
Auto-Regulation of Transcription and Translation: Oscillations, Excitability and Intermittency


Abstract
:1. Introduction
2. Materials and Methods
2.1. Model Development
2.2. Parameters
2.3. Bifurcation Analysis
2.4. Stochastic Model
3. Results
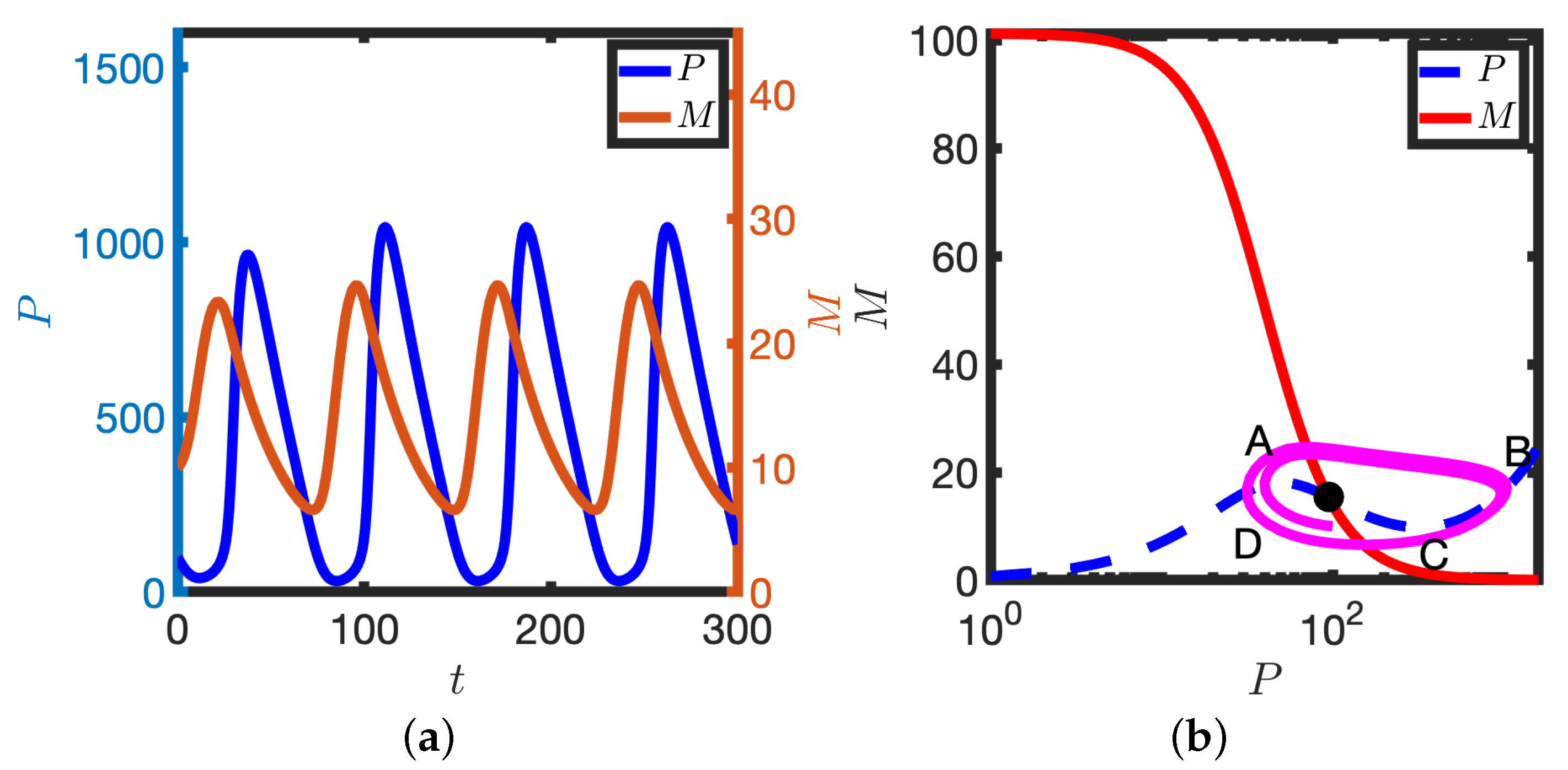
3.1. Exploring Model Behaviour
3.2. Experimentally Testable Predictions
3.2.1. Inhibition of Transcription via a Notch Signalling Inhibitor
3.2.2. Treatment with a Translational Inhibitor
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Van Es, J.H.; Van Gijn, M.E.; Riccio, O.; Van Den Born, M.; Vooijs, M.; Begthel, H.; Cozijnsen, M.; Robine, S.; Winton, D.J.; Radtke, F.; et al. Notch/γ-secretase inhibition turns proliferative cells in intestinal crypts and adenomas into goblet cells. Nature 2005, 435, 959–963. [Google Scholar] [CrossRef] [PubMed]
- Allenspach, E.J.; Maillard, I.; Aster, J.C.; Pear, W.S. Notch signaling in cancer. Cancer Biol. Ther. 2002, 1, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Siebel, C.; Lendahl, U. Notch signaling in development, tissue homeostasis, and disease. Physiol. Rev. 2017, 97, 1235–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, S.J. Notch signalling: A simple pathway becomes complex. Nat. Rev. Mol. Cell Biol. 2006, 7, 678–689. [Google Scholar] [CrossRef]
- Fischer, A.; Gessler, M. Delta-Notch– and then? Protein interactions and proposed modes of repression by Hes and Hey bHLH factors. Nucleic Acids Res. 2007, 35, 4583–4596. [Google Scholar] [CrossRef] [Green Version]
- Novák, B.; Tyson, J.J. Design principles of biochemical oscillators. Nat. Rev. Mol. Cell Biol. 2008, 9, 981–991. [Google Scholar] [CrossRef] [Green Version]
- Goodwin, B.C. Oscillatory behavior in enzymatic control processes. Adv. Enzym. Regul. 1965, 3, 425–437. [Google Scholar] [CrossRef]
- Monk, N.A. Oscillatory expression of Hes1, p53, and NF-κB driven by transcriptional time delays. Curr. Biol. 2003, 13, 1409–1413. [Google Scholar] [CrossRef] [Green Version]
- Lewis, J. Autoinhibition with Transcriptional Delay. Curr. Biol. 2003, 13, 1398–1408. [Google Scholar] [CrossRef] [Green Version]
- Sturrock, M.; Terry, A.J.; Xirodimas, D.P.; Thompson, A.M.; Chaplain, M.A. Spatio-temporal modelling of the Hes1 and p53-Mdm2 intracellular signalling pathways. J. Theor. Biol. 2011, 273, 15–31. [Google Scholar] [CrossRef] [Green Version]
- Chaplain, M.; Ptashnyk, M.; Sturrock, M. Hopf bifurcation in a gene regulatory network model: Molecular movement causes oscillations. Math. Model. Methods Appl. Sci. 2015, 25, 1179–1215. [Google Scholar] [CrossRef] [Green Version]
- Tyson, J.J.; Novak, B. Regulation of the eukaryotic cell cycle: Molecular antagonism, hysteresis, and irreversible transitions. J. Theor. Biol. 2001, 210, 249–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodgkin, A.L.; Huxley, A.F. A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 1952, 117, 500–544. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.Y.C.; Choi, Y.S.; Ma, W.; Pomerening, J.R.; Tang, C.; Ferrell, J.E. Robust, tunable biological oscillations from interlinked positive and negative feedback loops. Science 2008, 321, 126–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonev, B.; Stanley, P.; Papalopulu, N. MicroRNA-9 modulates Hes1 ultradian oscillations by forming a double-negative feedback loop. Cell Rep. 2012, 2, 10–18. [Google Scholar] [CrossRef]
- Goodfellow, M.; Phillips, N.E.; Manning, C.; Galla, T.; Papalopulu, N. microRNA input into a neural ultradian oscillator controls emergence and timing of alternative cell states. Nat. Commun. 2014, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Cibois, M.; Gautier-Courteille, C.; Legagneux, V.; Paillard, L. Post-transcriptional controls—Adding a new layer of regulation to clock gene expression. Trends Cell Biol. 2010, 20, 533–541. [Google Scholar] [CrossRef] [Green Version]
- Dankowicz, H.; Schilder, F. Recipes for Continuation; SIAM: Philadelphia, PA, USA, 2013. [Google Scholar]
- Webb, A.B.; Lengyel, I.M.; Jörg, D.J.; Valentin, G.; Jülicher, F.; Morelli, L.G.; Oates, A.C. Persistence, period and precision of autonomous cellular oscillators from the zebrafish segmentation clock. eLife 2016, 5, e08438. [Google Scholar] [CrossRef]
- Wiedermann, G.; Bone, R.A.; Silva, J.C.; Bjorklund, M.; Murray, P.J.; Dale, J.K. A balance of positive and negative regulators determines the pace of the segmentation clock. eLife 2015, 4. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka-Kobayashi, K.; Matsumiya, M.; Niino, Y.; Isomura, A.; Kori, H.; Miyawaki, A.; Kageyama, R. Coupling delay controls synchronized oscillation in the segmentation clock. Nature 2020, 580, 119–123. [Google Scholar] [CrossRef]
- Fitzhugh, R. Thresholds and plateaus in the Hodgkin-Huxley nerve equations. J. Gen. Physiol. 1960, 43, 867–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagumo, J.; Arimoto, S.; Yoshizawa, S. An active pulse transmission line simulating nerve axon. Proc. IRE 1962, 50, 2061–2070. [Google Scholar] [CrossRef]
- Hubaud, A.; Regev, I.; Mahadevan, L.; Pourquie, O. Excitable dynamics and Yap-dependent mechanical cues drive the segmentation clock. Cell 2017, 171, 668–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrgen, L.; Ares, S.; Morelli, L.G.; Schröter, C.; Jülicher, F.; Oates, A.C. Intercellular coupling regulates the period of the segmentation clock. Curr. Biol. 2010, 20, 1244–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, B.K.; Jörg, D.J.; Oates, A.C. Faster embryonic segmentation through elevated Delta-Notch signalling. Nat. Commun. 2016, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Description | Value | Units |
|---|---|---|---|
| Maximal transcription rate | 3.03 | min | |
| mRNA degradation rate | 0.03 | min | |
| Background translation rate | 0.12 | min | |
| X dependent translation rate | 20.12 | min | |
| P degradation rate | 0.30 | min | |
| Max. X | 70.43 | Nondim | |
| Translation IC50 | 1 | Nondim | |
| Transcription IC50 | 40 | Nondim | |
| Protein noise strength | 1.0 | min | |
| mRNA noise strength | 0.12 | min | |
| mRNA initial condition | 6 | Nondim | |
| protein initial condition | 60 | Nondim |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murray, P.J.; Ocana, E.; Meijer, H.A.; Dale, J.K. Auto-Regulation of Transcription and Translation: Oscillations, Excitability and Intermittency. Biomolecules 2021, 11, 1566. https://doi.org/10.3390/biom11111566
Murray PJ, Ocana E, Meijer HA, Dale JK. Auto-Regulation of Transcription and Translation: Oscillations, Excitability and Intermittency. Biomolecules. 2021; 11(11):1566. https://doi.org/10.3390/biom11111566
Chicago/Turabian StyleMurray, Philip J., Eleonore Ocana, Hedda A. Meijer, and Jacqueline Kim Dale. 2021. "Auto-Regulation of Transcription and Translation: Oscillations, Excitability and Intermittency" Biomolecules 11, no. 11: 1566. https://doi.org/10.3390/biom11111566