
Functions and Inhibition of Galectin-7, an Emerging Target in Cellular Pathophysiology


Abstract
:1. Introduction: Galectin-7
2. Galectin-7, a Convergence of Pathology with Physiology
2.1. Role in Epidermal Homeostasis of Skin, Corneal and Periodontal Tissue
2.2. Roles in Cancer
2.3. Role in Pre-Eclampsia, Menstruation and Recurrent Pregnancy Loss
2.4. Roles in Allergic Inflammatory and Autoimmune Diseases
2.5. Role in Transplant Rejection
3. Drug Potential of Galectin-7 Inhibitors and Galectin-7 as a Biomarker
3.1. Drug Potential of Galectin-7 Inhibitors
3.2. Galectin-7 as a Biomarker
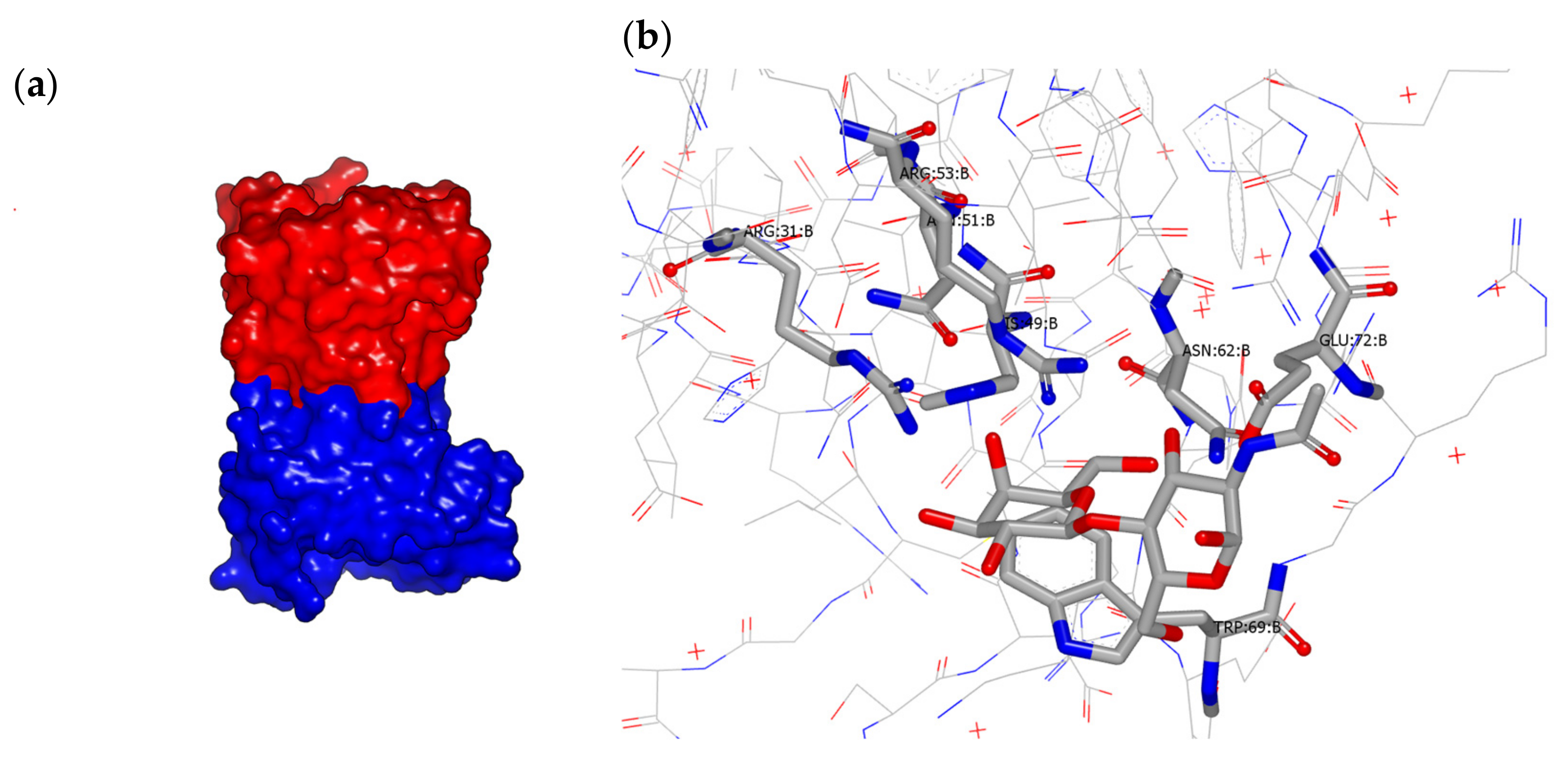
4. Structural Features
5. Small-Molecule Carbohydrate and Non-Carbohydrate Galectin-7 Inhibitors
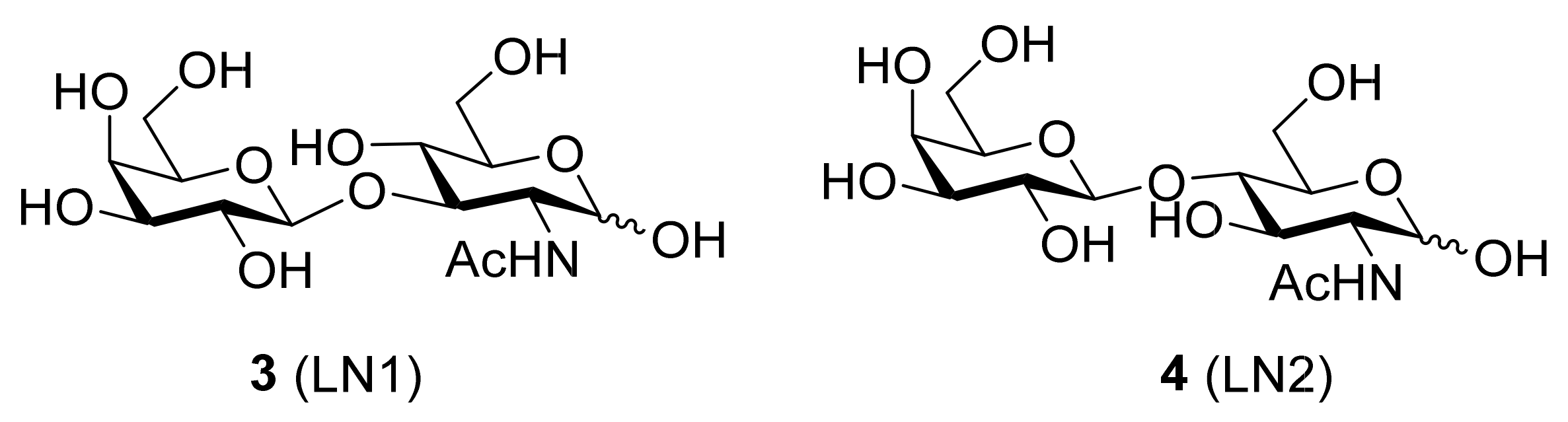
5.1. Inhibitors Based on a Carbohydrate Scaffold
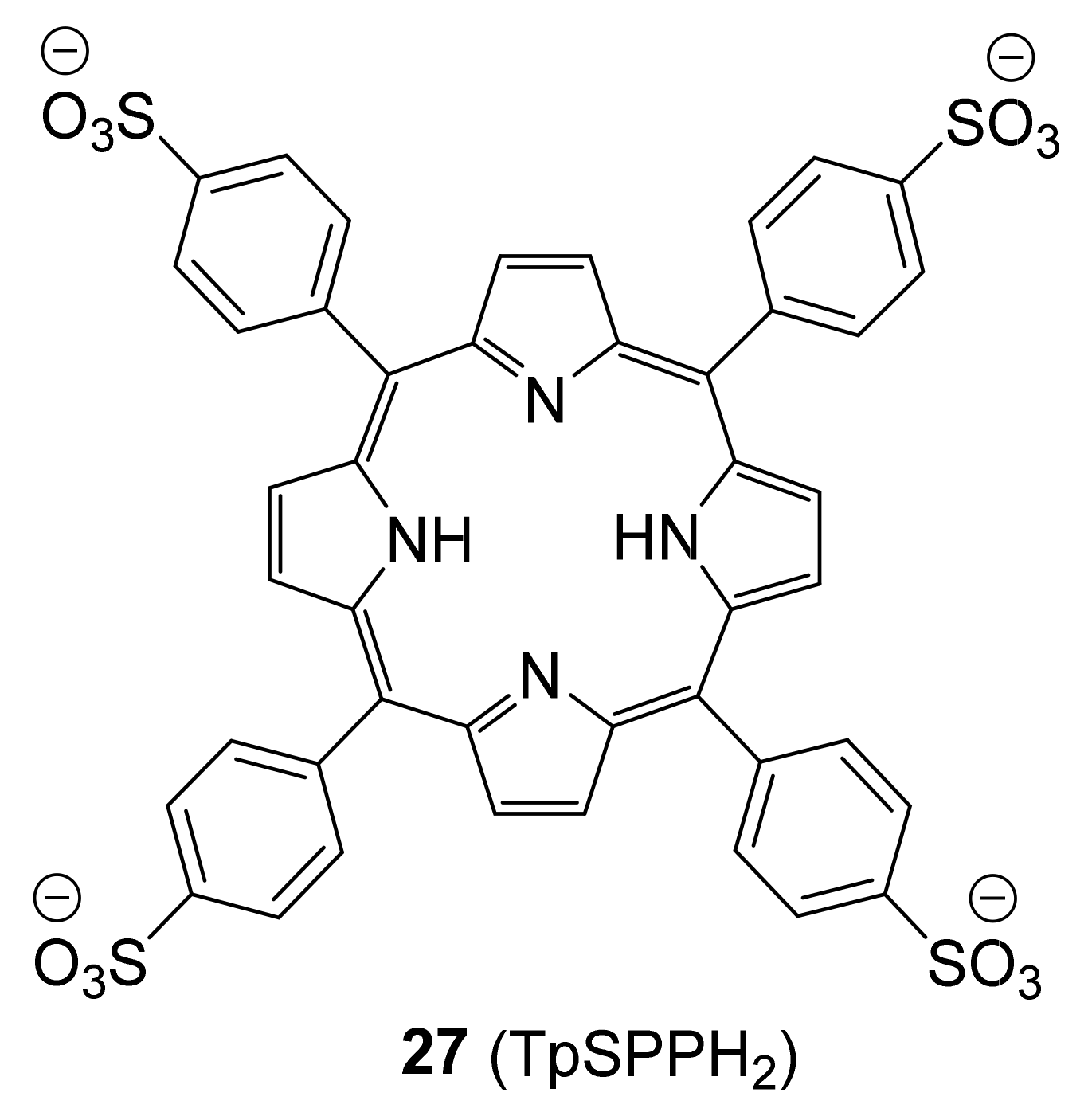
5.2. Inhibitors Based on a Non-Carbohydrate Scaffold
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Varki, A.; Cummings, R.D.; Esko, J.D.; Stanley, P.; Hart, G.W.; Aebi, M.; Darvill, A.G.; Kinoshita, T.; Packer, N.H.; Prestegard, J.H.; et al. Essentials of Glycobiology, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2017. [Google Scholar]
- Bertuzzi, S.; Quintana, J.I.; Ardá, A.; Gimeno, A.; Jiménez-Barbero, J. Targeting Galectins With Glycomimetics. Front. Chem. 2020, 8, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Si, Y.; Yao, Y.; Jaramillo Ayala, G.; Li, X.; Han, Q.; Zhang, W.; Xu, X.; Tai, G.; Mayo, K.H.; Zhou, Y.; et al. Human galectin-16 has a pseudo ligand binding site and plays a role in regulating c-Rel-mediated lymphocyte activity. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129755. [Google Scholar] [CrossRef] [PubMed]
- Madsen, P.; Rasmussen, H.H.; Flint, T.; Gromov, P.; Kruse, T.A.; Honore, B.; Vorum, H.; Celis, J.E. Cloning, expression, and chromosome mapping of human galectin-7. J. Biol. Chem. 1995, 270, 5823–5829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magnaldo, T.; Bernerd, F.; Darmon, M. Galectin-7, a Human 14 kDa S-lectin, Specifically Expressed in keratinocytes and Sensitive to Retinoic acid. Dev. Biol. 1995, 168, 259–271. [Google Scholar] [CrossRef]
- St-pierre, Y. Towards a Better Understanding of the Relationships between Galectin-7, p53 and MMP-9 during Cancer Progression. Biomolecules 2021, 11, 879. [Google Scholar] [CrossRef]
- Johannes, L.; Jacob, R.; Leffler, H. Galectins at a glance. J. Cell Sci. 2018, 131, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Leonidas, D.D.; Vatzaki, E.H.; Vorum, H.; Celis, J.E.; Madsen, P.; Acharya, K.R. Structural basis for the recognition of carbohydrates by human galectin- 7. Biochemistry 1998, 37, 13930–13940. [Google Scholar] [CrossRef]
- Cooper, D.N.W.; Barondes, S.H. God must love galectins; he made so many of them. Glycobiology 1999, 9, 979–984. [Google Scholar] [CrossRef]
- Morris, S.; Ahmad, N.; Andre, S.; Kaltner, H.; Gabius, H.J.; Brenowitz, M.; Brewer, F. Quaternary solution structures of galectins-1, -3, and -7. Glycobiology 2004, 14, 293–300. [Google Scholar] [CrossRef]
- Nesmelova, I.V.; Berbís, M.Á.; Miller, M.C.; Cañada, F.J.; André, S.; Jiménez-Barbero, J.; Gabius, H.J.; Mayo, K.H. 1H, 13C, and 15N backbone and side-chain chemical shift assignments for the 31 kDa human galectin-7 (p53-induced gene 1) homodimer, a pro-apoptotic lectin. Biomol. NMR Assign. 2012, 6, 127–129. [Google Scholar] [CrossRef]
- Saussez, S.; Kiss, R. Galectin-7. Cell. Mol. Life Sci. 2006, 63, 686–697. [Google Scholar] [CrossRef]
- Advedissian, T.; Deshayes, F.; Viguier, M. Galectin-7 in epithelial homeostasis and carcinomas. Int. J. Mol. Sci. 2017, 18, 2760. [Google Scholar] [CrossRef] [Green Version]
- Bernerd, F.; Sarasin, A.; Magnaldo, T. Galectin-7 overexpression is associated with the apoptotic process in UVB-induced sunburn keratinocytes. Proc. Natl. Acad. Sci. USA 1999, 96, 11329–11334. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, T.; Hiromasa, K.; Kabashima-Kubo, R.; Yoshioka, M.; Nakamura, M. Galectin-7, induced by cis-urocanic acid and ultraviolet B irradiation, down-modulates cytokine production by T lymphocytes. Exp. Dermatol. 2013, 22, 840–842. [Google Scholar] [CrossRef]
- Gendronneau, G.; Sidhu, S.S.; Delacour, D.; Dang, T.; Calonne, C.; Houzelstein, D.; Magnaldo, T.; Poirier, F. Galectin-7 in the Control of Epidermal Homeostasis after Injury. Mol. Biol. Cell 2008, 19, 5541–5549. [Google Scholar] [CrossRef] [Green Version]
- Gendronneau, G.; Sanii, S.; Dang, T.; Deshayes, F.; Delacour, D.; Pichard, E.; Advedissian, T.; Sidhu, S.S.; Viguier, M.; Magnaldo, T.; et al. Overexpression of galectin-7 in mouse epidermis leads to loss of cell junctions and defective skin repair. PLoS ONE 2015, 10, e0119031. [Google Scholar] [CrossRef] [Green Version]
- Advedissian, T.; Proux-Gillardeaux, V.; Nkosi, R.; Peyret, G.; Nguyen, T.; Poirier, F.; Viguier, M.; Deshayes, F. E-cadherin dynamics is regulated by galectin-7 at epithelial cell surface. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.L.; Chiang, P.C.; Lo, C.H.; Lo, Y.H.; Hsu, D.K.; Chen, H.Y.; Liu, F.T. Galectin-7 regulates keratinocyte proliferation and differentiation through JNK-miR-203-p63 signaling. J. Investig. Dermatol. 2016, 136, 182–191. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.L.; Lo, C.H.; Huang, C.C.; Lu, M.P.; Hu, P.Y.; Chen, C.S.; Chueh, D.Y.; Chen, P.; Lin, T.N.; Lo, Y.H.; et al. Galectin-7 downregulation in lesional keratinocytes contributes to enhanced IL-17A signaling and skin pathology in psoriasis. J. Clin. Investig. 2021, 131, e130740. [Google Scholar] [CrossRef]
- Asano, Y.; Sato, S. Vasculopathy in scleroderma. Semin. Immunopathol. 2015, 37, 489–500. [Google Scholar] [CrossRef]
- Saigusa, R.; Yamashita, T.; Miura, S.; Hirabayashi, M.; Nakamura, K.; Miyagawa, T.; Fukui, Y.; Yoshizaki, A.; Sato, S.; Asano, Y. A potential contribution of decreased galectin-7 expression in stratified epithelia to the development of cutaneous and oesophageal manifestations in systemic sclerosis. Exp. Dermatol. 2019, 28, 536–542. [Google Scholar] [CrossRef]
- Blair, M. Diabetes Mellitus Review. Urol. Nurs. 2016, 36, 27–36. [Google Scholar] [CrossRef]
- Huang, S.M.; Wu, C.S.; Chiu, M.H.; Yang, H.J.; Chen, G.S.; Lan, C.C.E. High-glucose environment induced intracellular O-GlcNAc glycosylation and reduced galectin-7 expression in keratinocytes: Implications on impaired diabetic wound healing. J. Dermatol. Sci. 2017, 87, 168–175. [Google Scholar] [CrossRef] [Green Version]
- Tazhitdinova, R.; Timoshenko, A.V. The Emerging Role of Galectins and O-GlcNAc Homeostasis in Processes of Cellular Differentiation. Cells 2020, 9, 1792. [Google Scholar] [CrossRef]
- Rousselle, P.; Braye, F.; Dayan, G. Re-epithelialization of adult skin wounds: Cellular mechanisms and therapeutic strategies. Adv. Drug Deliv. Rev. 2019, 146, 344–365. [Google Scholar] [CrossRef]
- Cao, Z.; Said, N.; Wu, H.K.; Kuwabara, I.; Liu, F.T.; Panjwani, N. Galectin-7 as a potential mediator of corneal epithelial cell migration. Arch. Ophthalmol. 2003, 121, 82–86. [Google Scholar] [CrossRef]
- Lin, T.; Yu, C.C.; Liu, C.M.; Hsieh, P.L.; Liao, Y.W.; Yu, C.H.; Chen, C.J. Er:YAG laser promotes proliferation and wound healing capacity of human periodontal ligament fibroblasts through Galectin-7 induction. J. Formos. Med. Assoc. 2021, 120, 388–394. [Google Scholar] [CrossRef]
- Picorino, L. Molecular Biology of Cancer - Mechanisms, Targets and Therapeutics, 4th ed.; Oxford University Press: Oxford, UK, 2016. [Google Scholar]
- Kaur, M.; Kaur, T.; Kamboj, S.S.; Singh, J. Roles of galectin-7 in cancer. Asian Pacific J. Cancer Prev. 2016, 17, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Cagnoni, A.J.; Pérez Sáez, J.M.; Rabinovich, G.A.; Mariño, K.V. Turning-off signaling by siglecs, selectins, and galectins: Chemical inhibition of glycan-dependent interactions in cancer. Front. Oncol. 2016, 6, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Méndez-Huergo, S.P.; Blidner, A.G.; Rabinovich, G.A. Galectins: Emerging regulatory checkpoints linking tumor immunity and angiogenesis. Curr. Opin. Immunol. 2017, 45, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Kaltner, H.; Toegel, S.; Caballero, G.G.; Manning, J.C.; Ledeen, R.W.; Gabius, H.J. Galectins: Their network and roles in immunity/tumor growth control. Histochem. Cell Biol. 2017, 147, 239–256. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.A.; Tsai, M.J.; Kuo, P.L.; Hung, J.Y. Role of galectins in lung cancer (Review). Oncol. Lett. 2017, 14, 5077–5084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wdowiak, K.; Francuz, T.; Gallego-Colon, E.; Ruiz-Agamez, N.; Kubeczko, M.; Grochoła, I.; Wojnar, J. Galectin Targeted Therapy in Oncology: Current Knowledge and Perspectives. Int. J. Mol. Sci. 2018, 19, 210. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, E.; Schetters, S.T.T.; Van Kooyk, Y. The tumour glyco-code as a novel immune checkpoint for immunotherapy. Nat. Rev. Immunol. 2018, 18, 204–211. [Google Scholar] [CrossRef]
- Rodrigues, J.G.; Balmaña, M.; Macedo, J.A.; Poças, J.; Fernandes, Â.; de Freitas Junior, J.C.M.; Pinho, S.S.; Gomes, J.; Magalhães, A.; Gomes, C.; et al. Glycosylation in cancer: Selected roles in tumour progression, immune modulation and metastasis. Cell. Immunol. 2018, 333, 46–57. [Google Scholar] [CrossRef] [Green Version]
- Chetry, M.; Thapa, S.; Hu, X.; Song, Y.; Zhang, J.; Zhu, H.; Zhu, X. The role of galectins in tumor progression, treatment and prognosis of gynecological cancers. J. Cancer 2018, 9, 4742–4755. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, Y.; Wang, Y.; Wu, X. The role of galectins in cervical cancer biology and progression. Biomed Res. Int. 2018, 2018, 2175927. [Google Scholar] [CrossRef]
- Dubé-Delarosbil, C.; St-Pierre, Y. The emerging role of galectins in high-fatality cancers. Cell. Mol. Life Sci. 2018, 75, 1215–1226. [Google Scholar] [CrossRef]
- Bartolazzi, A. Galectins in cancer and translational medicine: From bench to bedside. Int. J. Mol. Sci. 2018, 19, 2934. [Google Scholar] [CrossRef] [Green Version]
- Mereiter, S.; Balmaña, M.; Campos, D.; Gomes, J.; Reis, C.A. Glycosylation in the Era of Cancer-Targeted Therapy: Where Are We Heading? Cancer Cell 2019, 36, 6–16. [Google Scholar] [CrossRef]
- Shimada, C.; Xu, R.; Al-Alem, L.; Stasenko, M.; Spriggs, D.R.; Rueda, B.R. Galectins and ovarian cancer. Cancers 2020, 12, 1421. [Google Scholar] [CrossRef]
- Pergialiotis, V.; Papoutsi, E.; Androutsou, A.; Tzortzis, A.S.; Frountzas, M.; Papapanagiotou, A.; Kontzoglou, K. Galectins-1, -3, -7, -8 and -9 as prognostic markers for survival in epithelial ovarian cancer: A systematic review and meta-analysis. Int. J. Gynecol. Obstet. 2020, 299–307. [Google Scholar] [CrossRef]
- Manero-Rupérez, N.; Martínez-Bosch, N.; Barranco, L.E.; Visa, L.; Navarro, P. The Galectin Family as Molecular Targets: Hopes for Defeating Pancreatic Cancer. Cells 2020, 9, 689. [Google Scholar] [CrossRef] [Green Version]
- Hisrich, B.V.; Young, R.B.; Sansone, A.M.; Bowens, Z.; Green, L.J.; Lessey, B.A.; Blenda, A.V. Role of human galectins in inflammation and cancers associated with endometriosis. Biomolecules 2020, 10, 230. [Google Scholar] [CrossRef] [Green Version]
- Navarro, P.; Martínez-Bosch, N.; Blidner, A.G.; Rabinovich, G.A. Impact of Galectins in Resistance to Anticancer Therapies. Clin. Cancer Res. 2020, 26, 6086–6101. [Google Scholar] [CrossRef]
- Girotti, M.R.; Salatino, M.; Dalotto-Moreno, T.; Rabinovich, G.A. Sweetening the hallmarks of cancer: Galectins as multifunctional mediators of tumor progression. J. Exp. Med. 2020, 217, 1–14. [Google Scholar] [CrossRef]
- Perrotta, R.M.; Bach, C.A.; Salatino, M.; Rabinovich, G.A. Reprogramming the tumor metastasis cascade by targeting galectin-driven networks. Biochem. J. 2021, 478, 597–617. [Google Scholar] [CrossRef]
- Rorive, S.; Eddafali, B.; Fernandez, S.; Decaestecker, C.; André, S.; Kaltner, H.; Kuwabara, I.; Liu, F.T.; Gabius, H.J.; Kiss, R.; et al. Changes in galectin-7 and cytokeratin-19 expression during the progression of malignancy in thyroid tumors: Diagnostic and biological implications. Mod. Pathol. 2002, 15, 1294–1301. [Google Scholar] [CrossRef]
- Moisan, S.; Demers, M.; Mercier, J.; Magnaldo, T.; Potworowski, E.F.; St-Pierre, Y. Upregulation of galectin-7 in murine lymphoma cells is associated with progression toward an aggressive phenotype. Leukemia 2003, 17, 751–759. [Google Scholar] [CrossRef] [Green Version]
- Demers, M.; Couillard, J.; Giglia-Mari, G.; Magnaldo, T.; St-Pierre, Y. Increased galectin-7 gene expression in lymphoma cells is under the control of DNA methylation. Biochem. Biophys. Res. Commun. 2009, 387, 425–429. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Hwang, J.A.; Ro, J.Y.; Lee, Y.S.; Chun, K.H. Galectin-7 is epigenetically-regulated tumor suppressor in gastric cancer. Oncotarget 2013, 4, 1461–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demers, M.; Magnaldo, T.; St-Pierre, Y. A novel function for galectin-7: Promoting tumorigenesis by up-regulating MMP-9 gene expression. Cancer Res. 2005, 65, 5205–5210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demers, M.; Biron-Pain, K.; Hébert, J.; Lamarre, A.; Magnaldo, T.; St-Pierre, Y. Galectin-7 in lymphoma: Elevated expression in human lymphoid malignancies and decreased lymphoma dissemination by antisense strategies in experimental model. Cancer Res. 2007, 67, 2824–2829. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Ding, M.; Yu, M.L.; Feng, M.X.; Tan, L.J.; Zhao, F.K. Identification of galectin-7 as a potential biomarker for esophageal squamous cell carcinoma by proteomic analysis. BMC Cancer 2010, 10, 290. [Google Scholar] [CrossRef] [Green Version]
- Demers, M.; Rose, A.A.N.; Grosset, A.A.; Biron-Pain, K.; Gaboury, L.; Siegel, P.M.; St-Pierre, Y. Overexpression of galectin-7, a myoepithelial cell marker, enhances spontaneous metastasis of breast cancer cells. Am. J. Pathol. 2010, 176, 3023–3031. [Google Scholar] [CrossRef]
- Campion, C.G.; Labrie, M.; Grosset, A.A.; St-Pierre, Y. The CCAAT/enhancer-binding protein beta-2 isoform (CEBPβ-2) upregulates galectin-7 expression in human breast cancer cells. PLoS ONE 2014, 9, e95087. [Google Scholar] [CrossRef]
- Grosset, A.A.; Labrie, M.; Gagné, D.; Vladoiu, M.C.; Gaboury, L.; Doucet, N.; St-Pierre, Y. Cytosolic galectin-7 impairs p53 functions and induces chemoresistance in breast cancer cells. BMC Cancer 2014, 14, 1–10. [Google Scholar] [CrossRef]
- Grosset, A.A.; Poirier, F.; Gaboury, L.; St-Pierre, Y. Galectin-7 expression potentiates HER-2-Positive phenotype in breast cancer. PLoS ONE 2016, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Labrie, M.; Vladoiu, M.C.; Grosset, A.A.; Gaboury, L.; St-Pierre, Y. Expression and functions of galectin-7 in ovarian cancer. Oncotarget 2014, 5, 7705–7721. [Google Scholar] [CrossRef] [Green Version]
- Labrie, M.; Vladoiu, M.; Leclerc, B.G.; Grosset, A.A.; Gaboury, L.; Stagg, J.; St-Pierre, Y. A mutation in the carbohydrate recognition domain drives a phenotypic switch in the role of galectin-7 in prostate cancer. PLoS ONE 2015, 10, 1–19. [Google Scholar] [CrossRef]
- Chen, Y.S.; Chang, C.W.; Tsay, Y.G.; Huang, L.Y.; Wu, Y.C.; Cheng, L.H.; Yang, C.C.; Wu, C.H.; Teo, W.H.; Hung, K.F.; et al. HSP40 co-chaperone protein Tid1 suppresses metastasis of head and neck cancer by inhibiting Galectin-7-TCF3-MMP9 axis signaling. Theranostics 2018, 8, 3841–3855. [Google Scholar] [CrossRef]
- Guo, J.P.; Li, X.G. Galectin-7 promotes the invasiveness of human oral squamous cell carcinoma cells via activation of ERK and JNK signaling. Oncol. Lett. 2017, 13, 1919–1924. [Google Scholar] [CrossRef] [Green Version]
- Barkan, B.; Cox, A.D.; Kloog, Y. Ras inhibition boosts galectin-7 at the expense of galectin-1 to sensitize cells to apoptosis. Oncotarget 2013, 4, 256–268. [Google Scholar] [CrossRef] [Green Version]
- Kuwabara, I.; Kuwabara, Y.; Yang, R.Y.; Schuler, M.; Green, D.R.; Zuraw, B.L.; Hsu, D.K.; Liu, F.T. Galectin-7 (PIG1) exhibits pro-apoptotic function through JNK activation and mitochondrial cytochrome c release. J. Biol. Chem. 2002, 277, 3487–3497. [Google Scholar] [CrossRef] [Green Version]
- Ueda, S.; Kuwabara, I.; Liu, F.-T. Suppression of tumor growth by the β4-galactosyltransferase gene. Cancer Res. 2004, 64, 5672–5676. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Wu, T.C.; Chen, W.Q.; Zhou, L.J.; Wu, Y.; Zeng, L.; Pei, H.P. Roles of galectin-7 and S100A9 in cervical squamous carcinoma: Clinicopathological and in vitro evidence. Int. J. Cancer 2013, 132, 1051–1059. [Google Scholar] [CrossRef]
- Higareda-Almaraz, J.C.; Ruiz-Moreno, J.S.; Klimentova, J.; Barbieri, D.; Salvador-Gallego, R.; Ly, R.; Valtierra-Gutierrez, I.A.; Dinsart, C.; Rabinovich, G.A.; Stulik, J.; et al. Systems-level effects of ectopic galectin-7 reconstitution in cervical cancer and its microenvironment. BMC Cancer 2016, 16, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Menkhorst, E.; Griffiths, M.; van Sinderen, M.; Rainczuk, K.; Niven, K.; Dimitriadis, E. Galectin-7 is elevated in endometrioid (type I) endometrial cancer and promotes cell migration. Oncol. Lett. 2018, 16, 4721–4728. [Google Scholar] [CrossRef] [Green Version]
- Matsui, Y.; Ueda, S.; Watanabe, J.; Kuwabara, I.; Ogawa, O.; Nishiyama, H. Sensitizing effect of galectin-7 in urothelial cancer to cisplatin through the accumulation of intracellular reactive oxygen species. Cancer Res. 2007, 67, 1212–1220. [Google Scholar] [CrossRef] [Green Version]
- Kopitz, J.; André, S.; Von Reitzenstein, C.; Versluis, K.; Kaltner, H.; Pieters, R.J.; Wasano, K.; Kuwabara, I.; Liu, F.T.; Cantz, M.; et al. Homodimeric galectin-7 (p53-induced gene 1) is a negative growth regulator for human neuroblastoma cells. Oncogene 2003, 22, 6277–6288. [Google Scholar] [CrossRef] [Green Version]
- Ives, C.W.; Sinkey, R.; Rajapreyar, I.; Tita, A.T.N.; Oparil, S. Preeclampsia—Pathophysiology and Clinical Presentations: JACC State-of-the-Art Review. J. Am. Coll. Cardiol. 2020, 76, 1690–1702. [Google Scholar] [CrossRef]
- Phipps, E.A.; Thadhani, R.; Benzing, T.; Karumanchi, S.A. Pre-eclampsia: Pathogenesis, novel diagnostics and therapies. Nat. Rev. Nephrol. 2019, 15, 275–289. [Google Scholar] [CrossRef]
- Larsen, E.C.; Christiansen, O.B.; Kolte, A.M.; Macklon, N. New insights into mechanisms behind miscarriage. BMC Med. 2013, 11, 154. [Google Scholar] [CrossRef] [Green Version]
- Dimitriadis, E.; Menkhorst, E.; Saito, S.; Kutteh, W.H.; Brosens, J.J. Recurrent pregnancy loss. Nat. Rev. Dis. Prim. 2020, 6, 98. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, U.; Hutter, S.; Heublein, S.; Vrekoussis, T.; Andergassen, U.; Unverdorben, L.; Papadakis, G.; Makrigiannakis, A. Expression and function of galectins in the endometrium and at the human feto-maternal interface. Placenta 2013, 34, 863–872. [Google Scholar] [CrossRef]
- Blois, S.M.; Barrientos, G. Galectin signature in normal pregnancy and preeclampsia. J. Reprod. Immunol. 2014, 101–102, 127–134. [Google Scholar] [CrossRef]
- Evans, J.; Yap, J.; Gamage, T.; Salamonsen, L.; Dimitriadis, E.; Menkhorst, E. Galectin-7 is important for normal uterine repair following menstruation. Mol. Hum. Reprod. 2014, 20, 787–798. [Google Scholar] [CrossRef] [PubMed]
- Menkhorst, E.; Koga, K.; Van Sinderen, M.; Dimitriadis, E. Galectin-7 serum levels are altered prior to the onset of pre-eclampsia. Placenta 2014, 35, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Unverdorben, L.; Haufe, T.; Santoso, L.; Hofmann, S.; Jeschke, U.; Hutter, S. Prototype and chimera-type galectins in placentas with spontaneous and recurrent miscarriages. Int. J. Mol. Sci. 2016, 17, 644. [Google Scholar] [CrossRef] [Green Version]
- Menkhorst, E.; Zhou, W.; Santos, L.L.; Delforce, S.; So, T.; Rainczuk, K.; Loke, H.; Syngelaki, A.; Varshney, S.; Williamson, N.; et al. Galectin-7 impairs placentation and causes preeclampsia features in mice. Hypertension 2020, 76, 1185–1194. [Google Scholar] [CrossRef]
- Xu, W.D.; Huang, Q.; Huang, A.F. Emerging role of galectin family in inflammatory autoimmune diseases. Autoimmun. Rev. 2021, 20, 102847. [Google Scholar] [CrossRef]
- Wan, L.; Hsu, Y.A.; Wei, C.C.; Liu, F.T. Galectins in allergic inflammatory diseases. Mol. Aspects Med. 2021, 79, 100925. [Google Scholar] [CrossRef]
- Niiyama, S.; Yoshino, T.; Yasuda, C.; Yu, X.; Izumi, R.; Ishiwatari, S.; Matsukuma, S.; Mukai, H. Galectin-7 in the stratum corneum: A biomarker of the skin barrier function. Int. J. Cosmet. Sci. 2016, 38, 487–495. [Google Scholar] [CrossRef]
- Umayahara, T.; Shimauchi, T.; Iwasaki, M.; Sakabe, J.i.; Aoshima, M.; Nakazawa, S.; Yatagai, T.; Yamaguchi, H.; Phadungsaksawasdi, P.; Kurihara, K.; et al. Protective role of Galectin-7 for skin barrier impairment in atopic dermatitis. Clin. Exp. Allergy 2020, 922–931. [Google Scholar] [CrossRef]
- Luo, Z.; Ji, Y.; Tian, D.; Zhang, Y.; Chang, S.; Yang, C.; Zhou, H.; Chen, Z.K. Galectin-7 promotes proliferation and Th1/2 cells polarization toward Th1 in activated CD4+ T cells by inhibiting The TGFβ/Smad3 pathway. Mol. Immunol. 2018, 101, 80–85. [Google Scholar] [CrossRef]
- Tian, J.; He, R.; Fan, Y.; Zhang, Q.; Tian, B.; Zhou, C.; LIU, C.; Song, M.; Zhao, S. Galectin-7 overexpression destroy airway epithelial barrier in transgenic mice. Integr. Zool. 2020, 16, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Zhang, W. Silencing of Gal-7 inhibits TGF-β1-induced apoptosis of human airway epithelial cells through JNK signaling pathway. Exp. Cell Res. 2019, 375, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Nozawa, T.; Minowa-Nozawa, A.; Toh, H.; Hikichi, M.; Iibushi, J.; Nakagawa, I. Autophagy Receptor Tollip Facilitates Bacterial Autophagy by Recruiting Galectin-7 in Response to Group A Streptococcus Infection. Front. Cell. Infect. Microbiol. 2020, 10, 583137. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Ji, Y.; Zhou, H.; Huang, X.; Fang, J.; Guo, H.; Pan, T.; Chen, Z.K. Galectin-7 in cardiac allografts in mice: Increased expression compared with isografts and localization in infiltrating lymphocytes and vascular endothelial cells. Transplant. Proc. 2013, 45, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Dings, R.P.M.; Miller, M.C.; Griffin, R.J.; Mayo, K.H. Galectins as molecular targets for therapeutic intervention. Int. J. Mol. Sci. 2018, 19, 905. [Google Scholar] [CrossRef] [Green Version]
- Stegmayr, J.; Zetterberg, F.; Carlsson, M.C.; Huang, X.; Sharma, G.; Kahl-Knutson, B.; Schambye, H.; Nilsson, U.J.; Oredsson, S.; Leffler, H. Extracellular and intracellular small-molecule galectin-3 inhibitors. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Chatenet, D.; Doucet, N.S.; Pierre, Y. Galectin-7-specific monovalent antibodies and uses thereof. International Application No. PCT/CA2020/050024, 1 September 2020. [Google Scholar]
- Grosset, A.A.; Labrie, M.; Vladoiu, M.C.; Yousef, E.M.; Gaboury, L.; St-Pierre, Y. Galectin signatures contribute to the heterogeneity of breast cancer and provide new prognostic information and therapeutic targets. Oncotarget 2016, 7, 18183–18203. [Google Scholar] [CrossRef] [Green Version]
- Bibens-Laulan, N.; St-Pierre, Y. Intracellular galectin-7 expression in cancer cells results from an autocrine transcriptional mechanism and endocytosis of extracellular galectin-7. PLoS ONE 2017, 12, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Vladoiu, M.C.; Labrie, M.; St-Pierre, Y. Intracellular galectins in cancer cells: Potential new targets for therapy (review). Int. J. Oncol. 2014, 44, 1001–1014. [Google Scholar] [CrossRef] [Green Version]
- Hama, N.; Nishimura, K.; Hasegawa, A.; Yuki, A.; Kume, H.; Adachi, J.; Kinoshita, M.; Ogawa, Y.; Nakajima, S.; Nomura, T.; et al. Galectin-7 as a potential biomarker of Stevens-Johnson syndrome/toxic epidermal necrolysis: Identification by targeted proteomics using causative drug-exposed peripheral blood cells. J. Allergy Clin. Immunol. Pract. 2019, 7, 2894–2897.e7. [Google Scholar] [CrossRef]
- Kim, H.J.; Jeon, H.K.; Lee, J.K.; Sung, C.O.; Do, I.G.; Choi, C.H.; Kim, T.J.; Kim, B.G.; Bae, D.S.; Lee, J.W. Clinical significance of galectin-7 in epithelial ovarian cancer. Anticancer Res. 2013, 33, 1555–1562. [Google Scholar]
- Schulz, H.; Schmoeckel, E.; Kuhn, C.; Hofmann, S.; Mayr, D.; Mahner, S.; Jeschke, U. Galectins-1, -3, and-7 are prognostic markers for survival of ovarian cancer patients. Int. J. Mol. Sci. 2017, 18, 1230. [Google Scholar] [CrossRef] [Green Version]
- Trebo, A.; Ditsch, N.; Kuhn, C.; Heidegger, H.H.; Zeder-Goess, C.; Kolben, T.; Czogalla, B.; Schmoeckel, E.; Mahner, S.; Jeschke, U.; et al. High galectin-7 and low galectin-8 expression and the combination of both are negative prognosticators for breast cancer patients. Cancers 2020, 12, 953. [Google Scholar] [CrossRef] [Green Version]
- Matsukawa, S.; Morita, K.i.; Negishi, A.; Harada, H.; Nakajima, Y.; Shimamoto, H.; Tomioka, H.; Tanaka, K.; Ono, M.; Yamada, T.; et al. Galectin-7 as a potential predictive marker of chemo- and/or radio-therapy resistance in oral squamous cell carcinoma. Cancer Med. 2014, 3, 349–361. [Google Scholar] [CrossRef]
- Modenutti, C.P.; Capurro, J.I.B.; Di Lella, S.; Martí, M.A. The Structural Biology of Galectin-Ligand Recognition: Current Advances in Modeling Tools, Protein Engineering, and Inhibitor Design. Front. Chem. 2019, 7, 823. [Google Scholar] [CrossRef]
- Ermakova, E.; Miller, M.C.; Nesmelova, I.V.; López-Merino, L.; Berbís, M.A.; Nesmelov, Y.; Tkachev, Y.V.; Lagartera, L.; Daragan, V.A.; André, S.; et al. Lactose binding to human galectin-7 (p53-induced gene 1) induces long-range effects through the protein resulting in increased dimer stability and evidence for positive cooperativity. Glycobiology 2013, 23, 508–523. [Google Scholar] [CrossRef]
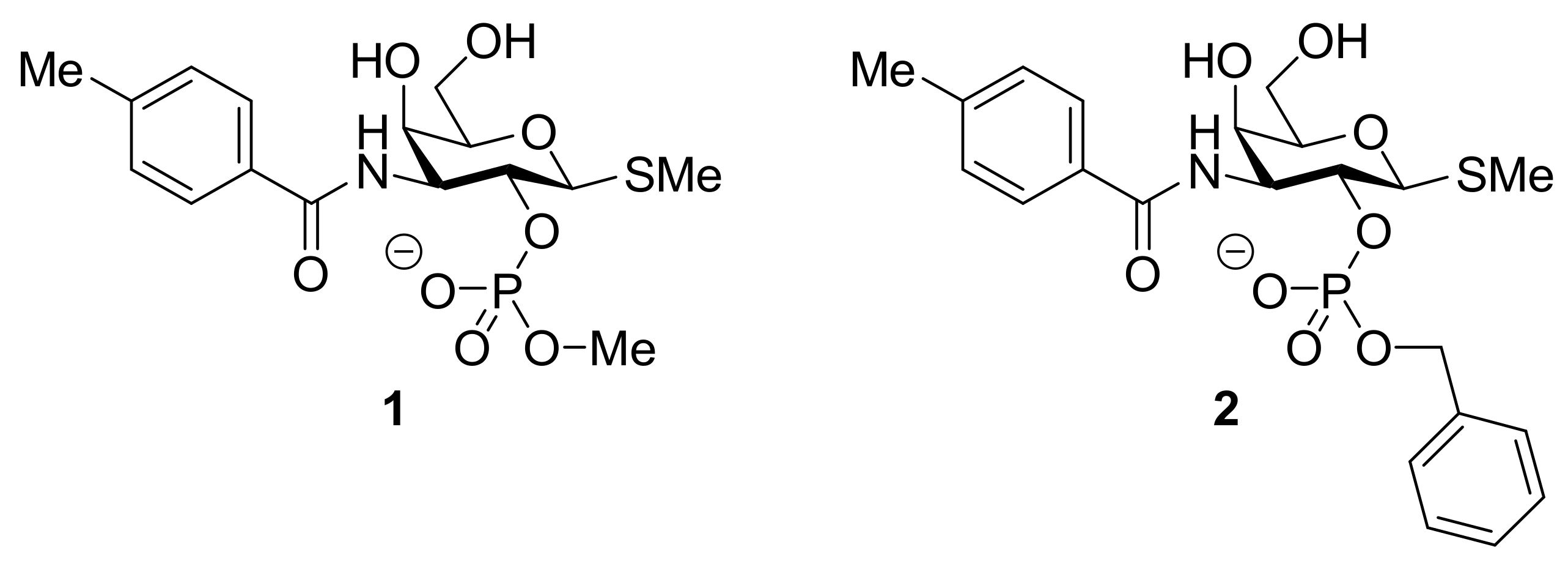
- Masuyer, G.; Jabeen, T.; Öberg, C.T.; Leffler, H.; Nilsson, U.J.; Acharya, K.R. Inhibition mechanism of human galectin-7 by a novel galactose- benzylphosphate inhibitor. FEBS J. 2012, 279, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, T.J.; Lin, H.Y.; Tu, Z.; Huang, B.S.; Wu, S.C.; Lin, C.H. Structural basis underlying the binding preference of human galectins-1, -3 and -7 for Galβ1-3/4GlcNAc. PLoS ONE 2015, 10, 1–19. [Google Scholar] [CrossRef]
- Ramaswamy, S.; Sleiman, M.H.; Masuyer, G.; Arbez-Gindre, C.; Micha-Screttas, M.; Calogeropoulou, T.; Steele, B.R.; Acharya, K.R. Structural basis of multivalent galactose-based dendrimer recognition by human galectin-7. FEBS J. 2015, 282, 372–387. [Google Scholar] [CrossRef]
- MacKinnon, A.C.; Gibbons, M.A.; Farnworth, S.L.; Leffler, H.; Nilsson, U.J.; Delaine, T.; Simpson, A.J.; Forbes, S.J.; Hirani, N.; Gauldie, J.; et al. Regulation of transforming growth factor-β1-driven lung fibrosis by galectin-3. Am. J. Respir. Crit. Care Med. 2012, 185, 537–546. [Google Scholar] [CrossRef] [Green Version]
- Garber, K. Galecto Biotech. Nat. Biotechnol. 2013, 31, 481–482. [Google Scholar] [CrossRef]
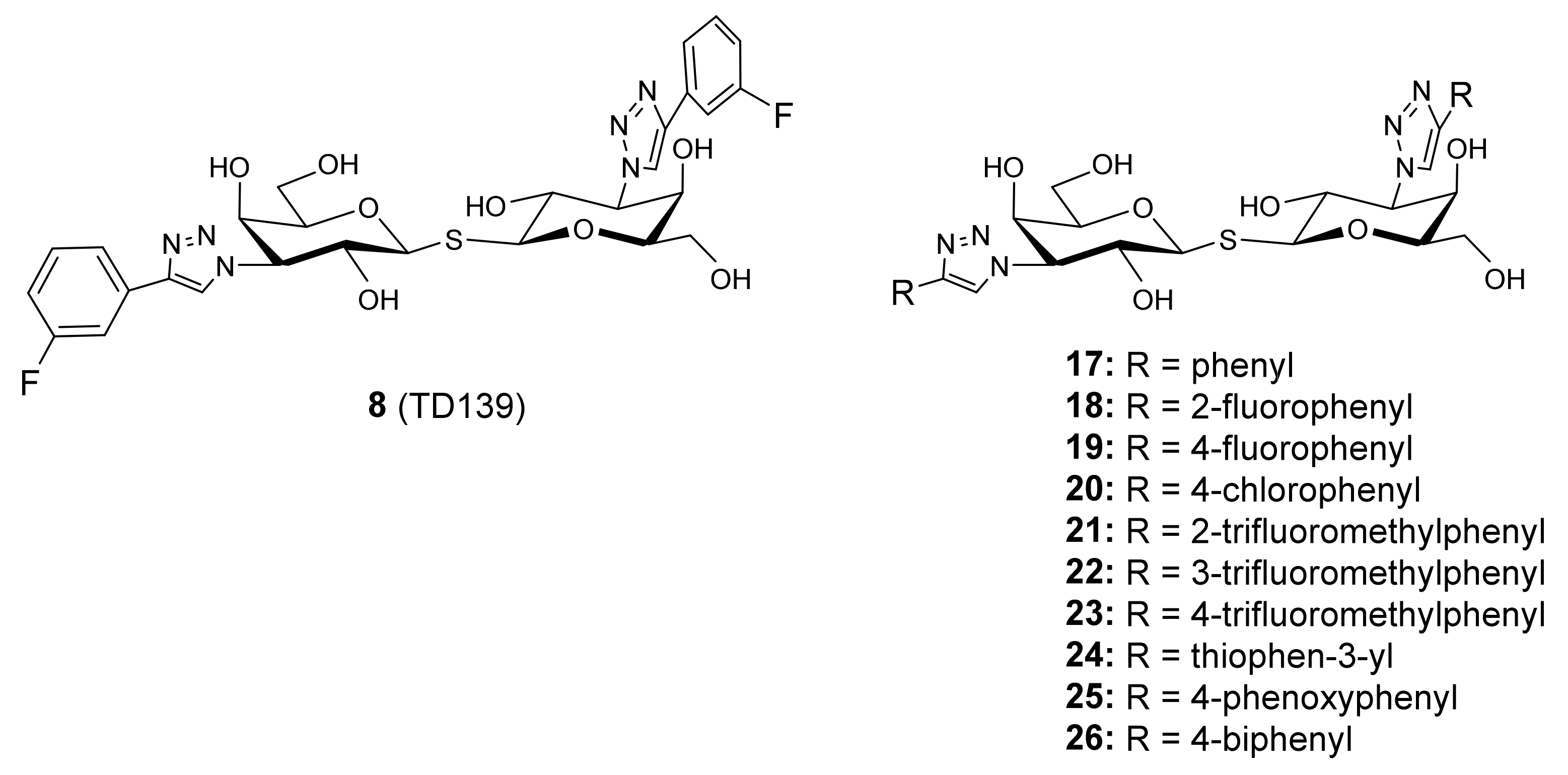
- Hirani, N.; MacKinnon, A.C.; Nicol, L.; Ford, P.; Schambye, H.; Pedersen, A.; Nilsson, U.J.; Leffler, H.; Sethi, T.; Tantawi, S.; et al. Target inhibition of galectin-3 by inhaled TD139 in patients with idiopathic pulmonary fibrosis. Eur. Respir. J. 2021, 57, 1–13. [Google Scholar] [CrossRef]
- Delaine, T.; Collins, P.; Mackinnon, A.; Sharma, G.; Stegmayr, J.; Rajput, V.K.; Mandal, S.; Cumpstey, I.; Larumbe, A.; Salameh, B.A.; et al. Galectin-3-Binding Glycomimetics that Strongly Reduce Bleomycin-Induced Lung Fibrosis and Modulate Intracellular Glycan Recognition. ChemBioChem 2016, 17, 1759–1770. [Google Scholar] [CrossRef]
- Hsieh, T.J.; Lin, H.Y.; Tu, Z.; Lin, T.C.; Wu, S.C.; Tseng, Y.Y.; Liu, F.T.; Hsu, S.T.D.; Lin, C.H. Dual thio-digalactoside-binding modes of human galectins as the structural basis for the design of potent and selective inhibitors. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- MacKinnon, A.; Chen, W.-S.; Leffler, H.; Panjwani, N.; Schambye, H.; Sethi, T.; Nilsson, U.J. Design, Synthesis, and Applications of Galectins Modulators in Human Health; Seeberger, P.H., Rademacher, C., Eds.; Springer International Publishing: Cham, Switzerland, 2014; Volume 12, ISBN 978-3-319-08674-3. [Google Scholar]
- Chan, Y.C.; Lin, H.Y.; Tu, Z.; Kuo, Y.H.; Hsu, S.T.D.; Lin, C.H. Dissecting the structure–Activity relationship of galectin–Ligand interactions. Int. J. Mol. Sci. 2018, 19, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
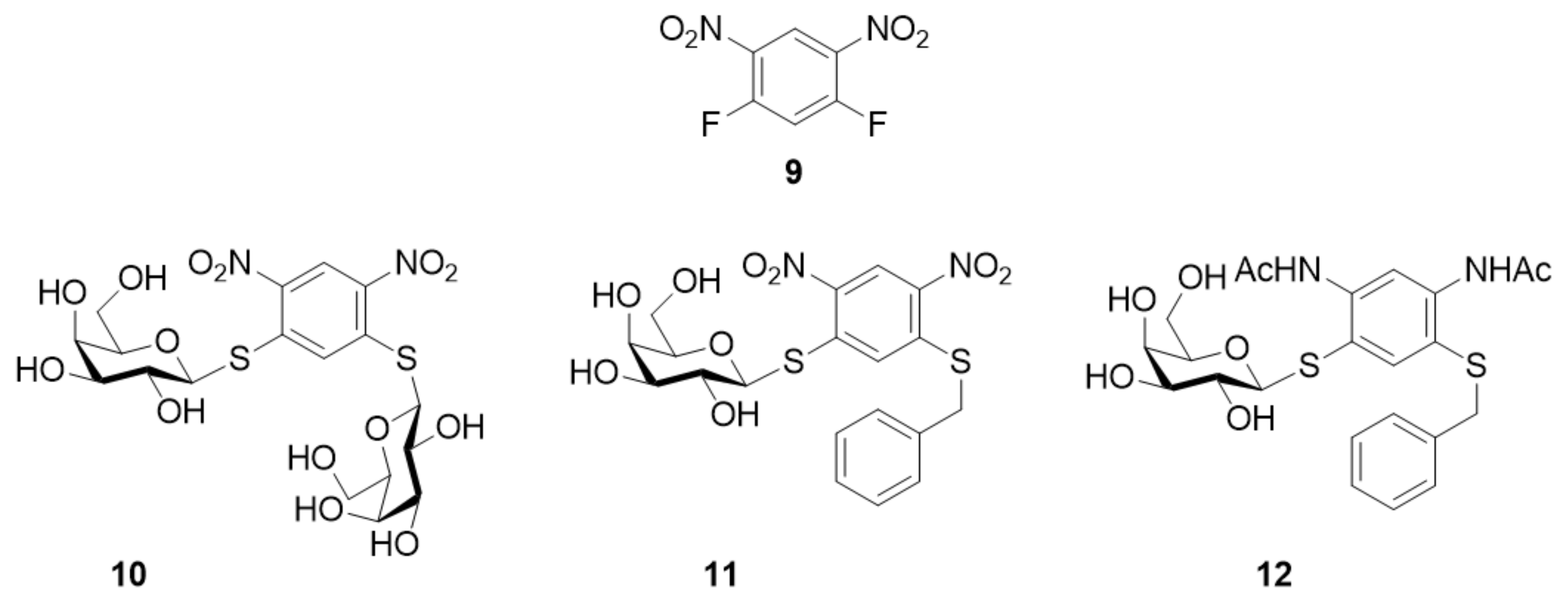
- Cumpstey, I.; Carlsson, S.; Leffler, H.; Nilsson, U.J. Synthesis of a phenyl thio-√ü-D-galactopyranoside library from 1,5-difluoro-2,4-dinitrobenzene: Discovery of efficient and selective monosaccharide inhibitors of galectin-7. Org. Biomol. Chem. 2005, 3, 1922–1932. [Google Scholar] [CrossRef] [PubMed]
- Bergh, A.; Leffler, H.; Sundin, A.; Nilsson, U.J.; Kann, N. Cobalt-mediated solid phase synthesis of 3-O-alkynylbenzyl galactosides and their evaluation as galectin inhibitors. Tetrahedron 2006, 62, 8309–8317. [Google Scholar] [CrossRef]
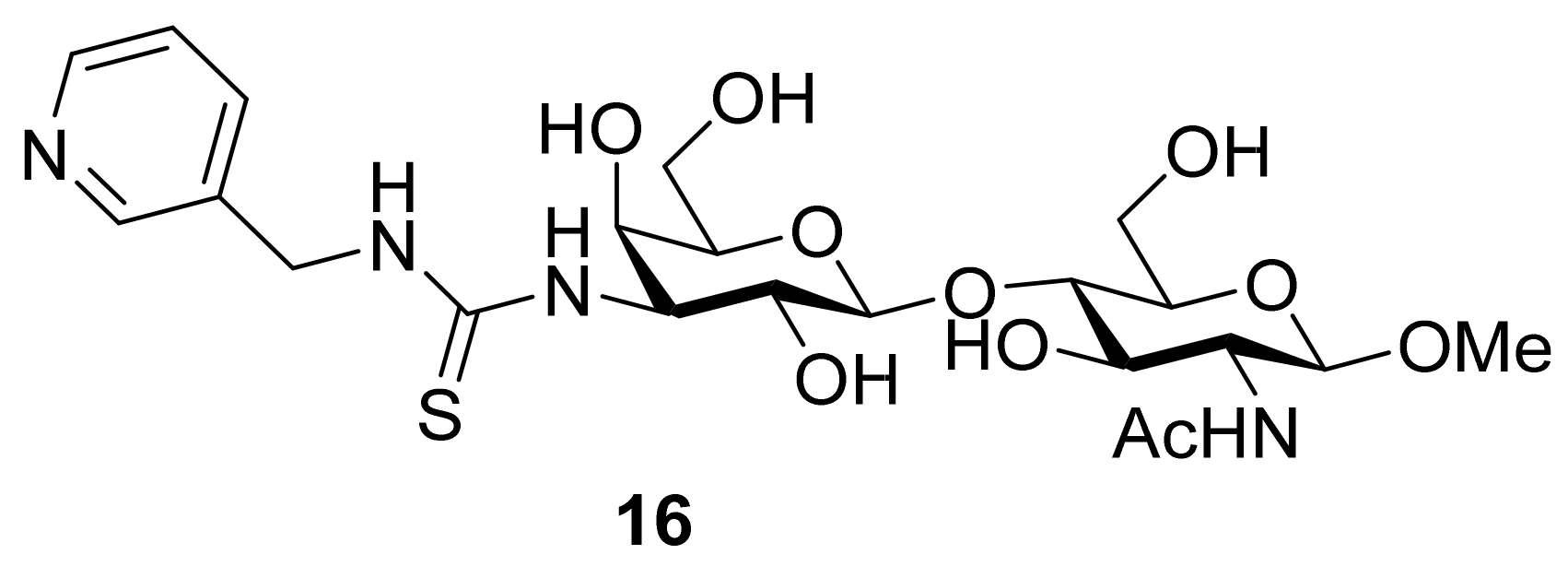
- Salameh, B.A.; Sundin, A.; Leffler, H.; Nilsson, U.J. Thioureido N-acetyllactosamine derivatives as potent galectin-7 and 9N inhibitors. Bioorganic Med. Chem. 2006, 14, 1215–1220. [Google Scholar] [CrossRef]
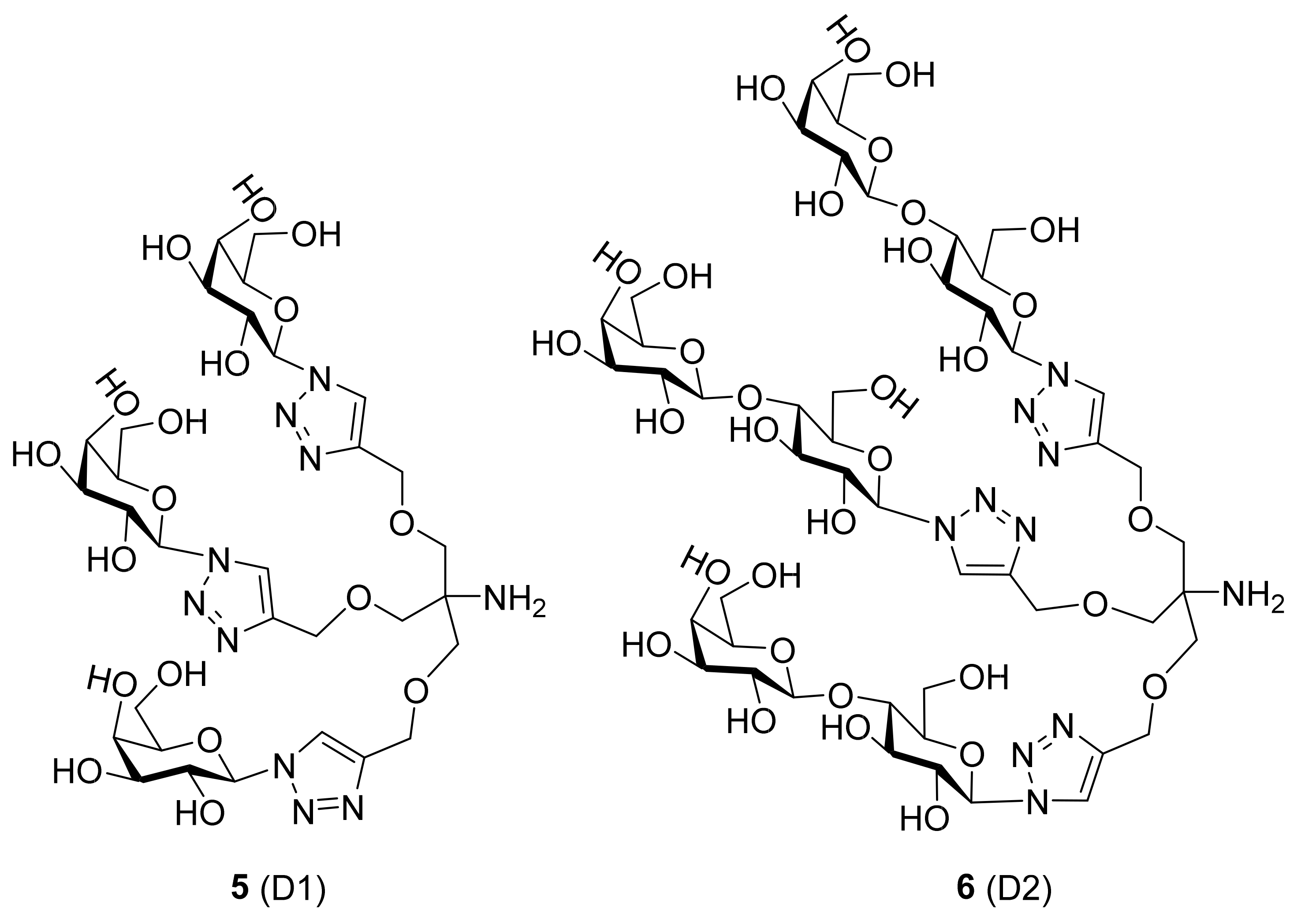
- Vladoiu, M.C.; Labrie, M.; Létourneau, M.; Egesborg, P.; Gagné, D.; Billard, É.; Grosset, A.A.; Doucet, N.; Chatenet, D.; St-Pierre, Y. Design of a peptidic inhibitor that targets the dimer interface of a prototypic galectin. Oncotarget 2015, 6, 40970–40980. [Google Scholar] [CrossRef]
- López De Los Santos, Y.; Bernard, D.N.; Egesborg, P.; Létourneau, M.; Lafortune, C.; Cuneo, M.J.; Urvoas, A.; Chatenet, D.; Mahy, J.P.; St-Pierre, Y.; et al. Binding of a Soluble meso-Tetraarylporphyrin to Human Galectin-7 Induces Oligomerization and Modulates Its Pro-Apoptotic Activity. Biochemistry 2020, 59, 4591–4600. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Role | Mode of Action | References |
|---|---|---|
| Epidermal homeostasis of skin | Regulation of keratinocyte proliferation, differentiation and migration | [14,15,16,17,18,19,20,21,22,23,24,25] |
| Re-epithelialization of corneal wounds | Mediating corneal epithelial cell migration | [27] |
| Wound healing of PDL fibroblasts | Promoting proliferation, migration and invasion of PDL fibroblasts | [28] |
| Promalignant activity in gastric cancer | Lower expression levels of galectin-7 cause increase in gastric cancer cell proliferation, migration and invasion | [54] |
| Promalignant activity in thymic lymphoma + HNSCC | Induce MMP-9 expression | [6,55,56,64] |
| Increasing invasive behavior of breast cancer cells | Protecting breast cancer cells from apoptosis | [58,59,60,61] |
| Reducing invasive behavior of prostate cancer cells | Inhibiting motility prostate cancer cells | [63] |
| Pro-invasive activity in oral squamous cell carcinoma | Induce MMP-2 and MMP-9 expression | [65,66] |
| Protective effect on the survival of cervical squamous carcinoma patients | Inhibiting MMP-9 expression and cell invasion in cervical squamous carcinoma cells | [67,69,70] |
| Promoting metastasis of endometrial cancer | Reducing cell–cell adhesion and enhancing cell migration | [71] |
| Sensitizing bladder cancer cells to chemotherapy | Increase generation of reactive oxygen species | [72] |
| Negative growth regulator of neuroblastoma cells | Switch from proliferation to differentiation of cancer cells | [73] |
| Mediation of endometrial epithelial wound repair | Endometrial re-epithelialization is dependent on integrin mediated signaling | [80] |
| Abnormal placentation hence leading to the development of pre-eclampsia | Acting via the placenta to induce the systemic features of pre-eclampsia via impaired placental formation, placental inflammation and placental release of anti-angiogenic factors | [81,83] |
| Skin barrier impairment in keratinocytes | Protecting disruption of cell-to-cell adhesion and/or cell-to-extracellular matrix adhesion | [87] |
| Anti-inflammation effects, inducing autoimmune disease and transplantation rejection | Promotion, proliferation and polarization of Th1/2 cells | [88] |
| Causing airway structural defects, injury, and other asthma responses | Increased apoptosis occurred in bronchial epithelial cells in asthma | [89,90] |
| Intracellular immunity in the response against bacterial infection | Colocalizing with and surrounding group A Streptococcus (GAS, intracellular bacterium) | [91] |
| Accelerating allograft rejection | Up-regulation of galectin-7 expression in the allografts was directly related to T cell response | [92] |
| Compound | Cross-Linked Form |
|---|---|
| 5 (D1) |  |
| 6 (D2) |  |
| 6 (D2) |  |
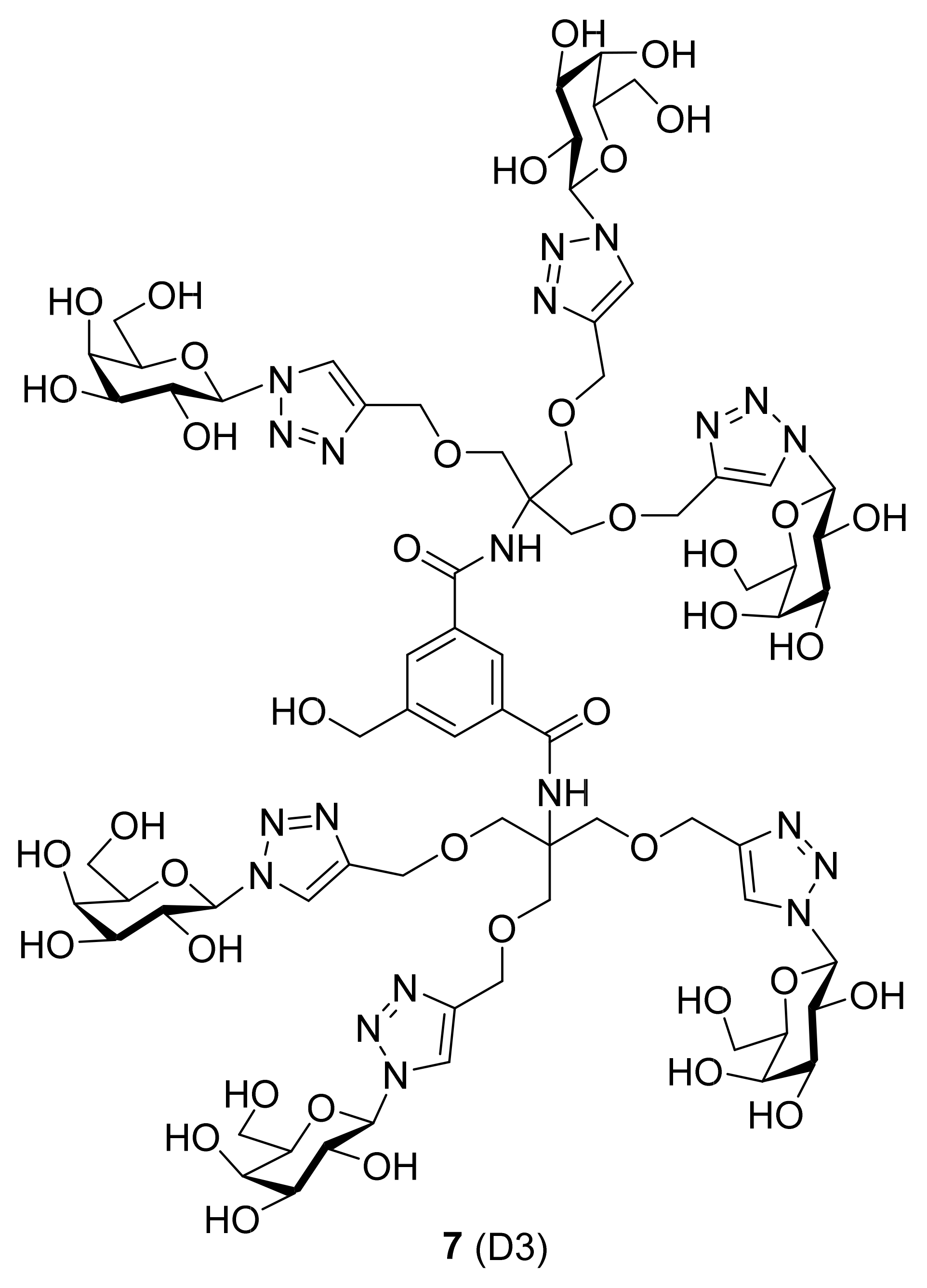
| 7 (D3) |  |
| Compound | Galectin-1 | Galectin-3 | Galectin-7 | Galectin-8N | Galectin-9N |
|---|---|---|---|---|---|
 | 27 | 2.4 | 0.39 | 1.0 | 1.0 |
 | 6.9 | 2.9 | 0.65 | 3.8 | 1.9 |
 | n.i. a | 5.4 | 0.74 | 2.4 | 2.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sewgobind, N.V.; Albers, S.; Pieters, R.J. Functions and Inhibition of Galectin-7, an Emerging Target in Cellular Pathophysiology. Biomolecules 2021, 11, 1720. https://doi.org/10.3390/biom11111720
Sewgobind NV, Albers S, Pieters RJ. Functions and Inhibition of Galectin-7, an Emerging Target in Cellular Pathophysiology. Biomolecules. 2021; 11(11):1720. https://doi.org/10.3390/biom11111720
Chicago/Turabian StyleSewgobind, Nishant V., Sanne Albers, and Roland J. Pieters. 2021. "Functions and Inhibition of Galectin-7, an Emerging Target in Cellular Pathophysiology" Biomolecules 11, no. 11: 1720. https://doi.org/10.3390/biom11111720