.H.; Ouyang, Z.; Fong, Y.; Pfeifer, G.P.; et al. Genome-wide DNA methylation profiling reveals cancer-associated changes within early colonic neoplasia. Oncogene 2017, 36, 5035–5044. [Google Scholar] [CrossRef] [Green Version]
- Slattery, M.L. Diet, lifestyle, and colon cancer. Semin. Gastrointest. Dis. 2000, 11, 142–146. [Google Scholar]
- Aran, V.; Victorino, A.P.; Thuler, L.C.; Ferreira, C.G. Colorectal Cancer: Epidemiology, Disease Mechanisms and Interventions to Reduce Onset and Mortality. Clin. Colorectal Cancer 2016, 15, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Burt, R.; Neklason, D.W. Genetic testing for inherited colon cancer. Gastroenterology 2005, 128, 1696–1716. [Google Scholar] [CrossRef] [PubMed]
- Fearon, E.R. Molecular genetics of colorectal cancer. Annu. Rev. Pathol. Mech. Dis. 2011, 6, 479–507. [Google Scholar] [CrossRef]
- Fearon, E.R.; Vogelstein, B. A genetic model for colorectal tumorigenesis. Cell 1990, 61, 759–767. [Google Scholar] [CrossRef]
- Tauriello, D.V.F.; Calon, A.; Lonardo, E.; Batlle, E. Determinants of metastatic competency in colorectal cancer. Mol. Oncol. 2017, 11, 97–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manfredi, S.; Lepage, C.; Hatem, C.; Coatmeur, O.; Faivre, J.; Bouvier, A.M. Epidemiology and management of liver metastases from colorectal cancer. Ann. Surg. 2006, 244, 254–259. [Google Scholar] [CrossRef]
- Brenner, H.; Kloor, M.; Pox, C.P. Colorectal cancer. Lancet 2014, 383, 1490–1502. [Google Scholar] [CrossRef]
- Huang, D.; Sun, W.; Zhou, Y.; Li, P.; Chen, F.; Chen, H.; Xia, D.; Xu, E.; Lai, M.; Wu, Y.; et al. Mutations of key driver genes in colorectal cancer progression and metastasis. Cancer Metastasis Rev. 2018, 37, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Schiffman, J.D.; Fisher, P.G.; Gibbs, P. Early Detection of Cancer: Past, Present, and Future. Introduction to cancer screening and Tumor Markers for Early Cancer Detection. ASCO Educ. B 2015, 35, 57–65. [Google Scholar]
- Wu, L.; Qu, X. Cancer biomarker detection: Recent achievements and challenges. Chem. Soc. Rev. 2015, 44, 2963–2997. [Google Scholar] [CrossRef] [PubMed]
- Lech, G.; Słotwiński, R.; Słodkowski, M.; Krasnodębski, I.W. Colorectal cancer tumour markers and biomarkers: Recent therapeutic advances. World J. Gastroenterol. 2016, 22, 1745–1755. [Google Scholar] [CrossRef]
- Zlobec, I. Novel biomarkers for the prediction of metastasis in colorectal cancer. Expert Opin. Med. Diagn. 2013, 7, 137–146. [Google Scholar] [CrossRef]
- Sartor, O.; de Bono, J.S. Metastatic Prostate Cancer. N. Engl. J. Med. 2018, 378, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Lou, J. DNA Methylation Analysis. Methods Mol. Biol. 2019, 1894, 181–227. [Google Scholar]
- Zeng, H.; He, B.; Yi, C.; Peng, J. Liquid biopsies: DNA methylation analyses in circulating cell-free DNA. J. Genet. Genom. 2018, 45, 185–192. [Google Scholar] [CrossRef]
- Muzny, D.M.; Bainbridge, M.N.; Chang, K.; Dinh, H.H.; Drummond, J.A.; Fowler, G.; Kovar, C.L.; Lewis, L.R.; Morgan, M.B.; Newsham, I.F.; et al. Comprehensive molecular characterization of human colon and rectal cancer. Nature 2012, 487, 330–337. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.H.; Goel, A.; Chung, D.C.; Hospital, M.G.; Unit, T.E.; Hospital, M.G.; Sammons, C.A.; Hospital, M.G.; Medical, H. Pathways of Colorectal Carcinogenesis. Gastroenterology 2020, 158, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Zhan, T.; Rindtorff, N.; Boutros, M. Wnt signaling in cancer. Oncogene 2017, 36, 1461–1473. [Google Scholar] [CrossRef]
- Cheng, X.; Xu, X.; Chen, D.; Zhao, F.; Wang, W. Therapeutic potential of targeting the Wnt/β-catenin signaling pathway in colorectal cancer. Biomed. Pharmacother. 2019, 110, 473–481. [Google Scholar] [CrossRef]
- Guo, Y.; Pan, W.; Liu, S.; Shen, Z.; Xu, Y.; Hu, L. ERK/MAPK signalling pathway and tumorigenesis (Review). Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauen, K.A. The RASopathies. Annu. Rev. Genomics Hum. Genet 2013, 14, 355–369. [Google Scholar] [CrossRef] [Green Version]
- Joerger, A.C.; Fersht, A.R. The p53 Pathway: Origins, Inactivation in Cancer, and Emerging Therapeutic Approaches. Annu. Rev. Biochem. 2016, 85, 375–404. [Google Scholar] [CrossRef]
- Mantovani, F.; Collavin, L.; Del Sal, G. Mutant p53 as a guardian of the cancer cell. Cell Death Differ. 2019, 26, 199–212. [Google Scholar] [CrossRef]
- Essmann, F.; Schulze-Osthoff, K. Translational approaches targeting the p53 pathway for anti-cancer therapy. Br. J. Pharmacol. 2012, 165, 328–344. [Google Scholar] [CrossRef] [PubMed]
- Vogelstein, B.; Lane, D.; Levine, A.J. Surfing the p53 network. Nature 2000, 408, 307–310. Available online: http://www.nature.com/nature/journal/v408/n6810/full/408307a0.html#B41 (accessed on 2 August 2021). [CrossRef]
- Levine, A.J. P53: 800 Million Years of Evolution and 40 Years of Discovery. Nat. Rev. Cancer 2020, 20, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; Yang, J.; Murphy, R.F.; Agrawal, D.K. Regulation of the p14ARF-Mdm2-p53 pathway: An overview in breast cancer. Exp. Mol. Pathol. 2006, 81, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Itatani, Y.; Kawada, K.; Sakai, Y. Transforming growth factor-β signaling pathway in colorectal cancer and its tumor microenvironment. Int. J. Mol. Sci. 2019, 20, 5822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syed, V. TGF-β Signaling in Cancer. J. Cell. Biochem. 2016, 117, 1279–1287. [Google Scholar] [CrossRef]
- Xie, F.; Ling, L.; Van Dam, H.; Zhou, F.; Zhang, L. TGF-β signaling in cancer metastasis. Acta Biochim. Biophys. Sin. 2018, 50, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Colak, S.; ten Dijke, P. Targeting TGF-β Signaling in Cancer. Trends Cancer 2017, 3, 56–71. [Google Scholar] [CrossRef]
- Tu, S.; Huang, W.; Huang, C.; Luo, Z.; Yan, X. Contextual Regulation of TGF-β Signaling in Liver Cancer. Cells 2019, 8, 1235. [Google Scholar] [CrossRef] [Green Version]
- Haque, S.; Morris, J.C. Transforming growth factor-β: A therapeutic target for cancer. Hum. Vaccines Immunother. 2017, 13, 1741–1750. [Google Scholar] [CrossRef]
- Gu, S.; Feng, X.H. TGF-β signaling in cancer. Acta Biochim. Biophys. Sin. 2018, 50, 941–949. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Mishra, L.; Deng, C.X. The role of TGF-β/SMAD4 signaling in cancer. Int. J. Biol. Sci. 2018, 14, 111–123. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Han, F.; Liu, W.; Chen, H.; Hao, X.; Jiang, X.; Yin, L.; Huang, Y.; Cao, J.; Zhang, H.; et al. ALX4, an epigenetically down regulated tumor suppressor, inhibits breast cancer progression by interfering Wnt/β-catenin pathway. J. Exp. Clin. Cancer Res. 2017, 36, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nassar, F.J.; Msheik, Z.S.; Nasr, R.R.; Temraz, S.N. Methylated circulating tumor DNA as a biomarker for colorectal cancer diagnosis, prognosis, and prediction. Clin. Epigenetics 2021, 13, 111. [Google Scholar] [CrossRef]
- Barchitta, M.; Maugeri, A.; Li Destri, G.; Basile, G.; Agodi, A. Epigenetic biomarkers in colorectal cancer patients receiving adjuvant or neoadjuvant therapy: A systematic review of epidemiological studies. Int. J. Mol. Sci. 2019, 20, 3842. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Su, X.; Long, Y.; Zhou, D.; Zhang, X.; Ye, Z.; Ma, J.; Tang, T.; Wang, F.; He, C. A systematic review and quantitative assessment of methylation biomarkers in fecal DNA and colorectal cancer and its precursor, colorectal adenoma. Mutat. Res.—Rev. Mutat. Res. 2019, 779, 45–57. [Google Scholar] [CrossRef]
- Petit, J.; Carroll, G.; Gould, T.; Pockney, P.; Dun, M.; Scott, R.J. Cell-free DNA as a Diagnostic Blood-Based Biomarker for Colorectal Cancer: A Systematic Review. J. Surg. Res. 2019, 236, 184–197. [Google Scholar] [CrossRef]
- Vacante, M.; Borzì, A.M.; Basile, F.; Biondi, A. Biomarkers in colorectal cancer: Current clinical utility and future perspectives. World J. Clin. Cases 2018, 6, 869–881. [Google Scholar] [CrossRef]
- Rasmussen, S.L.; Krarup, H.B.; Sunesen, K.G.; Johansen, M.B.; Stender, M.T.; Pedersen, I.S.; Madsen, P.H.; Thorlacius-Ussing, O. Hypermethylated DNA, a circulating biomarker for colorectal cancer detection. PLoS ONE 2017, 12, e0180809. [Google Scholar] [CrossRef] [Green Version]
- Salehi, R.; Atapour, N.; Vatandoust, N.; Farahani, N.; Ahangari, F.; Salehi, A. Methylation pattern of ALX4 gene promoter as a potential biomarker for blood-based early detection of colorectal cancer. Adv. Biomed. Res. 2015, 4, 252. [Google Scholar] [CrossRef] [PubMed]
- Laugsand, E.A.; Brenne, S.S.; Skorpen, F. DNA methylation markers detected in blood, stool, urine, and tissue in colorectal cancer: A systematic review of paired samples. Int. J. Colorectal Dis. 2021, 36, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.X.; Luo, G.H. DNA methylation detection methods used in colorectal cancer. World J. Clin. Cases 2019, 7, 2916–2929. [Google Scholar] [CrossRef] [PubMed]
- Jedi, M.; Young, G.P.; Pedersen, S.K.; Symonds, E.L. Methylation and Gene Expression of BCAT1 and IKZF1 in Colorectal Cancer Tissues. Clin. Med. Insights Oncol. 2018, 12, 1179554918775064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, J.; Liu, X.; Qi, Y.; Niu, F.; Niu, Z.; Geng, W.; Zou, Z.; Huang, R.; Wang, J.; Zou, H. BMP3 suppresses colon tumorigenesis via ActRIIB/SMAD2-dependent and TAK1/JNK signaling pathways. J. Exp. Clin. Cancer Res. 2019, 38, 428. [Google Scholar] [CrossRef] [PubMed]
- Jung, G.; Moreira, L.; Balaguer, F.; Goel, A.; Sammons, C.A.; Therapeutics, E. Epigenetics of colorectal cancer: Biomarker and therapeutic potential. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 111–130. [Google Scholar] [CrossRef] [PubMed]
- Gu, F.; Yuan, S.; Liu, L.; Zhu, P.; Yang, Y.; Pan, Z.; Zhou, W. EYA4 serves as a prognostic biomarker in hepatocellular carcinoma and suppresses tumour angiogenesis and metastasis. J. Cell. Mol. Med. 2019, 23, 4208–4216. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Williams, M.; Cheng, Y.Y.; Leung, W.K. Roles of methylated DNA biomarkers in patients with colorectal cancer. Dis. Markers 2019, 2019, 2673543. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Zhou, T.F.; Qiu, B.A.; Yang, Y.X.; Zhu, Y.J.; An, Y.; Zhao, W.C.; Wu, Y.T.; Ma, P.F.; Li, J.B.; et al. Methylation-Mediated Silencing of GATA5 Gene Suppresses Cholangiocarcinoma Cell Proliferation and Metastasis. Transl. Oncol. 2018, 11, 585–592. [Google Scholar] [CrossRef]
- Raut, J.R.; Guan, Z.; Schrotz-King, P.; Brenner, H. Fecal DNA methylation markers for detecting stages of colorectal cancer and its precursors: A systematic review. Clin. Epigenetics 2020, 12, 122. [Google Scholar] [CrossRef]
- Hu, B.; Zhang, K.; Li, S.; Li, H.; Yan, Z.; Huang, L.; Wu, J.; Han, X.; Jiang, W.; Mulatibieke, T.; et al. HIC1 attenuates invasion and metastasis by inhibiting the IL-6/STAT3 signalling pathway in human pancreatic cancer. Cancer Lett. 2016, 376, 387–398. [Google Scholar] [CrossRef]
- Liu, L.; Liu, H.; Zhou, Y.; He, J.; Liu, Q.; Wang, J.; Zeng, M.; Yuan, D.; Tan, F.; Zhou, Y.; et al. HLTF suppresses the migration and invasion of colorectal cancer cells via TGF-β/SMAD signaling in vitro. Int. J. Oncol. 2018, 53, 2780–2788. [Google Scholar] [CrossRef] [Green Version]
- Wallner, M.; Herbst, A.; Behrens, A.; Crispin, A.; Stieber, P.; Göke, B.; Lamerz, R.; Kolligs, F.T. Methylation of serum DNA is an independent prognostic marker in colorectal cancer. Clin. Cancer Res. 2006, 12, 7347–7352. [Google Scholar] [CrossRef] [Green Version]
- Cai, Q.; Dozmorov, M.; Oh, Y. IGFBP-3/IGFBP-3 Receptor System as an Anti-Tumor and Anti-Metastatic Signaling in Cancer. Cells 2020, 9, 1261. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, P.; Pandey, A.; Gosipatala, S.B. IGFBP3 gene promoter methylation analysis and its association with clinicopathological characteristics of colorectal carcinoma. Mol. Biol. Rep. 2020, 47, 6919–6927. [Google Scholar] [CrossRef]
- Lam, K.; Pan, K.; Linnekamp, J.F.; Medema, J.P.; Kandimalla, R. DNA methylation based biomarkers in colorectal cancer: A systematic review. Biochim. Biophys. Acta—Rev. Cancer 2016, 1866, 106–120. [Google Scholar] [CrossRef] [PubMed]
- Javierre, B.M.; Rodriguez-Ubreva, J.; Al-Shahrour, F.; Corominas, M.; Graña, O.; Ciudad, L.; Agirre, X.; Pisano, D.G.; Valencia, A.; Roman-Gomez, J.; et al. Long-range epigenetic silencing associates with deregulation of Ikaros targets in colorectal cancer cells. Mol. Cancer Res. 2011, 9, 1139–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Xu, J.; Wu, S.; Wang, R.; Qu, X.; Yu, W.; Li, J.; Chen, L. IRF4 promotes cell proliferation by JNK pathway in multiple myeloma. Med. Oncol. 2013, 30, 594. [Google Scholar] [CrossRef] [PubMed]
- Baiula, M.; Spampinato, S.; Gentilucci, L.; Tolomelli, A. Novel Ligands Targeting α4β1 Integrin: Therapeutic Applications and Perspectives. Front. Chem. 2019, 7, 489. [Google Scholar] [CrossRef]
- Kerachian, M.A.; Kerachian, M. Long interspersed nucleotide element-1 (LINE-1) methylation in colorectal cancer. Clin. Chim. Acta 2019, 488, 209–214. [Google Scholar] [CrossRef]
- Lou, Y.T.; Chen, C.W.; Fan, Y.C.; Chang, W.C.; Lu, C.Y.; Wu, I.C.; Hsu, W.H.; Huang, C.W.; Wang, J.Y. LINE-1 methylation status correlates significantly to post-therapeutic recurrence in stage III colon cancer patients receiving FOLFOX-4 adjuvant chemotherapy. PLoS ONE 2015, 10, e0123973. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Ren, B.; Gui, Q.; Zhao, J.; Wu, M.; Shen, M.; Li, D.; Li, D.; Chen, K.; Tao, M.; et al. Blocking MAPK/ERK pathway sensitizes hepatocellular carcinoma cells to temozolomide via downregulating MGMT expression. Ann. Transl. Med. 2020, 8, 1305. [Google Scholar] [CrossRef]
- Li, X.; Wang, Y.; Zhang, Z.; Yao, X.; Ge, J.; Zhao, Y. Correlation of MLH1 and MGMT methylation levels between peripheral blood leukocytes and colorectal tissue DNA samples in colorectal cancer patients. Oncol. Lett. 2013, 6, 1370–1376. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.Y.; Yang, Q.F.; Zeng, Y.L.; Liu, Y.Y.; Liu, Y.T.; Gu, F.F.; Hu, Y.; Zhang, K.; Zhong, H.; Liu, L. Clinical value of MLH1-negative circulating tumor cells in lung cancer patients. Medicine 2019, 98, e15721. [Google Scholar] [CrossRef]
- Tse, J.W.T.; Jenkins, L.J.; Chionh, F.; Mariadason, J.M. Aberrant DNA Methylation in Colorectal Cancer: What Should We Target? Trends Cancer 2017, 3, 698–712. [Google Scholar] [CrossRef] [PubMed]
- Constâncio, V.; Nunes, S.P.; Henrique, R.; Jerónimo, C. DNA Methylation-Based Testing in Liquid Biopsies as Detection and Prognostic Biomarkers for the Four Major Cancer Types. Cells 2020, 9, 624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; O’Rourke, D.; Sanchez-Garcia, J.F.; Cai, T.; Scheuenpflug, J.; Feng, Z. Development of a liquid biopsy based purely quantitative digital droplet PCR assay for detection of MLH-1promoter methylation. BMC Cancer 2020, 21. [Google Scholar] [CrossRef]
- Zheng, J.; Li, Y.; Zhu, S.; Li, J.; Zhao, Q.; Ji, G.; Wang, W.; Chu, D. NDRG4 stratifies the prognostic value of body mass index in colorectal cancer. Oncotarget 2016, 7, 1311–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Li, Z.; Wang, C.; Pan, T.; Chang, X.; Wang, X.; Zhou, Q.; Wu, X.; Li, J.; Zhang, J.; et al. Oncostatin M receptor, positively regulated by SP1, promotes gastric cancer growth and metastasis upon treatment with Oncostatin M. Gastric Cancer 2019, 22, 955–966. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Besch-Williford, C.; Hyder, S.M. PRIMA-1 inhibits growth of breast cancer cells by re-activating mutant p53 protein. Int. J. Oncol. 2009, 35, 1015–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malpeli, G.; Innamorati, G.; Decimo, I.; Bencivenga, M.; Kamdje, A.H.N.; Perris, R.; Bassi, C. Methylation dynamics of RASSF1A and its impact on cancer. Cancers 2019, 11, 959. [Google Scholar] [CrossRef] [Green Version]
- Matthaios, D.; Balgkouranidou, I.; Karayiannakis, A.; Bolanaki, H.; Xenidis, N.; Amarantidis, K.; Chelis, L.; Romanidis, K.; Chatzaki, A.; Lianidou, E.; et al. Methylation status of the APC and RASSF1A promoter in cell-free circulating DNA and its prognostic role in patients with colorectal cancer. Oncol. Lett. 2016, 12, 748–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, R.; Yu, J.; Yan, X.; Ni, Q.; Zhi, X.; Li, X.; Jiang, B.; Zhu, J. Syndecan-2 in colorectal cancer plays oncogenic role via epithelial-mesenchymal transition and MAPK pathway. Biomed. Pharmacother. 2020, 121, 109630. [Google Scholar] [CrossRef]
- Su, W.C.; Kao, W.Y.; Chang, T.K.; Tsai, H.L.; Huang, C.W.; Chen, Y.C.; Li, C.C.; Hsieh, Y.C.; Yeh, H.J.; Chang, C.C.; et al. Stool DNA test targeting methylated syndecan-2 (SDC2) as a noninvasive screening method for colorectal cancer. Biosci. Rep. 2021, 41, BSR20201930. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Cao, Y.; Liu, L.; Zhao, J.; Zhang, T.; Xiao, L.; Jia, M.; Tian, Q.; Yu, H.; Chen, S.; et al. SEPT9_i1 regulates human breast cancer cell motility through cytoskeletal and RhoA/FAK signaling pathway regulation. Cell Death Dis. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tham, C.; Chew, M.; Soong, R.; Lim, J.; Ang, M.; Tang, C.; Zhao, Y.; Ong, S.Y.K.; Liu, Y. Postoperative serum methylation levels of TAC1 and SEPT9 are independent predictors of recurrence and survival of patients with colorectal cancer. Cancer 2014, 120, 3131–3141. [Google Scholar] [CrossRef]
- Majchrzak-Celińska, A.; Słocińska, M.; Barciszewska, A.M.; Nowak, S.; Baer-Dubowska, W. Wnt pathway antagonists, SFRP1, SFRP2, SOX17, and PPP2R2B, are methylated in gliomas and SFRP1 methylation predicts shorter survival. J. Appl. Genet. 2016, 57, 189–197. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Qiao, J.; He, H.; Song, J.; Zhao, S.; Yu, J. TFPI2 suppresses breast cancer progression through inhibiting TWIST-integrin α5 pathway. Mol. Med. 2020, 26, 27. [Google Scholar] [CrossRef] [PubMed]
- Satelli, A.; Li, S. Vimentin as a potential molecular target in cancer therapy Or Vimentin, an overview and its potential as a molecular target for cancer therapy. Cell Mol. Life Sci. 2011, 68, 3033–3046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Xie, A.; Cao, Q.; Li, X.; Chen, J. INHBB Is a Novel Prognostic Biomarker Associated with Cancer-Promoting Pathways in Colorectal Cancer. BioMed Res. Int. 2020, 2020, 6909672. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Yu, Y.; Song, Y.; Li, X.; Lan, D.; Zhang, P.; Xiao, Y.; Xing, Y. Activation of BDNF/TrkB pathway promotes prostate cancer progression via induction of epithelial-mesenchymal transition and anoikis resistance. FASEB J. 2020, 34, 9087–9101. [Google Scholar] [CrossRef]
- Ili, C.; Buchegger, K.; Demond, H.; Castillo-Fernandez, J.; Kelsey, G.; Zanella, L.; Abanto, M.; Riquelme, I.; López, J.; Viscarra, T.; et al. Landscape of genome-wide dna methylation of colorectal cancer metastasis. Cancers 2020, 12, 2710. [Google Scholar] [CrossRef]
- Jang, B.G.; Kim, H.S.; Bae, J.M.; Kim, W.H.; Kim, H.U.; Kang, G.H. SMOC2, an intestinal stem cell marker, is an independent prognostic marker associated with better survival in colorectal cancers. Sci. Rep. 2020, 10, 14591. [Google Scholar] [CrossRef]
- Yuan, L.; Guo, F.; Wang, L.; Zou, Q. Prediction of tumor metastasis from sequencing data in the era of genome sequencing. Brief. Funct. Genom. 2019, 18, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.X.; Bos, P.D.; Massagué, J. Metastasis: From dissemination to organ-specific colonization. Nat. Rev. Cancer 2009, 9, 274–284. [Google Scholar] [CrossRef]
- Wan, L.; Pantel, K.; Kang, Y. Tumor metastasis: Moving new biological insights into the clinic. Nat. Med. 2013, 19, 1450–1464. [Google Scholar] [CrossRef]
- Fidler, I.J. The pathogenesis of cancer metastasis: The “seed and soil” hyphotesis revisited. Nat. Rev. Cancer 2003, 3, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.; Haider, A.; Rashid, S.; Al-Nabet, A.D.M.H. Paget’s “seed and Soil” Theory of Cancer Metastasis: An Idea Whose Time has Come. Adv. Anat. Pathol. 2019, 26, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Seretis, F.; Seretis, C.; Youssef, H.; Chapman, M. Colorectal cancer: Seed and soil hypothesis revisited. Anticancer Res. 2014, 34, 2087–2094. [Google Scholar] [PubMed]
- Fokas, E.; Engenhart-Cabillic, R.; Daniilidis, K.; Rose, F.; An, H.X. Metastasis: The seed and soil theory gains identity. Cancer Metastasis Rev. 2007, 26, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Weinberg, R.A. Epithelial-to-mesenchymal transition in cancer: Complexity and opportunities. Front. Med. 2018, 12, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.; Xu, E.; Liu, H.; Wan, L.; Lai, M. Epithelial-mesenchymal transition in colorectal cancer metastasis: A system review. Pathol. Res. Pract. 2015, 211, 557–569. [Google Scholar] [CrossRef]
- Teeuwssen, M.; Fodde, R. Wnt Signaling in Ovarian Cancer Stemness, EMT, and Therapy Resistance. J. Clin. Med. 2019, 8, 1658. [Google Scholar] [CrossRef] [Green Version]
- Basu, S.; Cheriyamundath, S.; Ben-Ze’ev, A. Cell–cell adhesion: Linking wnt/β-catenin signaling with partial emt and stemness traits in tumorigenesis [version 1; peer review: 4 approved]. F1000Research 2018, 7, 1488. [Google Scholar] [CrossRef] [Green Version]
- Vincan, E.; Barker, N. The upstream components of the Wnt signalling pathway in the dynamic EMT and MET associated with colorectal cancer progression. Clin. Exp. Metastasis 2008, 25, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Baker, D.; Dijke, P. Ten TGF-β-mediated epithelial-mesenchymal transition and cancer metastasis. Int. J. Mol. Sci. 2019, 20, 2767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villalba, M.; Evans, S.R.; Vidal-Vanaclocha, F.; Calvo, A. Role of TGF-β in metastatic colon cancer: It is finally time for targeted therapy. Cell Tissue Res. 2017, 370, 29–39. [Google Scholar] [CrossRef]
- Seoane, J.; Gomis, R.R. TGF-B Family Signaling in Tumor Suppression and Cancer Progression. Cold Spring Harb. Perspect. Biol. 2017, 9, a022277. [Google Scholar] [CrossRef] [Green Version]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanism of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [Green Version]
- Castro-Giner, F.; Aceto, N. Tracking cancer progression: From circulating tumor cells to metastasis. Genome Med. 2020, 12, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, C.; Yang, C.; Wang, S.; Shi, D.; Zhang, C.; Lin, X.; Liu, Q.; Dou, R.; Xiong, B. Crosstalk between cancer cells and tumor associated macrophages is required for mesenchymal circulating tumor cell-mediated colorectal cancer metastasis. Mol. Cancer 2019, 18, 64. [Google Scholar] [CrossRef]
- Park, H.A.; Brown, S.R.; Kim, Y. Cellular mechanisms of circulating tumor cells during breast cancer metastasis. Int. J. Mol. Sci. 2020, 21, 5040. [Google Scholar] [CrossRef]
- Hamilton, G.; Rath, B. Mesenchymal-epithelial transition and circulating tumor cells in small cell lung cancer. Adv. Exp. Med. Biol. 2017, 994, 229–245. [Google Scholar] [CrossRef]
- Hamidi, S.; Nakaya, Y.; Nagai, H.; Alev, C.; Kasukawa, T.; Chhabra, S.; Lee, R.; Niwa, H.; Warmflash, A.; Shibata, T.; et al. Mesenchymal-epithelial transition regulates initiation of pluripotency exit before gastrulation. Development 2020, 147, dev.184960. [Google Scholar] [CrossRef]
- Gkountela, S.; Castro-Giner, F.; Szczerba, B.M.; Vetter, M.; Landin, J.; Scherrer, R.; Krol, I.; Scheidmann, M.C.; Beisel, C.; Stirnimann, C.U.; et al. Circulating Tumor Cell Clustering Shapes DNA Methylation to Enable Metastasis Seeding. Cell 2019, 176, 98–112. [Google Scholar] [CrossRef] [Green Version]
- Meng, H.; Cao, Y.; Qin, J.; Song, X.; Zhang, Q.; Shi, Y.; Cao, L. DNA methylation, its mediators and genome integrity. Int. J. Biol. Sci. 2015, 11, 604–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda Furtado, C.L.; Dos Santos Luciano, M.C.; Da Silva Santos, R.; Furtado, G.P.; Moraes, M.O.; Pessoa, C. Epidrugs: Targeting epigenetic marks in cancer treatment. Epigenetics 2019, 14, 1164–1176. [Google Scholar] [CrossRef] [PubMed]
- Skvortsova, K.; Stirzaker, C.; Taberlay, P. The DNA methylation landscape in cancer. Essays Biochem. 2019, 63, 797–811. [Google Scholar] [CrossRef]
- Morgan, A.E.; Davies, T.J.; McAuley, M.T. The role of DNA methylation in ageing and cancer. Proc. Nutr. Soc. 2018, 77, 412–422. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Aguilera, O.; Depreux, P.; Halby, L.; Arimondo, P.B.; Goossens, L. DNA Methylation Targeting: The DNMT/HMT Crosstalk Challenge. Biomolecules 2017, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Yiu, A.J.; Yiu, C.Y. Biomarkers in Colorectal Cancer. Anticancer Res. 2016, 36, 1093–1102. [Google Scholar]
- Bakhoum, S.F.; Cantley, L.C. The multifaceted role of chromosomal instability in cancer and its microenvironment. Cell 2018, 174, 1347–1360. [Google Scholar] [CrossRef] [Green Version]
- Mármol, I.; Sánchez-de-Diego, C.; Dieste, A.P.; Cerrada, E.; Yoldi, M.J.R. Colorectal carcinoma: A general overview and future perspectives in colorectal cancer. Int. J. Mol. Sci. 2017, 18, 197. [Google Scholar] [CrossRef] [Green Version]
- Setaffy, L.; Langner, C. Microsatellite instability in colorectal cancer: Clinicopathological significance. Polish J. Pathol. 2015, 66, 203–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, H.; Imai, K. Microsatellite instability: An update. Arch. Toxicol. 2015, 89, 899–921. [Google Scholar] [CrossRef]
- Nojadeh, J.N.; Sharif, S.B.; Sakhinia, E. Microsatellite instability in colorectal cancer. EXCLI J. 2018, 17, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Toyota, M.; Ahuja, N.; Ohe-Toyota, M.; Herman, J.G.; Baylin, S.B.; Issa, J.-P. CpG island methylator phenotype in colorectal cancer. Proc. Natl. Acad. Sci. USA 1999, 96, 8681–8686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, M.; Gao, X.; Zhang, Y.; Hoffmeister, M.; Brenner, H. Different definitions of CpG island methylator phenotype and outcomes of colorectal cancer: A systematic review. Clin. Epigenetics 2016, 8, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Tang, L.; Yi, J.; Li, G.; Lu, Y.; Xu, Y.; Zhao, S.; Mao, R.; Li, X.; Ren, L.; et al. Unique characteristics of CpG island methylator phenotype (CIMP) in a Chinese population with colorectal cancer. BMC Gastroenterol. 2019, 19, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, H.; Yamamoto, E.; Maruyama, R.; Niinuma, T.; Kai, M. Biological significance of the CpG island methylator phenotype. Biochem. Biophys. Res. Commun. 2014, 455, 35–42. [Google Scholar] [CrossRef]
- Kim, Y.S.; Deng, G. Epigenetic Changes (Aberrant DNA Methylation) in Colorectal Neoplasia. Gut Liver 2007, 1, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Park, J.Y. Promoter Hypermethylation in Prostate Cancer. Cancer Control 2010, 17, 245–255. [Google Scholar] [CrossRef]
- Lee, D.D.; Leão, R.; Komosa, M.; Gallo, M.; Zhang, C.H.; Lipman, T.; Remke, M.; Heidari, A.; Nunes, N.M.; Apolónio, J.D.; et al. DNA hypermethylation within TERT promoter upregulates TERT expression in cancer. J. Clin. Investig. 2019, 129, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Imperiale, T.F.; Ransohoff, D.F.; Itzkowitz, S.H.; Levin, T.R.; Lavin, P.; Lidgard, G.P.; Ahlquist, D.A.; Berger, B.M. Multitarget Stool DNA Testing for Colorectal-Cancer Screening. N. Engl. J. Med. 2014, 370, 1287–1297. [Google Scholar] [CrossRef] [Green Version]
- Cassinotti, E.; Melson, J.; Liggett, T.; Melnikov, A.; Yi, Q.; Replogle, C.; Mobarhan, S.; Boni, L.; Segato, S.; Levenson, V. DNA methylation patterns in blood of patients with colorectal cancer and adenomatous colorectal polyps. Int. J. Cancer 2012, 131, 1153–1157. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Chen, H.Y.; Bai, E.Q.; Luo, Y.X.; Fu, R.J.; He, Y.S.; Jiang, J.; Wang, H.Q. Development of a multiplex MethyLight assay for the detection of multigene methylation in human colorectal cancer. Cancer Genet. Cytogenet. 2010, 202, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Guinney, J.; Dienstmann, R.; Wang, X.; de Reyniès, A.; Schlicker, A.; Soneson, C.; Marisa, L.; Roepman, P.; Nyamundanda, G.; Angelino, P.; et al. The consensus molecular subtypes of colorectal cancer. Nat. Med. 2015, 21, 1350–1356. [Google Scholar] [CrossRef] [PubMed]
- Casalino, L.; Verde, P. Multifaceted roles of dna methylation in neoplastic transformation, from tumor suppressors to emt and metastasis. Genes 2020, 11, 922. [Google Scholar] [CrossRef]
- Greenberg, M.V.C.; Bourc’his, D. The diverse roles of DNA methylation in mammalian development and disease. Nat. Rev. Mol. Cell Biol. 2019, 20, 590–607. [Google Scholar] [CrossRef]
- Yin, Y.; Morgunova, E.; Jolma, A.; Kaasinen, E.; Sahu, B.; Das, P.K.; Kivioja, T.; Dave, K.; Zhong, F.; Nitta, R.; et al. Impact of cytosine methylation on DNA bindgin specificities of human transcription factors. Science 2017, 356, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Mikeska, T.; Bock, C.; Do, H.; Dobrovic, A. DNA methylation biomarkers in cancer: Progress towards clinical implementation. Expert Rev. Mol. Diagn. 2012, 12, 473–487. [Google Scholar] [CrossRef] [Green Version]
- Dehan, P.; Kustermans, G.; Guenin, S.; Horion, J.; Boniver, J.; Delvenne, P. DNA methylation and cancer diagnosis. Expert Rev. Mol. Diagn. 2009, 9, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Levenson, V.V. DNA methylation as a universal biomarker. Expert Rev. Mol. Diagn. 2010, 10, 481–488. [Google Scholar] [CrossRef] [Green Version]
- Sobanski, T.; Arantes, L.M.R.B.; dos Santos, W.; Matsushita, M.; de Oliveira, M.A.; Costa, M.; de Carvalho, A.C.; Berardinelli, G.N.; Syrjänen, K.; Reis, R.M.; et al. Methylation profile of colon cancer genes in colorectal precursor lesions and tumor tissue: Perspectives for screening. Scand. J. Gastroenterol. 2021, 56, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Warren, J.D.; Xiong, W.; Bunker, A.M.; Vaughn, C.P.; Furtado, L.V.; Roberts, W.L.; Fang, J.C.; Samowitz, W.S.; Heichman, K.A. Septin 9 methylated DNA is a sensitive and specific blood test for colorectal cancer. BMC Med. 2011, 9, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, S.M.; Ross, J.P.; Drew, H.R.; Ho, T.; Brown, G.S.; Saunders, N.F.W.; Duesing, K.R.; Buckley, M.J.; Dunne, R.; Beetson, I.; et al. A panel of genes methylated with high frequency in colorectal cancer. BMC Cancer 2014, 14, 54. [Google Scholar] [CrossRef] [Green Version]
- Ju, H.X.; An, B.; Okamoto, Y.; Shinjo, K.; Kanemitsu, Y.; Komori, K.; Hirai, T.; Shimizu, Y.; Sano, T.; Sawaki, A.; et al. Distinct profiles of epigenetic evolution between colorectal cancers with and without metastasis. Am. J. Pathol. 2011, 178, 1835–1846. [Google Scholar] [CrossRef]
- De Maglio, G.; Casagrande, M.; Guardascione, M.; Fontanella, C.; Lutrino, S.E.; Rihawi, K.; Pisa, F.E.; Tuniz, F.; Fasola, G.; Pizzolitto, S.; et al. MGMT promoter methylation status in brain metastases from colorectal cancer and corresponding primary tumors. Future Oncol. 2015, 11, 1201–1209. [Google Scholar] [CrossRef] [PubMed]
- Umetani, N.; Takeuchi, H.; Fujimoto, A.; Shinozaki, M.; Bilchik, A.J.; Hoon, D.S.B. Epigenetic inactivation of ID4 in colorectal carcinomas correlates with poor differentiation and unfavorable prognosis. Clin. Cancer Res. 2004, 10, 7475–7483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.S.; Lee, J.; Sidransky, D. DNA methylation markers in colorectal cancer. Cancer Metastasis Rev. 2010, 29, 181–206. [Google Scholar] [CrossRef]
- Tshuikina, M.; Jernberg-Wiklund, H.; Nilsson, K.; Öberg, F. Epigenetic silencing of the interferon regulatory factor ICSBP/IRF8 in human multiple myeloma. Exp. Hematol. 2008, 36, 1673–1681. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, L.; Li, L.L.; Wang, Z.; Ying, J.; Fan, Y.; Xu, B.; Wang, L.; Liu, Q.; Chen, G.; et al. Interferon regulatory factor 8 functions as a tumor suppressor in renal cell carcinoma and its promoter methylation is associated with patient poor prognosis. Cancer Lett. 2014, 354, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Petko, Z.; Dzieciatkowski, S.; Lin, L.; Ghiassi, M.; Stain, S.; Chapman, W.C.; Washington, M.K.; Willis, J.; Markowitz, S.D.; et al. CpG island methylation of genes accumulates during the adenoma progression step of the multistep pathogenesis of colorectal cancer. Genes Chromosom. Cancer 2006, 45, 781–789. [Google Scholar] [CrossRef]
- Wynter, C.V.A.; Walsh, M.D.; Higuchi, T.; Leggett, B.A.; Young, J.; Jass, J.R. Methylation patterns define two types of hyperplastic polyp associated with colorectal cancer. Gut 2004, 53, 573–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, S.; Hou, Y.; Zhou, X.; Li, X.; Yong, J.; Wang, Y.; Wang, W.; Yan, J.; Hu, B.; Guo, H.; et al. Single-cell multiomics sequencing and analyses of human colorectal cancer. Science 2018, 362, 1060–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Guo, L.; Tang, W.; Ma, Y.; Wang, X.; Shao, Y.; Zhao, H.; Ying, J. Identification of DNA methylation biomarkers for risk of liver metastasis in early-stage colorectal cancer. Clin. Epigenetics 2021, 13, 126. [Google Scholar] [CrossRef] [PubMed]
- Rezasoltani, S.; Asadzadeh-Aghdaei, H.; Nazemalhosseini-Mojarad, E.; Dabiri, H.; Ghanbari, R.; Zali, M.R. Gut microbiota, epigenetic modification and colorectal cancer. Iran. J. Microbiol. 2017, 9, 55–63. [Google Scholar]
- Ghadimi, D.; Helwig, U.; Schrezenmeir, J.; Heller, K.J.; de Vrese, M. Epigenetic imprinting by commensal probiotics inhibits the IL-23/IL-17 axis in an in vitro model of the intestinal mucosal immune system. J. Leukoc. Biol. 2012, 92, 895–911. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Owen, J.L.; Lightfoot, Y.L.; Kladde, M.P.; Mohamadzadeh, M. Microbiota impact on the epigenetic regulation of colorectal cancer. Trends Mol. Med. 2013, 19, 714–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Zhu, M.-J. Butyrate Inhibits Indices of Colorectal Carcinogenesis via Enhancing α-Ketoglutarate-Dependent DNA Demethylation of Mismatch Repair Genes. Mol. Nutr. Food Res. 2018, 62, 1700932. [Google Scholar] [CrossRef]
- Wang, Q.; Ye, J.; Fang, D.; Lv, L.; Wu, W.; Shi, D.; Li, Y.; Yang, L.; Bian, X.; Wu, J.; et al. Multi-omic profiling reveals associations between the gut mucosal microbiome, the metabolome, and host DNA methylation associated gene expression in patients with colorectal cancer. BMC Microbiol. 2020, 20, 83. [Google Scholar] [CrossRef]
- Tahara, T.; Yamamoto, E.; Suzuki, H.; Maruyama, R.; Chung, W.; Garriga, J.; Jelinek, J.; Yamano, H.-O.; Sugai, T.; An, B.; et al. Fusobacterium in Colonic Flora and Molecular Features of Colorectal Carcinoma. Cancer Res. 2014, 74, 1311–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.E.; Kim, J.H.; Cho, N.Y.; Lee, H.S.; Kang, G.H. Intratumoral Fusobacterium nucleatum abundance correlates with macrophage infiltration and CDKN2A methylation in microsatellite-unstable colorectal carcinoma. Virchows Arch. 2017, 471, 329–336. [Google Scholar] [CrossRef]
- Denzer, L.; Schroten, H.; Schwerk, C. From gene to protein—How bacterial virulence factors manipulate host gene expression during infection. Int. J. Mol. Sci. 2020, 21, 3730. [Google Scholar] [CrossRef]
- Moon, D.C.; Choi, C.H.; Lee, S.M.; Lee, J.H.; Kim, S., II; Kim, D.S.; Lee, J.C. Nuclear translocation of Acinetobacter baumannii transposase induces DNA methylation of CpG regions in the promoters of E-cadherin gene. PLoS ONE 2012, 7, e38974. [Google Scholar] [CrossRef] [Green Version]
- Makanji, Y.; Zhu, J.; Mishra, R.; Holmquist, C.; Wong, W.P.S.; Schwartz, N.B.; Mayo, K.E.; Woodruff, T.K. Inhibin at 90: From Discovery to Clinical Application, a Historical Review. Endocr. Rev. 2014, 35, 747–794. [Google Scholar] [CrossRef]
- Robertson, D.M.; Burger, H.G.; Fuller, P.J. Inhibin/activin and ovarian cancer. Endocr. Relat. Cancer 2004, 11, 35–49. [Google Scholar] [CrossRef] [Green Version]
- Goney, M.P.; Wilce, M.C.; Wilce, J.A.; Stocker, W.A.; Goodchild, G.M.; Chan, K.L.; Harrison, C.A.; Walton, K.L. Engineering the ovarian hormones inhibin A and inhibin B to enhance synthesis and activity. Endocrinology 2020, 161, bqaa099. [Google Scholar] [CrossRef]
- Mayor, R.; Casadomé, L.; Azuara, D.; Moreno, V.; Clark, S.J.; Capellà, G.; Peinado, M.A. Long-range epigenetic silencing at 2q14.2 affects most human colorectal cancers and may have application as a non-invasive biomarker of disease. Br. J. Cancer 2009, 100, 1534–1539. [Google Scholar] [CrossRef] [Green Version]
- Karpinski, P.; Ramsey, D.; Grzebieniak, Z.; Sasiadek, M.M.; Blin, N. The CpG island methylator phenotype correlates with long-range epigenetic silencing in colorectal cancer. Mol. Cancer Res. 2008, 6, 585–591. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Li, Y.; Cui, Y.; Guo, Y. Identifications of Candidate Genes Significantly Associated With Rectal Cancer by Integrated Bioinformatics Analysis. Technol. Cancer Res. Treat. 2020, 19, 1533033820973270. [Google Scholar] [CrossRef]
- Gerarduzzi, C.; Kumar, R.K.; Trivedi, P.; Ajay, A.K.; Iyer, A.; Boswell, S.; Hutchinson, J.N.; Waikar, S.S.; Vaidya, V.S. Silencing SMOC2 ameliorates kidney fibrosis by inhibiting fibroblast to myofibroblast transformation. JCI Insight 2017, 2, e90299. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.C.; Dong, Q.Z.; Zhang, X.F.; Deng, B.; Jia, H.L.; Ye, Q.H.; Qin, L.X.; Wu, X.Z. MicroRNA-29a suppresses cell proliferation by targeting SPARC in hepatocellular carcinoma. Int. J. Mol. Med. 2012, 30, 1321–1326. [Google Scholar] [CrossRef] [PubMed]
- Robert, G.; Gaggioli, C.; Bailet, O.; Chavey, C.; Abbe, P.; Aberdam, E.; Sabatié, E.; Cano, A.; Garcia De Herreros, A.; Ballotti, R.; et al. SPARC represses E-cadherin and induces mesenchymal transition during melanoma development. Cancer Res. 2006, 66, 7516–7523. [Google Scholar] [CrossRef] [Green Version]
- Deng, B.; Qu, L.; Li, J.; Fang, J.; Yang, S.; Cao, Z.; Mei, Z.; Sun, X. MiRNA-211 suppresses cell proliferation, migration and invasion by targeting SPARC in human hepatocellular carcinoma. Sci. Rep. 2016, 6, 26679. [Google Scholar] [CrossRef] [Green Version]
- Su, J.R.; Kuai, J.H.; Li, Y.Q. Smoc2 potentiates proliferation of hepatocellular carcinoma cells via promotion of cell cycle progression. World J. Gastroenterol. 2016, 22, 10053–10063. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; dan Ju, D.; Yang, G.D.; Zhu, L.Y.; Yang, X.M.; Li, J.; Song, W.W.; Wang, J.H.; Zhang, C.C.; Zhang, Z.G.; et al. Targeting cancer stem cell signature gene SMOC-2 Overcomes chemoresistance and inhibits cell proliferation of endometrial carcinoma. EBioMedicine 2019, 40, 276–289. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Choi, J.H.; Lee, J.Y.; Kang, J.H.; Myung, J.K.; Kim, W.H.; Jang, B.G. Downregulation of SMOC2 expression in papillary thyroid carcinoma and its prognostic significance. Sci. Rep. 2020, 10, 4853. [Google Scholar] [CrossRef]
- Dhont, L.; Mascaux, C.; Belayew, A. The helicase-like transcription factor (HLTF) in cancer: Loss of function or oncomorphic conversion of a tumor suppressor? Cell. Mol. Life Sci. 2016, 73, 129–145. [Google Scholar] [CrossRef]
- Hishiki, A.; Hara, K.; Ikegaya, Y.; Yokoyama, H.; Shimizu, T.; Sato, M.; Hashimoto, H. Structure of a novel DNA-binding domain of Helicase-like Transcription Factor (HLTF) and its functional implication in DNA damage tolerance. J. Biol. Chem. 2015, 290, 13215–13223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallo, D.; Brown, G.W. Post-replication repair: Rad5/HLTF regulation, activity on undamaged templates, and relationship to cancer. Crit. Rev. Biochem. Mol. Biol. 2019, 54, 301–332. [Google Scholar] [CrossRef]
- Helmer, R.A.; Martinez-Zaguilan, R.; Kaur, G.; Smith, L.A.; Dufour, J.M.; Chilton, B.S. Helicase-like transcription factor-deletion from the tumor microenvironment in a cell linederived xenograft model of colorectal cancer reprogrammed the human transcriptome-Snitroso-proteome to promote inflammation and redirect metastasis. PLoS ONE 2021, 16, e0251132. [Google Scholar] [CrossRef]
- Unk, I.; Hajdú, I.; Blastyák, A.; Haracska, L. Role of yeast Rad5 and its human orthologs, HLTF and SHPRH in DNA damage tolerance. DNA Repair 2010, 9, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Elserafy, M.; Abugable, A.A.; Atteya, R.; El-Khamisy, S.F. Rad5, HLTF, and SHPRH: A Fresh View of an Old Story. Trends Genet. 2018, 34, 574–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Zhang, L.; Vartuli, R.L.; Ford, H.L.; Zhao, R. The Eya Phosphatase: Its unique role in cancer. Int. J. Biochem. Cell Biol. 2018, 96, 165–170. [Google Scholar] [CrossRef]
- Tadjuidje, E.; Hegde, R.S. The Eyes Absent proteins in development and disease. Cell. Mol. Life Sci. 2013, 70, 1897–1913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, X.; Peng, J.X.; Hao, X.Y.; Cai, J.P.; Liang, L.J.; Zhai, J.M.; Zhang, K.S.; Lai, J.M.; Yin, X.Y. DNA Methylation Profiling Identifies EYA4 Gene as a Prognostic Molecular Marker in Hepatocellular Carcinoma. Ann. Surg. Oncol. 2014, 21, 3891–3899. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Tae, C.H.; Hong, S.N.; Min, B.H.; Chang, D.K.; Rhee, P.L.; Kim, J.J.; Kim, H.C.; Kim, D.H.; Kim, Y.H. EYA4 Acts as a New Tumor Suppressor Gene in Colorectal Cancer. Mol. Carcinog. 2015, 54, 1748–1757. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Li, Y.; Shi, X.; Yang, W.; Zhou, F.; Sun, N.; He, J. Aberrant methylation of EYA4 promotes epithelial-mesenchymal transition in esophageal squamous cell carcinoma. Cancer Sci. 2018, 109, 1811–1824. [Google Scholar] [CrossRef]
- Peters, I.; Eggers, H.; Atschekzei, F.; Hennenlotter, J.; Waalkes, S.; Tränkenschuh, W.; Groãhennig, A.; Merseburger, A.S.; Stenzl, A.; Kuczyk, M.A.; et al. GATA5 CpG island methylation in renal cell cancer: A potential biomarker for metastasis and disease progression. BJU Int. 2012, 110, E144–E152. [Google Scholar] [CrossRef]
- Guo, M.; Akiyama, Y.; House, M.G.; Hooker, C.M.; Heath, E.; Gabrielson, E.; Yang, S.C.; Han, Y.; Baylin, S.B.; Herman, J.G.; et al. Hypermethylation of the GATA genes in lung cancer. Clin. Cancer Res. 2004, 10, 7917–7924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, X.Z.; Akiyama, Y.; Pan, K.F.; Liu, Z.J.; Lu, Z.M.; Zhou, J.; Gu, L.K.; Dong, C.X.; Zhu, B.D.; Ji, J.F.; et al. Methylation of GATA-4 and GATA-5 and development of sporadic gastric carcinomas. World J. Gastroenterol. 2010, 16, 1201–1208. [Google Scholar] [CrossRef]
- Akiyama, Y.; Watkins, N.; Suzuki, H.; Jair, K.-W.; van Engeland, M.; Esteller, M.; Sakai, H.; Ren, C.-Y.; Yuasa, Y.; Herman, J.G.; et al. GATA-4 and GATA-5 Transcription Factor Genes and Potential Downstream Antitumor Target Genes Are Epigenetically Silenced in Colorectal and Gastric Cancer. Mol. Cell. Biol. 2003, 23, 8429–8439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hempstead, B.L. Brain-Derived Neurotrophic Factor: Three Ligands, Many Actions. Trans. Am. Clin. Climatol. Assoc. 2015, 126, 9–19. [Google Scholar]
- Lu, B.; Pang, P.T.; Woo, N.H. The yin and yang of neurotrophin action. Nat. Rev. Neurosci. 2005, 6, 603–614. [Google Scholar] [CrossRef] [Green Version]
- Tajbakhsh, A.; Mokhtari-Zaer, A.; Rezaee, M.; Afzaljavan, F.; Rivandi, M.; Hassanian, S.M.; Ferns, G.A.; Pasdar, A.; Avan, A. Therapeutic Potentials of BDNF/TrkB in Breast Cancer; Current Status and Perspectives. J. Cell. Biochem. 2017, 118, 2502–2515. [Google Scholar] [CrossRef]
- Mazouffre, C.; Geyl, S.; Perraud, A.; Blondy, S.; Jauberteau, M.O.; Mathonnet, M.; Verdier, M. Dual inhibition of BDNF/TrkB and autophagy: A promising therapeutic approach for colorectal cancer. J. Cell. Mol. Med. 2017, 21, 2610–2622. [Google Scholar] [CrossRef]
- Kundakovic, M.; Gudsnuk, K.; Herbstman, J.B.; Tang, D.; Perera, F.P.; Champagne, F.A. DNA methylation of BDNF as a biomarker of early-life adversity. Proc. Natl. Acad. Sci. USA 2015, 112, 6807–6813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boehm, E.; Zaganelli, S.; Maundrell, K.; Jourdain, A.A.; Thore, S.; Martinou, J.C. FASTKD1 and FASTKD4 have opposite effects on expression of specific mitochondrial RNAs, depending upon their endonuclease-like RAP domain. Nucleic Acids Res. 2017, 45, 6135–6146. [Google Scholar] [CrossRef]
- Jourdain, A.A.; Popow, J.; De La Fuente, M.A.; Martinou, J.C.; Anderson, P.; Simarro, M. Survey and summary: The FASTK family of proteins: Emerging regulators of mitochondrial RNA biology. Nucleic Acids Res. 2017, 45, 10941–10947. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Zhao, C.; Liu, X.; Su, W.; Duan, G.; Xie, Z.; Chu, S.; Gao, Y. Knockdown of TBRG4 affects tumorigenesis in human H1299 lung cancer cells by regulating DDIT3, CAV1 and RRM2. Oncol. Lett. 2018, 15, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Luo, Q.; Liu, M.; Zhang, C.; Jia, Y.; Tong, R.; Yang, L.; Fu, X. TBRG4 silencing promotes progression of squamous cell carcinoma via regulation of CAV-1 expression and ROS formation. Cell. Mol. Biol. 2020, 66, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Zhou, P.; Wang, Z.; Zhang, X.L.; Liao, F.X.; Hu, Y.; Chang, J. Knockdown of TBRG4 suppresses proliferation, invasion and promotes apoptosis of osteosarcoma cells by downregulating TGF-β1 expression and PI3K/AKT signaling pathway. Arch. Biochem. Biophys. 2020, 686, 108351. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Gene | Pathways | Type of Marker | Samples | References |
|---|---|---|---|---|
| ALX4 | Tumor suppressor of Wnt/β-catenin pathway [40] | Diagnosis | Plasma | [41,42,43,44,45,46,47] |
| BCAT1 | mTOR1 regulator | Diagnosis/Prognosis | Plasma, FFPE | [41,44,48,49,50] |
| BMP3 | Tumor suppressor via TAK1/JNK and ActRIIB/SMAD2-dependent gene [51] | Diagnosis | Plasma, Stool, FFPE | [41,43,46,48,49,52] |
| EYA4 | Tumor suppressor of Wnt/β-catenin pathway [53] | Diagnosis/Prognosis | Plasma | [41,43,44,54] |
| GATA4/GATA5 | Tumor suppressor of Wnt/β-catenin pathway [55] | Diagnosis | Plasma, Stool | [41,43,44,49,56] |
| HIC | Tumor suppressor of IL-6/STAT3 pathway [57] | Prognosis | Plasma | [41,43,44,56] |
| HLTF | Tumor suppressor TGF- β/SMAD pathway [58] | Diagnosis/Prognosis/Predictive | Plasma, Stool | [43,44,48,59] |
| IGFBP-3 | Involved in TGF-β independent function [60] | Prognosis/Predictive | Plasma, FFPE | [42,45,54,61,62] |
| IKZF1/IKZF2 | Related to binding to relevant genes as PTPN6 and MEIS2 [50,63] | Diagnosis/Prognosis | Plasma, FFPE | [41,44,48,49,50] |
| IRF4 | Promotes proliferation through JNK/Jun pathway [64] | Diagnosis | Plasma | [41,44,49] |
| ITGA4 | Increases migration through interacting with VCAM-1 [65] | Diagnosis | Stool, FFPE | [43,44,48,56] |
| LINE-1 | Inserts into genes causing gene disruption [66] | Diagnosis/Prognosis | Plasma | [41,42,44,67] |
| MGMT | DNA repair system [68] | Diagnosis | Plasma, Stool, FFPE | [41,42,43,44,45,49,69] |
| MLH1 | Member of mismatch repair (MMR) machinery [70] | Diagnosis | Stool, FFPE | [42,43,44,48,49,54,69,71,72,73] |
| NDRG4 | Inhibition of PI3K/AKT pathway [74] | Diagnosis/Prognosis | Stool, FFPE | [41,43,48,49,52,56] |
| OSMR | Activation of STAT3/FAK/Src pathway [75] | Diagnosis | Plasma, Stool | [41,43,48,56] |
| PRIMA1 | Activates mutant p53 pathway [76] | Diagnosis | Plasma | [41,49,52] |
| RASSF1A/RASSF2A | Interacts with Hippo and Wnt Pathway [77] | Diagnosis/Prognosis/Predictive | Plasma, Stool, FFPE | [41,43,44,49,52,78] |
| SDC2 | Promotes MAPK and EMT pathway activation [79] | Diagnosis | Plasma, Stool | [41,44,45,46,49,52,56,80] |
| SEPT9 | Activates Rho/ROCK and FAK pathway [81] | Diagnosis/Prognosis/Predictive | Plasma, FFPE | [41,42,45,46,48,49,52,54,82] |
| SFRP1/SFRP2 | Inactivation of Wnt pathway [83] | Diagnosis | Plasma, Stool, FFPE | [41,43,44,45,48,49,52,56] |
| TAC1 | - | Diagnosis/Prognosis/Predictive | Plasma | [41,44,54,82] |
| TFPI2 | Involved in TWIST-integrin α5 [84] | Diagnosis | Plasma, Stool | [43,48,49] |
| VIM | Interacts with MAPK/ERK Pathway [85] | Diagnosis/Prognosis | Plasma, Stool, FFPE, Urine | [41,45,46,48,49,52,56] |
| INHBB | Activates ERK/Smad2/3 pathway [86] | Diagnosis/Prognosis | FFPE | [86] |
| BDNF | Activates AKT pathway [87] | Prognosis | FFPE | [88] |
| SMOC2 | Activates Wnt/β-catenin pathway [89] | Prognosis | FFPE | [89] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutierrez, A.; Demond, H.; Brebi, P.; Ili, C.G. Novel Methylation Biomarkers for Colorectal Cancer Prognosis. Biomolecules 2021, 11, 1722. https://doi.org/10.3390/biom11111722
Gutierrez A, Demond H, Brebi P, Ili CG. Novel Methylation Biomarkers for Colorectal Cancer Prognosis. Biomolecules. 2021; 11(11):1722. https://doi.org/10.3390/biom11111722
Chicago/Turabian StyleGutierrez, Alvaro, Hannah Demond, Priscilla Brebi, and Carmen Gloria Ili. 2021. "Novel Methylation Biomarkers for Colorectal Cancer Prognosis" Biomolecules 11, no. 11: 1722. https://doi.org/10.3390/biom11111722