Sarocladium and Lecanicillium Associated with Maize Seeds and Their Potential to Form Selected Secondary Metabolites

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Collection
2.2. Fungal Isolation and Identification
2.3. Chemicals and Reagents
2.4. Secondary Metabolite Production
2.5. Sample Extraction and HPLC Analysis of Enns and BEA
2.6. Qualitative Analysis of Selected Metabolites Formed by L. lecanii, S. zeae and S. strictum Isolates Using UPLC/TQD
3. Results and Discussion
3.1. Isolation and Identification or Re-Identification of Sarocladium and Lecanicillium Species Associated with Maize Seeds
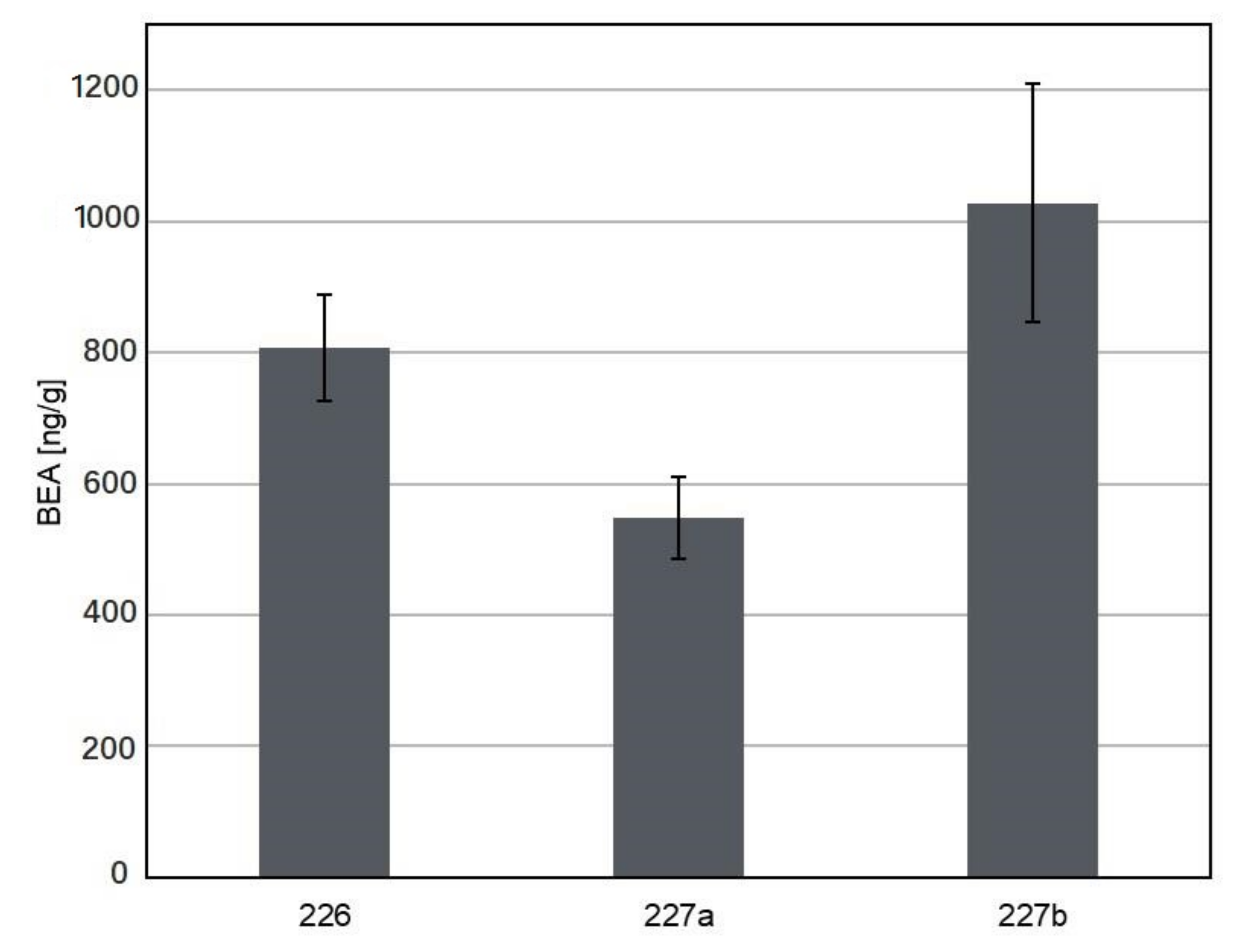
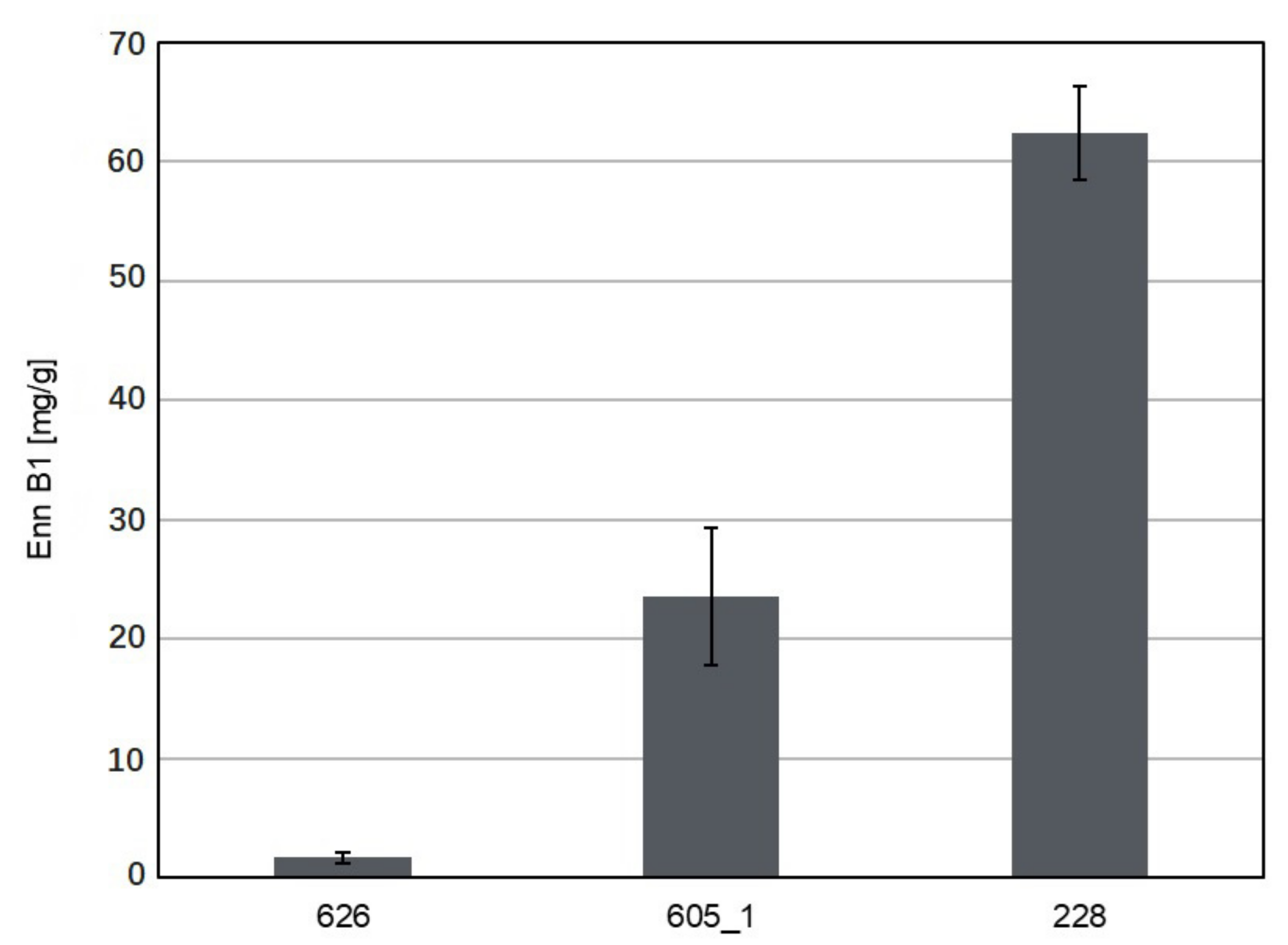
3.2. Production of Mycotoxins and Other Compounds by the Investigated Sarocladium and Lecanicilium Isolates
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abe, C.A.; Faria, C.B.; de Castro, F.F.; de Souza, S.R.; dos Santos, F.C.; da Silva, C.N.; Tessmann, D.J.; Barbosa-Tessmann, I.P. Fungi Isolated from Maize (Zea mays L.) Grains and Production of Associated Enzyme Activities. Int. J. Mol. Sci. 2015, 7, 15328–15346. [Google Scholar] [CrossRef]
- Xing, H.Q.; Ma, J.C.; Xu, B.L.; Zhang, S.W.; Wang, J.; Cao, L.; Yang, X.M. Mycobiota of maize seeds revealed by rDNA-ITS sequence analysis of samples with varying storage times. MicrobiologyOpen 2018, 7, e00609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Liu, B.; Jin, J.; Ma, L.; Dai, X.; Pan, L.; Liu, Y.; Zhao, Y.; Xing, F. The Complex Essential Oils Highly Control the Toxigenic Fungal Microbiome and Major Mycotoxins During Storage of Maize. Front. Microbiol. 2019, 10, 1643. [Google Scholar] [CrossRef] [Green Version]
- Gromadzka, K.; Błaszczyk, L.; Chełkowski, J.; Waśkiewicz, A. Occurrence of Mycotoxigenic Fusarium Species and Competitive Fungi on Preharvest Maize Ear Rot in Poland. Toxins 2019, 11, 224. [Google Scholar] [CrossRef] [Green Version]
- Shoresh, M.; Harman, G.E. The molecular basis of shoot responses of maize 517 seedlings to Trichoderma harzianum T22 inoculation of the root: A proteomic approach. Plant Physiol. 2008, 147, 2147–2163. [Google Scholar] [CrossRef] [Green Version]
- Wicklow, D.T.; Roth, S.; Deyrup, S.T.; Gloer, J.B. A protective endophyte of maize: Acremonium zeae antibiotics inhibitory to Aspergillus flavus and Fusarium verticillioides. Mycol. Res. 2005, 109, 610–618. [Google Scholar] [CrossRef] [Green Version]
- Wicklow, D.T.; Poling, S.M. Antimicrobial activity of pyrrocidines from Acremonium zeae against endophytes and pathogens of maize. Phytopathology 2009, 99, 109–115. [Google Scholar] [CrossRef] [Green Version]
- Summerbell, R.C.; Gueidan, C.; Schreors, H.J.; de Hoog, G.S.; Staring, M.; Arocha Rosete, Y.; Guarro, J.; Scott, J.A. Acremonium phylogenetic overview and revision of Gliomastix, Sarocladium, and Trichothecium. Stud. Mycol. 2011, 68, 139–162. [Google Scholar] [CrossRef]
- Ownley, B.H.; Gwinn, K.D.; Vega, F.E. Endophytic fungal entomopathogens with activity against plant pathogens: Ecology and evolution. BioControl 2010, 55, 113–128. [Google Scholar] [CrossRef]
- Dolatabad, H.K.; Javan-Nikkhah, M.; Shier, W.T. Evaluation of antifungal, phosphate solubilisation, and siderophore and chitinase release activities of endophytic fungi from Pistacia vera. Mycol. Prog. 2017, 16, 777–790. [Google Scholar] [CrossRef]
- Nirenberg, H. Untersuchungen über Die Morphologische und Biologische Differenzierung in der Fusarium-Sektion Liseola; Kommissionsverlag Paul Parey: Berlin, Germany, 1976. [Google Scholar]
- Shinde, S.V.; Patel, K.G.; Purohit, M.S.; Pandya, J.R.; Sabalpara, A.N. Lecanicillium lecanii (Zimm) Zare and Gams an important biocontrol agent for the management of insect pests—A Review. Agric. Rev. 2010, 31, 235–252. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehner, S.A.; Samuels, G.J. Molecular systematics of the Hypocreales: A teleomorph gene phylogeny and the status of their anamorphs. Can. J. Bot. 1995, 73, 816–823. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 1, 553–556. [Google Scholar] [CrossRef]
- National Centre for Biotechnology Information. Available online: http://www.ncbi.nlm.nih.gov (accessed on 12 January 2021).
- Jestoi, M.; Rokka, M.; Yli-Mattila, T.; Parikka, P.; Rizzo, A.; Peltonen, K. Presence and concentrations of the Fusarium-related mycotoxins beauvericin, enniatins and moniliformin in finish grain samples. Food Addit. Contam. 2004, 21, 794–802. [Google Scholar] [CrossRef]
- Kanaoka, M.; Isogai, A.; Murakoshi, S.; Ichinoe, M.; Suzuki, A.; Tamura, S. Bassianolide, a new insecticidal cyclodepsipeptide from Beauveria bassiana and Verticillium lecanii. Agric. Biol. Chem. 1978, 42, 629–635. [Google Scholar] [CrossRef]
- Soman, A.G.; Gloer, J.B.; Angawi, R.F.; Wicklow, D.T.; Dowd, P.F. Vertilecanins: New phenopicolinic acid analogues from Verticillium lecanii. J. Nat. Prod. 2001, 64, 189–192. [Google Scholar] [CrossRef]
- He, H.; Yang, H.Y.; Bigelis, R.; Solum, E.H.; Greenstein, M.; Carter, G.T. Pyrrocidines A and B, new antibiotics produced by a filamentous fungus. Tetrahedron Lett. 2002, 43, 1633–1636. [Google Scholar] [CrossRef]
- Reddy, C.S.; Holbert, J.R. The black bundle disease of corn. J. Agric. Res. 1924, 27, 177–206. [Google Scholar]
- Tagne, A.; Neergaard, E.; Hansen, H. Studies of Host—pathogen Interaction between Maize and Acremonium strictum From Cameroon. Eur. J. Plant Pathol. 2002, 108, 93–102. [Google Scholar] [CrossRef]
- Racedo, J.; Salazar, S.M.; Castagnaro, A.P.; Díaz Ricci, J.C. A strawberry disease caused by Acremonium strictum. Eur J. Plant Pathol. 2013, 137, 649–654. [Google Scholar] [CrossRef]
- Potshangbam, M.; Devi, S.I.; Sahoo, D.; Strobel, G.A. Functional Characterization of Endophytic Fungal Community Associated with Oryza sativa L. and Zea mays L. Front. Microbiol. 2017, 8, 325. [Google Scholar] [CrossRef]
- Cobo-Díaz, J.F.; Baroncelli, R.; Le Floch, G.; Picot, A. Combined Metabarcoding and Co-occurrence Network Analysis to Profile the Bacterial, Fungal and Fusarium Communities and Their Interactions in Maize Stalks. Front. Microbiol. 2019, 10, 261. [Google Scholar] [CrossRef] [Green Version]
- Zare, R.; Gams, W. A revision of Verticillium section Prostrata. IV. The genera Lecanicillium and Simplicillium gen. nov. Nova Hedwig. 2001, 73, 1–50. [Google Scholar]
- Kepler, R.M.; Luangsa-Ard, J.J.; Hywel-Jones, N.L.; Quandt, C.A.; Sung, G.H.; Rehner, S.A.; Aime, M.C.; Henkel, T.W.; Sanjuan, T.; Zare, R.; et al. A phylogenetically-based nomenclature for Cordycipitaceae (Hypocreales). IMA Fungus 2017, 8, 335–353. [Google Scholar] [CrossRef]
- Sung, G.-H.; Hywel-Jones, N.L.; Sung, J.-M.; Luangsa-ard, J.J.; Shrestha, B.; Spatafora, J.W. Phylogenetic classification of Cordyceps and the clavicipitaceous fungi. Stud. Mycol. 2007, 57, 5–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goettel, M.S.; Koike, M.; Kim, J.J.; Aiuchi, D.; Shinya, R.; Brodeur, J. Potential of Lecanicillium spp. for management of insects, nematodes and plant diseases. J. Invertebr. Pathol. 2008, 98, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Nicoletti, R.; Becchimanzi, A. Endophytism of Lecanicillium and Akanthomyces. Agriculture 2020, 10, 205. [Google Scholar] [CrossRef]
- Sánchez-Márquez, S.; Bills, G.F.; Zabalgogeazcoa, I. Diversity and structure of the fungal endophytic assemblages from two sympatric coastal grasses. Fungal Divers. 2008, 33, 87–100. [Google Scholar]
- Sánchez-Márquez, M.; Bills, G.F.; Zabalgogeazcoa, I. The endophytic mycobiota of the grass Dactylis Glomerata. Fungal Divers. 2007, 27, 171–195. [Google Scholar]
- Poosakkannu, A.; Nissinen, R.; Kytöviita, M.M. Culturable endophytic microbial communities in the circumpolar grass, Deschampsia flexuosa in a sub-Arctic inland primary succession are habitat and growth stage specific. Environ. Microbiol. Rep. 2015, 7, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Molina-Montenegro, M.A.; Oses, R.; Torres-Díaz, C.; Atala, C.; Núñez, M.A.; Armas, C. Fungal endophytes associated with roots of nurse cushion species have positive efects on native and invasive beneficiary plants in an alpine ecosystem. Perspect. Plant Ecol. Evol. Syst. 2015, 17, 218–226. [Google Scholar] [CrossRef]
- Giordano, L.; Gonthier, P.; Varese, G.C.; Miserere, L.; Nicolotti, G. Mycobiota inhabiting sapwood of healthy and declining Scots pine (Pinus sylvestris L.) trees in the Alps. Fungal Divers. 2009, 38, 69–83. [Google Scholar]
- Behnke-Borowczyk, J.; Kwaśna, H.; Kulawinek, B. Fungi associated with Cyclaneusma needle cast in Scots pine in the west of Poland. For. Pathol. 2019, 49, e12487. [Google Scholar] [CrossRef]
- Gurulingappa, P.; Sword, G.A.; Murdoch, G.; McGee, P.A. Colonization of crop plants by fungal entomopathogens and their efects on two insect pests when in planta. Biol. Control 2010, 55, 34–41. [Google Scholar] [CrossRef]
- de Souza Vieira, P.D.; de Souza Motta, C.M.; Lima, D.; Torres, J.B.; Quecine, M.C.; Azevedo, J.L.; de Oliveira, N.T. Endophytic fungi associated with transgenic and non-transgenic cotton. Mycology 2011, 2, 91–97. [Google Scholar] [CrossRef]
- Dash, C.K.; Bamisile, B.S.; Keppanan, R.; Qasim, M.; Lin, Y.; Islam, S.U.; Hussain, M.; Wang, L. Endophytic entomopathogenic fungi enhance the growth of Phaseolus vulgaris L. (Fabaceae) and negatively affect the development and reproduction of Tetranychus urticae Koch (Acari: Tetranychidae). Microb. Pathog. 2018, 125, 385–392. [Google Scholar] [CrossRef]
- González, V.; Tello, M.L. The endophytic mycota associated with Vitis vinifera in central Spain. Fungal Divers. 2011, 47, 29–42. [Google Scholar] [CrossRef]
- Gromadzka, K.; Górna, K.; Chełkowski, J.; Waśkiewicz, A. Mycotoxins and related Fusarium species in preharvest maize ear rot in Poland. Plant Soil Environ. 2016, 62, 348–354. [Google Scholar] [CrossRef] [Green Version]
- Wicklow, D.T.; Poling, S.M.; Summerbell, R. Occurrence of pyrrocidine and dihydroresorcylide production among Acremonium zeae populations from maize grown in different regions. Can. J. Plant Pathol. 2008, 30, 425–433. [Google Scholar] [CrossRef]
- Poling, S.M.; Wicklow, D.T.; Rogers, K.D.; Gloer, J.B. Acremonium zeae, a protective endophyte of maize, produces dihydro- resorcylide and 7-hydroxydihydroresorcylides. J. Agric. Food Chem. 2008, 56, 3006–3009. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | Isolate Code | Sampling Year | NCBI GenBank Accession No. | |
|---|---|---|---|---|
| ITS | LSU | |||
| Lecanicillium lecanii | 207 | 2015 | MT372981 | MT373083 |
| 213 | MT372982 | MT373084 | ||
| 224 | MT375133 | MT375130 | ||
| 350 | MT372983 | MT373085 | ||
| 345 | MT372984 | MT373086 | ||
| Sarocladium strictum | 228 | 2015 | MT375132 | MT375131 |
| 626 | 2018 | MT372901 | MT374074 | |
| 605_1 | MT372902 | MT374075 | ||
| Sarocladium zeae | 215 | 2015 | MT372974 | MT373076 |
| 217 | MT372976 | MT373078 | ||
| 220 | MT372977 | MT373079 | ||
| 221 | MT372978 | MT373080 | ||
| 226 | MT372979 | MT373081 | ||
| 227A | MT372980 | MT373082 | ||
| 227B | MT372975 | MT373077 | ||
| 601 | 2018 | MT372893 | MT374066 | |
| 603 | MT372894 | MT374067 | ||
| 605 | MT372895 | MT374068 | ||
| 606 | MT372896 | MT374069 | ||
| 613 | MT372893 | MT374066 | ||
| 614 | MT372899 | MT374072 | ||
| 636 | MT372897 | MT374070 | ||
| 651 | MT372898 | MT374071 | ||
| 652 | MT372899 | MT374072 | ||
| 658 | MT372900 | MT374073 | ||
| Compound | 10-hydoksy-8-decenoic acid | 2-decenedioic acid | Verttilecanin A | Verttilecanin A methy ester | Verttilecanin C | Pyrrocidine A | Pyrrocidine B | Enns B1 | BEA | Bassianolide | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | Isolate Code | ||||||||||
| L. lecanii | 207 | -1 | +2 | - | + | + | - | + | - | - | + |
| 213 | - | + | - | + | - | - | + | - | - | + | |
| 224 | - | - | - | + | - | - | + | - | - | + | |
| 350 | - | - | - | + | - | - | - | - | - | + | |
| 345 | - | - | - | + | - | - | - | - | - | + | |
| S. strictum | 228 | + | + | + | + | - | + | + | + | - | + |
| 626 | + | + | + | + | - | - | + | + | - | + | |
| 605_1 | + | + | + | + | - | + | - | + | - | + | |
| S. zeae | 215 | + | - | + | + | + | - | - | - | - | + |
| 217 | + | + | + | + | + | - | - | - | - | + | |
| 220 | - | - | + | + | - | - | + | - | - | + | |
| 221 | + | + | + | + | - | - | - | - | - | + | |
| 226 | + | + | + | + | - | + | + | - | + | + | |
| 227A | + | + | + | + | - | + | + | - | + | + | |
| 227B | + | - | + | + | - | + | + | - | + | + | |
| 601 | - | - | + | + | - | + | + | - | - | + | |
| 603 | + | - | + | - | - | - | + | - | - | + | |
| 605 | - | + | + | + | - | - | + | - | - | + | |
| 606 | + | - | + | - | - | - | + | - | - | + | |
| 613 | - | - | + | + | - | - | + | - | - | + | |
| 614 | - | - | + | - | - | + | + | - | - | + | |
| 636 | - | + | + | + | - | - | + | - | - | + | |
| 651 | - | - | + | - | - | - | - | - | - | + | |
| 652 | - | + | + | - | - | - | + | - | - | + | |
| 658 | - | - | + | - | - | - | - | - | - | + | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Błaszczyk, L.; Waśkiewicz, A.; Gromadzka, K.; Mikołajczak, K.; Chełkowski, J. Sarocladium and Lecanicillium Associated with Maize Seeds and Their Potential to Form Selected Secondary Metabolites. Biomolecules 2021, 11, 98. https://doi.org/10.3390/biom11010098
Błaszczyk L, Waśkiewicz A, Gromadzka K, Mikołajczak K, Chełkowski J. Sarocladium and Lecanicillium Associated with Maize Seeds and Their Potential to Form Selected Secondary Metabolites. Biomolecules. 2021; 11(1):98. https://doi.org/10.3390/biom11010098
Chicago/Turabian StyleBłaszczyk, Lidia, Agnieszka Waśkiewicz, Karolina Gromadzka, Katarzyna Mikołajczak, and Jerzy Chełkowski. 2021. "Sarocladium and Lecanicillium Associated with Maize Seeds and Their Potential to Form Selected Secondary Metabolites" Biomolecules 11, no. 1: 98. https://doi.org/10.3390/biom11010098