Naphthoquinone-Based Meroterpenoids from Marine-Derived Streptomyces sp. B9173

Abstract
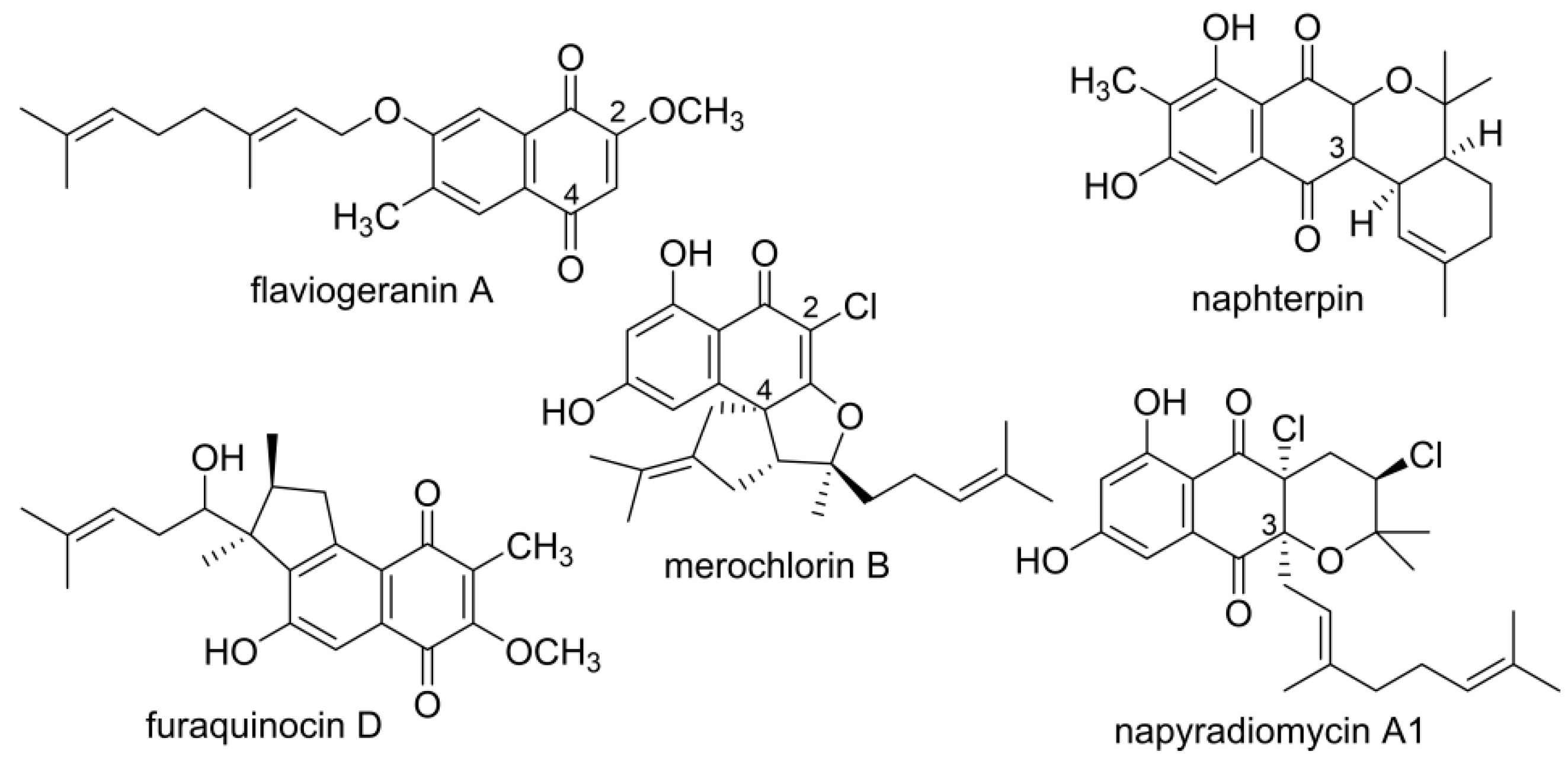
:1. Introduction
2. Materials and Methods
2.1. General Experimental Procedures
2.2. Bacterial Strains, Culture Conditions, and Biochemicals
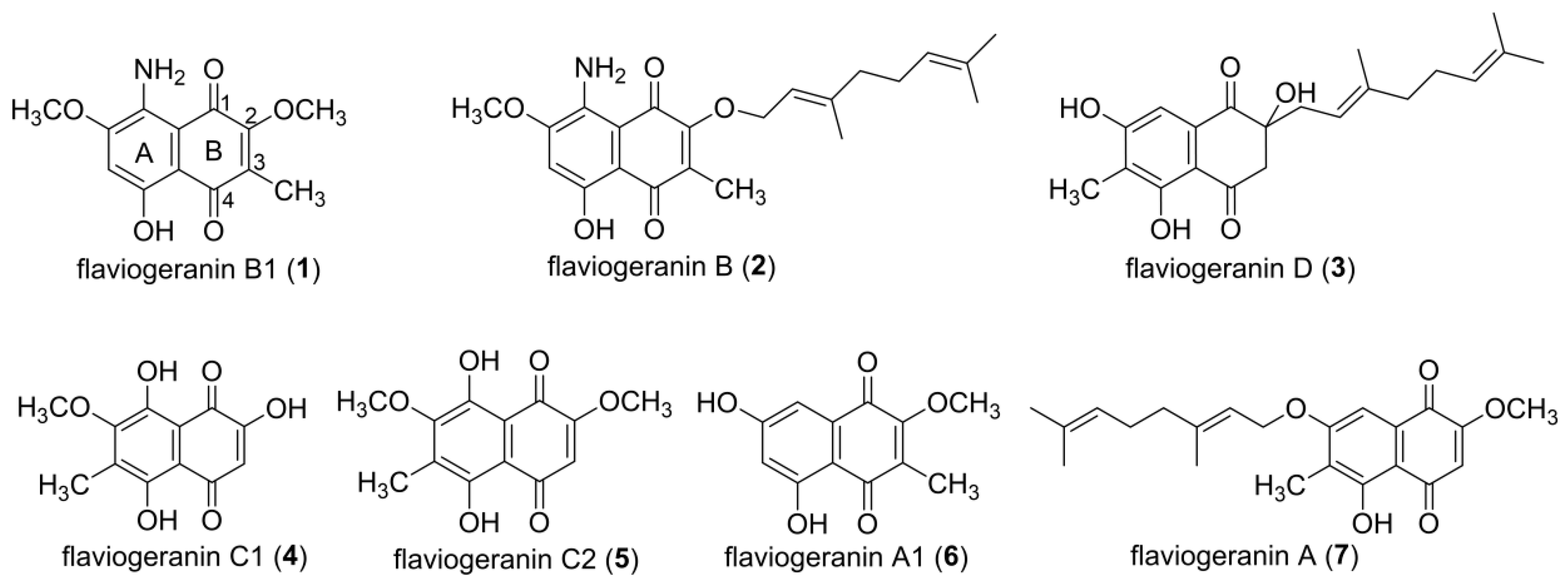
2.3. Extraction and Isolation of Flaviogeranin Congeners
2.4. Bioactivity Assay
3. Results
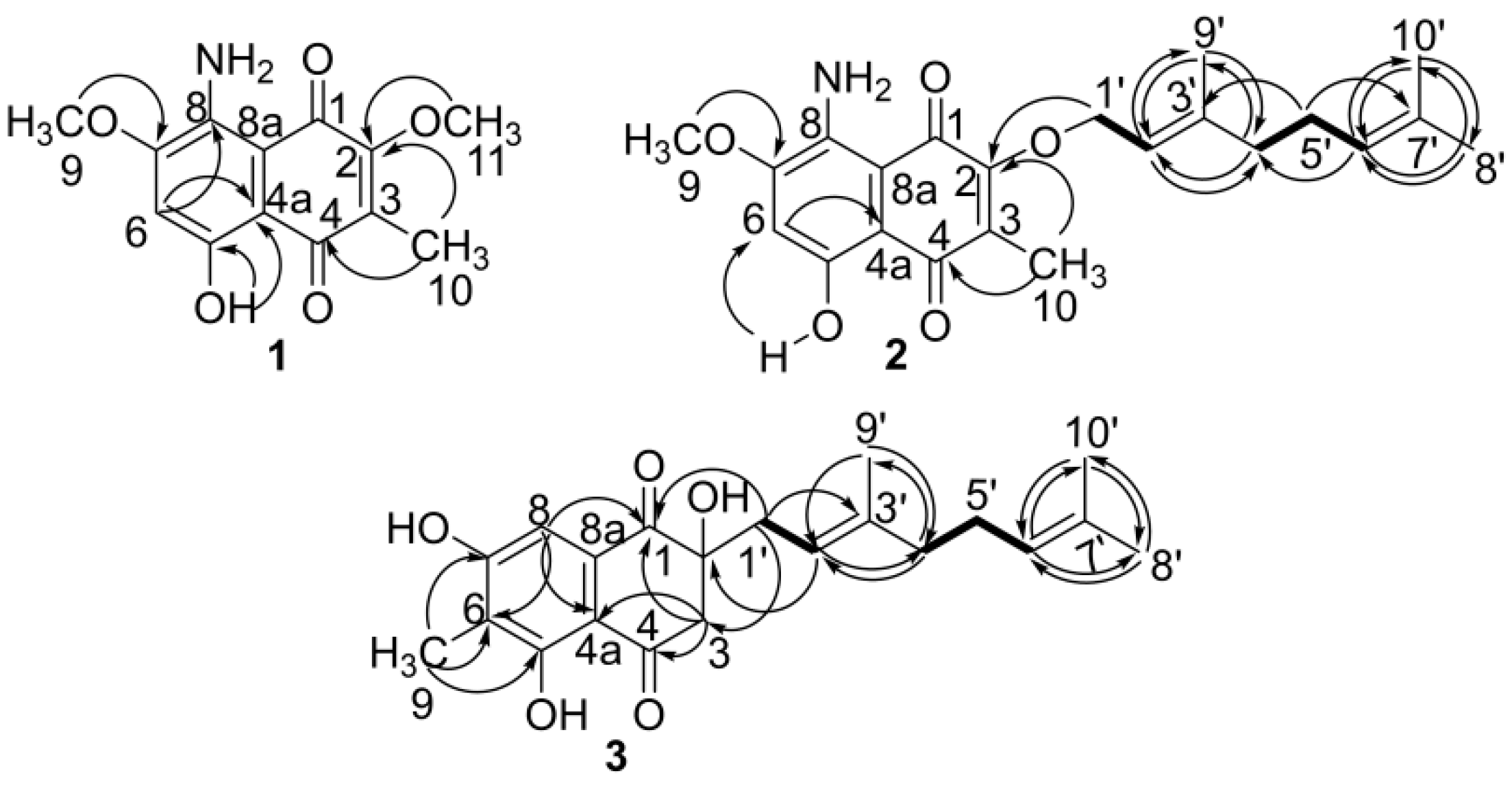
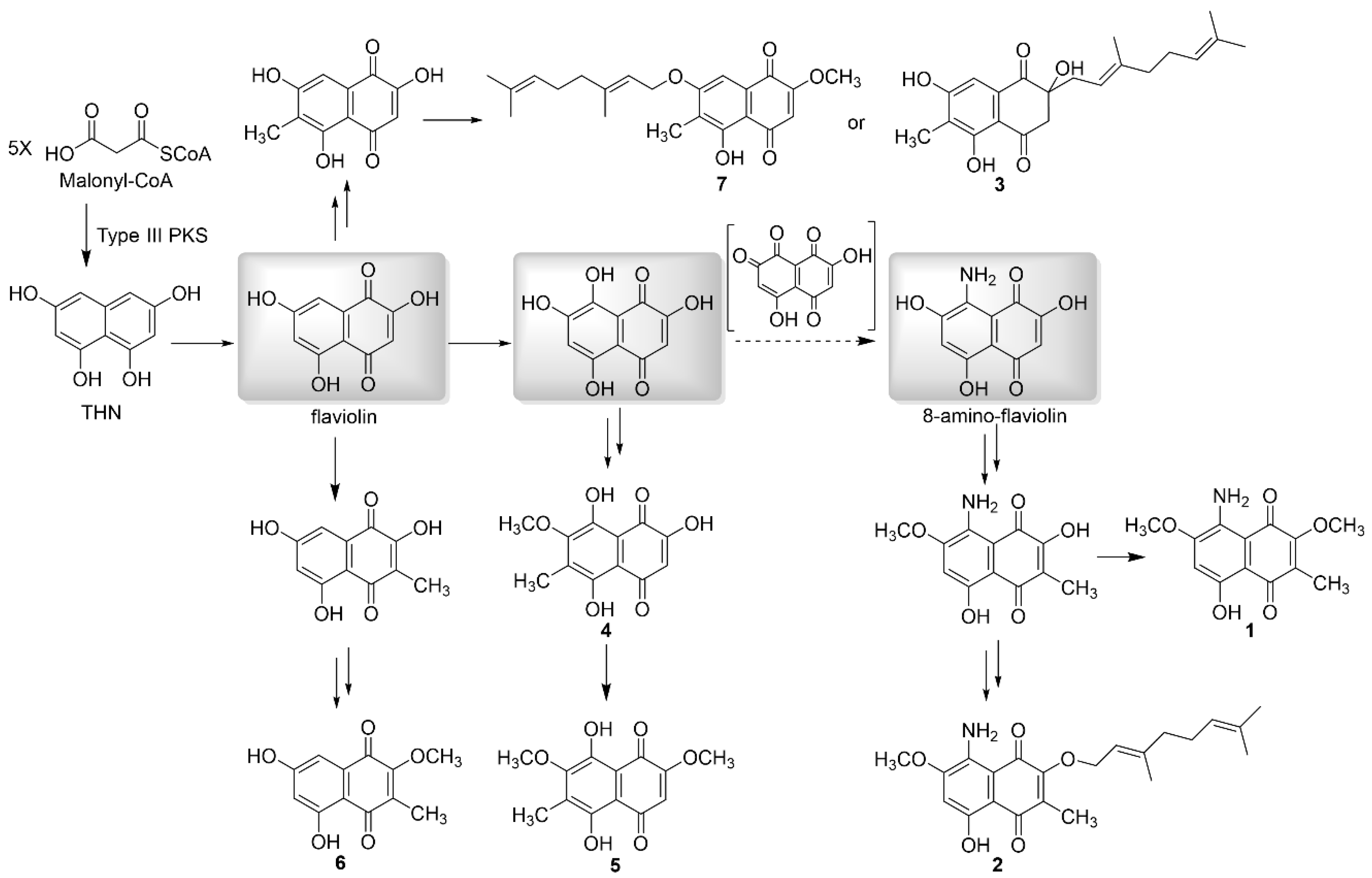
3.1. Structural Elucidation of Naphthoquinone Family Metabolites
3.2. Evaluation of Bioactivities
3.3. Isolated New Natural Products Revealing New Insight into Flaviogeranin Biosynthesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kobayashi, J. Search for new bioactive marine natural products and application to drug development. Chem. Pharm. Bull. (Tokyo) 2016, 64, 1079–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuart, K.A.; Welsh, K.; Walker, M.C.; Edrada-Ebel, R. Metabolomic tools used in marine natural product drug discovery. Exp. Opin. Drug Discov. 2020, 15, 499–522. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Chung-Davidson, Y.W.; Bussy, U.; Li, W. Recent advances and applications of experimental technologies in marine natural product research. Mar. Drugs 2015, 13, 2694–2713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2020, 37, 175–223. [Google Scholar] [CrossRef]
- Schorn, M.A.; Alanjary, M.M.; Aguinaldo, K.; Korobeynikov, A.; Podell, S.; Patin, N.; Lincecum, T.; Jensen, P.R.; Ziemert, N.; Moore, B.S. Sequencing rare marine actinomycete genomes reveals high density of unique natural product biosynthetic gene clusters. Microbiology 2016, 162, 2075–2086. [Google Scholar] [CrossRef]
- Bode, H.B.; Bethe, B.; Hofs, R.; Zeeck, A. Big effects from small changes: Possible ways to explore nature’s chemical diversity. ChemBioChem 2002, 3, 619–627. [Google Scholar] [CrossRef]
- Romano, S.; Jackson, S.A.; Patry, S.; Dobson, A.D.W. Extending the “One Strain Many Compounds” (OSMAC) Principle to Marine Microorganisms. Mar. Drugs 2018, 16, 244. [Google Scholar] [CrossRef] [Green Version]
- Sunassee, S.N.; Davies-Coleman, M.T. Cytotoxic and antioxidant marine prenylated quinones and hydroquinones. Nat. Prod. Rep. 2012, 29, 513–535. [Google Scholar] [CrossRef]
- Shin-ya, K.; Imai, S.; Furihata, K.; Hayakawa, Y.; Kato, Y.; Vanduyne, G.D.; Clardy, J.; Seto, H. Isolation and structural elucidation of an antioxidative agent, naphterpin. J. Antibiot (Tokyo) 1990, 43, 444–447. [Google Scholar] [CrossRef] [Green Version]
- Hayakawa, Y.; Yamazaki, Y.; Kurita, M.; Kawasaki, T.; Takagi, M.; Shin-Ya, K. Flaviogeranin, a new neuroprotective compound from Streptomyces sp. J. Antibiot (Tokyo) 2010, 63, 379–380. [Google Scholar] [CrossRef] [Green Version]
- Ishibashi, M.; Funayama, S.; Anraku, Y.; Komiyama, K.; Omura, S. Novel antibiotics, furaquinocins C, D, E, F, G and H. J. Antibiot (Tokyo) 1991, 44, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Funa, N.; Ohnishi, Y.; Fujii, I.; Shibuya, M.; Ebizuka, Y.; Horinouchi, S. A new pathway for polyketide synthesis in microorganisms. Nature 1999, 400, 897–899. [Google Scholar] [CrossRef] [PubMed]
- Izumikawa, M.; Shipley, P.R.; Hopke, J.N.; O’Hare, T.; Xiang, L.; Noel, J.P.; Moore, B.S. Expression and characterization of the type III polyketide synthase 1,3,6,8-tetrahydroxynaphthalene synthase from Streptomyces coelicolor A3(2). J. Ind. Microbiol. Biotechnol. 2003, 30, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Austin, M.B.; Izumikawa, M.; Bowman, M.E.; Udwary, D.W.; Ferrer, J.L.; Moore, B.S.; Noel, J.P. Crystal structure of a bacterial type III polyketide synthase and enzymatic control of reactive polyketide intermediates. J. Biol. Chem. 2004, 279, 45162–45174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawasaki, T.; Hayashi, Y.; Kuzuyama, T.; Furihata, K.; Itoh, N.; Seto, H.; Dairi, T. Biosynthesis of a natural polyketide-isoprenoid hybrid compound, furaquinocin A: Identification and heterologous expression of the gene cluster. J. Bacteriol 2006, 188, 1236–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaysser, L.; Bernhardt, P.; Nam, S.J.; Loesgen, S.; Ruby, J.G.; Skewes-Cox, P.; Jensen, P.R.; Fenical, W.; Moore, B.S. Merochlorins A-D, cyclic meroterpenoid antibiotics biosynthesized in divergent pathways with vanadium-dependent chloroperoxidases. J. Am. Chem. Soc. 2012, 134, 11988–11991. [Google Scholar] [CrossRef] [Green Version]
- Kuzuyama, T.; Noel, J.P.; Richard, S.B. Structural basis for the promiscuous biosynthetic prenylation of aromatic natural products. Nature 2005, 435, 983–987. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.Y.; Liu, Y.Y.; Huang, T.; Zou, Y.; Huang, T.T.; Hu, K.F.; Deng, Z.X.; Lin, S.J. Divergent biosynthesis of indole alkaloids FR900452 and spiro-maremycins. Org. Biomol. Chem. 2018, 16, 5446–5451. [Google Scholar] [CrossRef]
- Lan, Y.X.; Zou, Y.; Huang, T.T.; Wang, X.Z.; Brock, N.L.; Deng, Z.X.; Lin, S.J. Indole methylation protects diketopiperazine configuration in the maremycin biosynthetic pathway. Sci. China-Chem. 2016, 59, 1224–1228. [Google Scholar] [CrossRef]
- Balkbindseil, W.; Helmke, E.; Weyland, H.; Laatsch, H. Marine Bacteria.8. Maremycin-a and maremycin-B, new diketopiperazines from a marine Streptomyces sp. Liebigs Ann. 1995, 7, 1291–1294. [Google Scholar] [CrossRef]
- Tang, Y.Q.; Sattler, I.; Thiericke, R.; Grabley, S.; Feng, X.Z. Maremycins C and D, new diketopiperazines, and maremycins E and F, novel polycyclic spiro-indole metabolites isolated from Streptomyces sp. Eur. J. Org. Chem. 2001, 2, 261–267. [Google Scholar] [CrossRef]
- Luo, F.; Zou, Y.; Huang, T.; Lin, S. Draft Genome Sequence of Streptomyces sp. B9173, a producer of indole diketopiperazine maremycins. Genome Announc. 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Isogai, S.; Nishiyama, M.; Kuzuyama, T. Identification of 8-amino-2,5,7-trihydroxynaphthalene-1,4-dione, a novel intermediate in the biosynthesis of Streptomyces meroterpenoids. Bioorg Med. Chem. Lett. 2012, 22, 5823–5826. [Google Scholar] [CrossRef] [PubMed]
- Funa, N.; Funabashi, M.; Yoshimura, E.; Horinouchi, S. A novel quinone-forming monooxygenase family involved in modification of aromatic polyketides. J. Biol. Chem. 2005, 280, 14514–14523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumano, T.; Tomita, T.; Nishiyama, M.; Kuzuyama, T. Functional characterization of the promiscuous prenyltransferase responsible for furaquinocin biosynthesis: Identification of a physiological polyketide substrate and its prenylated reaction products. J. Biol. Chem. 2010, 285, 39663–39671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glazunov, V.P.; Tchizhova, A.Y.; Shestak, O.P.; Sopel’nyak, G.I.; Anufriev, V.P. Chemistry of naphthazarin derivatives-8. Determination of structures of substituted 2-hydroxy-6(7)-methoxynaphthazarins and 7(8)-hydroxypyranonaphthazarins by IR spectroscopy. Russ. Chem. Bull. 2001, 50, 95–100. [Google Scholar] [CrossRef]
- Zhou, H.; Li, Y.; Tang, Y. Cyclization of aromatic polyketides from bacteria and fungi. Nat. Prod. Rep. 2010, 27, 839–868. [Google Scholar] [CrossRef] [Green Version]
- Gogineni, V.; Hamann, M.T. Marine natural product peptides with therapeutic potential: Chemistry, biosynthesis, and pharmacology. Biochim. Biophys. Acta-Gen. Subj. 2018, 1862, 81–196. [Google Scholar] [CrossRef]
- Dairi, T.; Kuzuyama, T.; Nishiyama, M.; Fujii, I. Convergent strategies in biosynthesis. Nat. Prod. Rep. 2011, 28, 1054–1086. [Google Scholar] [CrossRef]
- Hiratsuka, T.; Furihata, K.; Ishikawa, J.; Yamashita, H.; Itoh, N.; Seto, H.; Dairi, T. An alternative menaquinone biosynthetic pathway operating in microorganisms. Science 2008, 321, 1670–1673. [Google Scholar] [CrossRef]
- Cane, D.E.; Ikeda, H. Exploration and mining of the bacterial terpenome. Acc. Chem. Res. 2012, 45, 463–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | δH(Multi, J/Hz) | δC(DEPT) | 1H-1H COSY | gHMBC |
|---|---|---|---|---|
| 1 | 180.1 (s) | |||
| 2 | 159.4 (s) | |||
| 3 | 131.6 (s) | |||
| 4 | 186.8 (s) | |||
| 4a | 106.1 (s) | |||
| 5 | 160.3 (s) | |||
| 6 | 6.48 (1H,s) | 104.3 (d) | 5,7,4a,8 | |
| 7 | 155.4 (s) | |||
| 8 | 139.9 (s) | |||
| 8a | 107.5 (s) | |||
| 9 | 3.97 (3H,s) | 56.4 (q) | 7 | |
| 10 | 2.09 (3H,s) | 8.8 (q) | 6,2 | |
| 11 | 4.07 (3H,s) | 61.0 (q) | 2 | |
| 5-OH | 14.41 (brs) | 4,4a |
| No | δH(Multi, J/Hz) | δC(DEPT) | 1H-1H COSY | gHMBC |
|---|---|---|---|---|
| 1 | 180.4 (s) | |||
| 2 | 158.7 (s) | |||
| 3 | 132.5 (s) | |||
| 4 | 186.8 (s) | |||
| 4a | 106.1 (s) | |||
| 5 | 160.1 (s) | |||
| 6 | 6.47 (1H,s) | 104.3 (d) | 5,7,4a | |
| 7 | 155.4 (s) | |||
| 8 | 139.7 (s) | |||
| 8a | 107.5 (s) | |||
| 9 | 3.96 (3H,s) | 56.4 (q) | 7 | |
| 10 | 2.09 (3H,s) | 9.1 (q) | 2,4 | |
| 1′ | 4.87 (2H,d,7.2) | 69.8 (t) | 2′ | 2,3′ |
| 2′ | 5.49 (1H,m) | 119.8 (d) | 1′ | 9′,4′ |
| 3′ | 142.6 (s) | |||
| 4′ | 2.06 (2H,m) | 39.6 (t) | 5′ | 3′,2′,9′ |
| 5′ | 2.07 (2H,m) | 26.3 (t) | 4′,6′ | 3′,7′ |
| 6′ | 5.05 (1H,m) | 123.7 (d) | 5′ | 8′,4′,10′ |
| 7′ | 131.8 (s) | |||
| 8′ | 1.65 (3H,s) | 25.6 (q) | 7′,10′,6′ | |
| 9′ | 1.70 (3H,s) | 16.5 (q) | 4′,2′,3′ | |
| 10′ | 1.58 (3H,s) | 17.6 (q) | 7′,8′,6′ | |
| 5-OH | 14.42 (brs) | 4a,5 |
| No | δH(Multi, J/Hz) | δC(DEPT) | 1H-1H COSY | gHMBC |
|---|---|---|---|---|
| 1 | 199.7 (s) | |||
| 2 | 77.9 (s) | |||
| 3 | 3.23 (1H,d,16.8) 3.13 (1H,d,16.8) | 49.1 (t) | 1,4a | |
| 4 | 198.9 (s) | |||
| 4a | 112.0 (s) | |||
| 5 | 161.2 (s) | |||
| 6 | 119.2 (s) | |||
| 7 | 162.1 (s) | |||
| 8 | 7.07 (1H,s) | 106.5 (d) | 4a,6 | |
| 8a | 131.4 (s) | |||
| 9 | 2.19 (3H,s) | 8.1 (q) | 5,7 | |
| 1′ | 2.49 (1H,dd,14.5,8.3) 2.38 (1H,dd,14.5,8.3) | 38.9 (t) | 2′ | 2,3′,1 |
| 2′ | 4.98 (1H,t,6.9) | 116.0 (d) | 1′ | 9′,4′ |
| 3′ | 141.0 (s) | |||
| 4′ | 1.94 (2H,m) | 39.8 (t) | 5′ | 3′,2′,9′ |
| 5′ | 2.02 (2H,m) | 26.3 (t) | 4′,6′ | 3′,7′ |
| 6′ | 5.03 (1H,t,7.5) | 123.9 (d) | 5′ | 8′,4′,10′ |
| 7′ | 131.3 (s) | |||
| 8′ | 1.67 (3H,s) | 25.6 (q) | 7′,10′,6′ | |
| 9′ | 1.43 (3H,s) | 16.1 (q) | 4′,2′,3′ | |
| 10′ | 1.57 (3H,s) | 17.6 (q) | 7′,8′,6′ | |
| 4-OH | 12.55 (brs) | 3,4a,5 |
| Compound | MIC (μg/mL) | IC50 (μM) | ||
|---|---|---|---|---|
| S. aureus | M. smegmatis | A549 | Hela | |
| 1 | 14.6 ± 0.3 | 12.4 ± 0.3 | 25.7 ± 0.4 | 34.7 ± 0.6 |
| 2 | 28.1 ± 0.4 | 35.1 ± 0.6 | 46.6 ± 0.4 | 50.2 ± 0.5 |
| 3 | 9.2 ± 0.2 | 5.2 ± 0.2 | 0.6 ± 0.2 | 0.4 ± 0.2 |
| 5 | 8.1 ± 0.2 | 7.7 ± 0.2 | 0.9 ± 0.2 | 1.1 ± 0.3 |
| erythromycin | 7.6 ± 0.2 | 4.5 ± 0.2 | NA | NA |
| doxorubicin | NA | NA | 0.1 ± 0.0 | 0.5 ± 0.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, X.; Wang, X.; Huang, T.; Deng, Z.; Lin, S. Naphthoquinone-Based Meroterpenoids from Marine-Derived Streptomyces sp. B9173. Biomolecules 2020, 10, 1187. https://doi.org/10.3390/biom10081187
Shen X, Wang X, Huang T, Deng Z, Lin S. Naphthoquinone-Based Meroterpenoids from Marine-Derived Streptomyces sp. B9173. Biomolecules. 2020; 10(8):1187. https://doi.org/10.3390/biom10081187
Chicago/Turabian StyleShen, Xinqian, Xiaozheng Wang, Tingting Huang, Zixin Deng, and Shuangjun Lin. 2020. "Naphthoquinone-Based Meroterpenoids from Marine-Derived Streptomyces sp. B9173" Biomolecules 10, no. 8: 1187. https://doi.org/10.3390/biom10081187