The Role of Fe, Zn, and Cu in Pregnancy


Abstract
:1. Introduction
2. Iron (Fe)
3. Copper (Cu)
4. Zinc (Zn)
5. Fe vs. Cu
6. Fe vs. Zn
7. Cu vs. Zn
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Choi, R.; Sun, J.; Yoo, H.; Kim, S.; Cho, Y.Y.; Kim, H.J.; Kim, W.K.; Chung, J.H.; Oh, S.; Lee, S.-Y. A Prospective study of serum trace elements in healthy korean pregnant women. Nutrients 2016, 8, 749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kot, K.; Kosik-Bogacka, D.; Łanocha-Arendarczyk, N.; Malinowski, W.; Szymański, S.; Mularczyk, M.; Tomska, N.; Rotter, I. Interactions between 14 Elements in the Human Placenta, Fetal Membrane and Umbilical Cord. Int. J. Environ. Res. Public Health 2019, 16, 1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocyłowski, R.; Lewicka, I.; Grzesiak, M.; Gaj, Z.; Oszukowski, P.; von Kaisenberg, C.; Suliburska, J. Evaluation of Mineral Concentrations in Maternal Serum Before and After Birth and in Newborn Cord Blood Postpartum-Preliminary Study. Biol. Trace Elem. Res. 2018, 182, 217–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasowicz, W.; Wolkanin, P.; Bednarski, M.; Gromadzinska, J.; Sklodowska, M.; Grzybowska, K. Plasma trace element (Se, Zn, Cu) concentrations in maternal and umbilical cord blood in Poland. Biol. Trace Elem. Res. 1993, 38, 205–215. [Google Scholar] [CrossRef]
- Kovalchuk, L.; Tarkhanova, A.; Tarkhanov, A. The essential and toxic effects of trace elements in the biological tissues of pregnant women and newborn babies (an ecologically unfavourable region). Int. J. Saf. Secur. Eng. 2013, 3, 265–277. [Google Scholar] [CrossRef]
- Skalnaya, M.G.; Tinkov, A.A.; Lobanova, Y.N.; Chang, J.S.; Skalny, A.V. Serum levels of copper, iron, and manganese in women with pregnancy, miscarriage, and primary infertility. J. Trace Elem. Med. Biol. 2019, 56, 124–130. [Google Scholar] [CrossRef]
- Sabra, S.; Malmqvist, E.; Saborit, A.; Gratacós, E.; Gomez Roig, M.D. Heavy metals exposure levels and their correlation with different clinical forms of fetal growth restriction. PLoS ONE 2017, 12, e0185645. [Google Scholar] [CrossRef]
- Mbofung, C.M.F.; Subbarau, V.V. Trace element (Zinc, copper, iron and magnesium) concentrations in human placenta and their relationship to birth weight of babies. Nutr. Res. 1990, 10, 359–366. [Google Scholar] [CrossRef]
- Punshon, T.; Li, Z.; Jackson, B.P.; Parks, W.T.; Romano, M.; Conway, D.; Baker, E.R.; Karagas, M.R. Placental metal concentrations in relation to placental growth, efficiency and birth weight. Environ. Int. 2019, 126, 533–542. [Google Scholar] [CrossRef]
- Young, S.M.; Gryder, L.K.; David, W.B.; Teng, Y.; Gerstenberger, S.; Benyshek, D.C. Human placenta processed for encapsulation contains modest concentrations of 14 trace minerals and elements. Nutr. Res. 2016, 36, 872–878. [Google Scholar] [CrossRef]
- de Angelis, P.; Miller, R.K.; Darrah, T.H.; Katzman, P.J.; Pressman, E.K.; Kent, T.R.; O’Brien, K.O. Elemental content of the placenta: A comparison between two high-risk obstetrical populations, adult women carrying multiples and adolescents carrying singletons. Environ. Res. 2017, 158, 553–565. [Google Scholar] [CrossRef] [PubMed]
- Mikelson, C.K.; Troisi, J.; LaLonde, A.; Symes, S.J.K.; Thurston, S.W.; DiRe, L.M.; Adair, C.D.; Miller, R.K.; Richards, S.M. Placental concentrations of essential, toxic, and understudied metals and relationships with birth outcomes in Chattanooga, T.N. Environ. Res. 2019, 168, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Rathore, S.; Gupta, A.; Batra, H.S.; Rathore, R. Comparative study of trace elements and serum ceruloplasmin level in normal and pre-eclamptic pregnancies with their cord blood. Biomed. Res. 2010, 22, 209–212. [Google Scholar]
- Singh, L.; Agarwal, P.; Anand, M.; Taneja, A. Toxic and essential metals in placenta and its relation with lipid peroxides/glutathione status in pre-term and full-term deliveries. Asian J. Med. Sci. 2015, 7, 34–39. [Google Scholar] [CrossRef] [Green Version]
- Upadhyaya, C.; Mishra, S.; Ajmera, P.; Sharma, P. Serum iron, copper and zinc status in maternal and cord blood. Indian J. Clin Biochem. 2004, 19, 48–52. [Google Scholar] [CrossRef] [Green Version]
- Thaker, R.; Oza, H.; Shaikh, I.; Kumar, S. Correlation of Copper and Zinc in Spontaneous Abortion. Int. J. Fertil. Steril. 2019, 13, 97–101. [Google Scholar] [CrossRef]
- Reddy, Y.S.; Aparna, Y.; Ramalaksmi, B.A.; Kumar, B.D. Lead and trace element levels in placenta, maternal and cord blood: A cross-sectional pilot study. J. Obstet. Gynaecol. Res. 2014, 40, 2184–2190. [Google Scholar] [CrossRef]
- Irwinda, R.; Wibowo, N.; Putri, A.S. The Concentration of Micronutrients and Heavy Metals in Maternal Serum, Placenta, and Cord Blood: A Cross-Sectional Study in Preterm Birth. J. Pregnancy 2019, 2019, 5062365. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, M.; Yasutake, A.; Domingo, J.L.; Chan, H.M.; Kubota, M.; Murata, K. Relationships between trace element concentrations in chorionic tissue of placenta and umbilical cord tissue: Potential use as indicators for prenatal exposure. Environ. Int. 2013, 60, 106–111. [Google Scholar] [CrossRef] [Green Version]
- Shen, P.J.; Gong, B.; Xu, F.Y.; Luo, Y. Four trace elements in pregnant women and their relationships with adverse pregnancy outcomes. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 4690–4697. [Google Scholar]
- Awadallah, S.M.; Abu-Elteen, K.H.; Elkarmi, A.Z.; Qaraein, S.H.; Salem, N.M.; Mubarak, M.S. Maternal and cord blood serum levels of zinc, copper, and iron in healthy pregnant Jordanian women. J. Trace Elem. Exp. Med. 2004, 17, 1–8. [Google Scholar] [CrossRef]
- King, J.C. Physiology of pregnancy and nutrient metabolism. Am. J. Clin. Nutr. 2000, 71, 1218–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caan, B.; Horgen, D.M.; Margen, S.; King, J.C.; Jewell, N.P. Benefits associated with WIC supplemental feeding during the interpregnancy interval. Am. J. Clin. Nutr. 1987, 45, 29–41. [Google Scholar] [CrossRef] [PubMed]
- McAlpine, J.M.; McKeating, D.R.; Vincze, L.; Vanderlelie, J.J.; Perkins, A.V. essential mineral intake during pregnancy and its association with maternal health and birth outcomes in South East Queensland, Australia. Nutr. Metab. Insights 2019, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Szymankiewicz, M. Incidence and mortality of newborns from multiple pregnancies. Perinatol. Neonatol. i Ginekol. 2010, 3, 245–248. [Google Scholar]
- Filipecka-Tyczka, D. Medical care for a pregnant woman with a multiple pregnancy. Postępy Nauk Med. 2016, 7, 475–482. [Google Scholar]
- Gertig, H.; Przysławski, J. BBromatology: An Outline of Food and Nutrition Science; Wydawnictwo Lekarskie PZWL: Warsaw, Poland, 2007. [Google Scholar]
- Camaschella, C. Iron-deficiency anemia. N. Engl. J. Med. 2015, 372, 1832–1843. [Google Scholar] [CrossRef] [Green Version]
- Artym, J.; Zimecki, M. The role of lactoferrin in the proper development of newborns. Postępy Hig. Med. Dosw. 2005, 59, 421–432. [Google Scholar]
- Terada, N.; Or, R.; Szepesi, A.; Lucas, J.J.; Gelfand, E.W. Definition of the roles for iron and essential fatty acids in cell cycle progression of normal human T lymphocytes. Exp. Cell Res. 1993, 204, 260–267. [Google Scholar] [CrossRef]
- Zhang, C. Essential functions of iron-requiring proteins in DNA replication, repair and cell cycle control. Protein Cell. 2014, 5, 750–760. [Google Scholar] [CrossRef] [Green Version]
- Beard, J.L. Iron biology in immune function, muscle metabolism and neuronal functioning. J. Nutr. 2001, 131, 568S–580S. [Google Scholar] [CrossRef] [PubMed]
- Dejkhamron, P.; Wejaphikul, K.; Mahatumarat, T.; Silvilairat, S.; Charoenkwan, P.; Saekho, S.; Unachak, K. Vitamin D deficiency and its relationship with cardiac iron and function in patients with transfusion-dependent thalassemia at Chiang Mai University Hospital. Pediatr. Hematol. Oncol. 2018, 35, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Tavsan, Z.; Ayar Kayali, H. The Effect of Iron and Copper as an Essential Nutrient on Mitochondrial Electron Transport System and Lipid Peroxidation in Trichoderma harzianum. Appl Biochem. Biotechnol. 2013, 170, 1665–1675. [Google Scholar] [CrossRef]
- Aisen, P.; Enns, C.; Wessling-Resnick, M. Chemistry and biology of eukaryotic iron metabolism. Int. J. Biochem. Cell Biol. 2001, 33, 940–959. [Google Scholar] [CrossRef]
- Lieu, P.T.; Heiskala, M.; Peterson, P.A.; Yang, Y. The roles of iron in health and disease. Mol. Asp. Med. 2001, 22, 1–87. [Google Scholar] [CrossRef]
- Abbaspour, N.; Hurrell, R.; Kelishadi, R. Review on iron and its importance for human health. J. Res. Med. Sci. 2014, 19, 164–174. [Google Scholar]
- Bothwell, T.H.; Charlton, R.W. A general approach to the problems of iron deficiency and iron overload in the population at large. Semin Hematol. 1982, 19, 54–67. [Google Scholar]
- DeMaeyer, E.; Dallman, P.; Gurney, J.; Hallberg, L.; Sood, S.K.; Srikantia, S.G. Preventing and Controlling Iron Deficiency Anaemia through Primary Health Care: A Guide for Health Administrators and Programme Managers; World Health Organization, Ed.; WHO: Geneva, Switzerland, 1989; 58p. [Google Scholar]
- Scholl, T.O.; Reilly, T. Anemia, iron and pregnancy outcome. J. Nutr. 2000, 130, 443–447. [Google Scholar] [CrossRef] [Green Version]
- Hytten, F. Blood volume changes in normal pregnancy. Clin. Haematol. 1985, 14, 601–612. [Google Scholar] [CrossRef]
- Khoigani, M.G.; Goli, S.; Hasanzadeh, A. The relationship of hemoglobin and hematocrit in the first and second half of pregnancy with pregnancy outcome. Iran. J. Nurs Midwifery Res. 2012, 17 (Suppl. 1), S165–S170. [Google Scholar]
- Soma-Pillay, P.; Nelson-Piercy, C.; Tolppanen, H.; Mebazaa, A.; Tolppanen, H.; Mebazaa, A. Physiological changes in pregnancy. Cardiovasc J. Afr. 2016, 27, 89–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laflamme, E.M. Maternal hemoglobin concentration and pregnancy outcome: A study of the effects of elevation in el alto, bolivia. McGill J. Med. 2011, 13, 47. [Google Scholar] [PubMed]
- Rogne, T.; Tielemans, M.J.; Chong, M.F.-F.; Yajnik, C.S.; Krishnaveni, G.V.; Poston, L.; Jaddoe, V.W.V.; Steegers, E.A.P.; Joshi, S.; Chong, Y.-S.; et al. Associations of maternal vitamin B12 concentration in pregnancy with the risks of preterm birth and low birth weight: A systematic review and meta-analysis of individual participant data. Am. J. Epidemiol. 2017, 185, 212–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciobanu, A.M.; Colibaba, S.; Cimpoca, B.; Peltecu, G.; Panaitescu, A.M. Thrombocytopenia in pregnancy. Maedica 2016, 11, 55–60. [Google Scholar]
- Kopeć, I. Hematologic complications during pregnancy. Hematologia 2016, 7, 295–302. [Google Scholar] [CrossRef] [Green Version]
- WHO/UNICEF/UNO IDA. Prevention, Assessment and Control. Report of a WHO/UNICEF/UNO Consultation; WHO: Geneva, Switzerland, 1998. [Google Scholar]
- Centers for Disease Control (CDC). CDC criteria for anemia in children and childbearing-aged women. Morb. Mortal. Wkly. Rep. 1989, 38, 400–404. [Google Scholar]
- Paweł, L.; Rafał, R.; Starzyński, A.S.; Robert Staroń, A.G. Dietary iron deficiency anemia. Kosmos 2014, 63, 373–379. [Google Scholar]
- Anderson, G.J.; Frazer, D.M.; McKie, A.T.; Vulpe, C.D.; Smith, A. Mechanisms of haem and non-haem iron absorption: Lessons from inherited disorders of iron metabolism. BioMetals 2005, 18, 339–348. [Google Scholar] [CrossRef]
- McKie, A.T.; Barrow, D.; Latunde-Dada, G.O.; Rolfs, A.; Sager, G.; Mudaly, E.; Richardson, C.; Barlow, D.; Bomford, A.; Peters, T.J.; et al. An iron-regulated ferric reductase associated with the absorption of dietary iron. Science 2001, 291, 1755–1759. [Google Scholar] [CrossRef]
- Donovan, A.; Brownlie, A.; Dorschner, M.O.; Zhou, Y.; Pratt, S.J.; Paw, B.H.; Phillips, R.B.; Thisse, C.; Thisse, B.; Zon, L.I. The zebrafish mutant gene chardonnay (cdy) encodes divalent metal transporter 1 (DMT-1). Blood 2002, 100, 4655–4659. [Google Scholar] [CrossRef] [Green Version]
- Lipiński, P.; Starzyński, R.R.; Gajowiak, A.; Staroń, R. Iron Metabolism- state of the art. Kosm Ser A 2014, 63, 299–308. [Google Scholar]
- Wang, X.; Flores, S.R.; Ha, J.-H.; Doguer, C.; Woloshun, R.R.; Xiang, P.; Grosche, A.; Vidyasagar, S.; Collins, J.F. Intestinal DMT1 Is Essential for Optimal Assimilation of Dietary Copper in Male and Female Mice with Iron-Deficiency Anemia. J. Nutr. 2018, 148, 1244–1252. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, A.; Le Blanc, S.; Olivares, M.; Pizarro, F.; Ruz, M.; Arredondo, M. Iron, copper, and zinc transport: Inhibition of divalent metal transporter 1 (DMT 1) and human copper transporter 1 (hCTR1) by shRNA. Biol. Trace Elem. Res. 2012, 146, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Ha, J.-H.; Doguer, C.; Collins, J.F. Knockdown of copper-transporting ATPase 1 (Atp7a) impairs iron flux in fully-differentiated rat (IEC-6) and human (Caco-2) intestinal epithelial cells. Metallomics 2016, 8, 963–972. [Google Scholar] [CrossRef] [Green Version]
- Latunde-Dada, G.O.; Takeuchi, K.; Simpson, R.J.; McKie, A.T. Haem carrier protein 1 (HCP1): Expression and functional studies in cultured cells. FEBS Lett. 2006, 580, 6865–6870. [Google Scholar] [CrossRef] [Green Version]
- Macara, I.G.; Hoy, T.G.; Harrison, P.M. The formation of ferritin from apoferritin. Catalytic action of apoferritin. Biochem. J. 1973, 135, 343. [Google Scholar] [CrossRef] [Green Version]
- Doguer, C.; Ha, J.-H.; Collins, J.F. Intersection of iron and copper metabolism in the mammalian intestine and liver. Compr. Physiol. 2018, 8, 1433. [Google Scholar] [CrossRef]
- Qiu, A.; Jansen, M.; Sakaris, A.; Min, S.H.; Chattopadhyay, S.; Tsai, E.; Sandoval, C.; Zhao, R.; Akabas, M.; Goldman, I.D. Identification of an Intestinal Folate Transporter and the Mol.ecular Basis for Hereditary Folate Malabsorption. Cell 2006, 127, 917–928. [Google Scholar] [CrossRef] [Green Version]
- Le, N.T.; Richardson, D.R. Ferroportin1, a new iron export molecule? Int. J. Biochem. Cell Biol. 2002, 34, 103–108. [Google Scholar] [CrossRef]
- Vulpe, C.D.; Kuo, Y.-M.; Murphy, T.L.; Cowley, L.; Askwith, C.; Libina, N.; Gitschier, J.; Anderson, G.J. Hephaestin, a ceruloplasmin homologue implicated in intestinal iron transport, is defective in the sla mouse. Nat. Genet. 1999, 21, 195–199. [Google Scholar] [CrossRef]
- Donovan, A.; Brownlie, A.; Zhou, Y.; Shepard, J.; Pratt, S.J.; Moynihan, J.; Paw, B.H.; Drejer, A.; Barut, B.; Zapata, A.; et al. Positional cloning of zebrafish ferroportin1 identifies a conserved vertebrate iron exporter. Nature 2000, 403, 776–781. [Google Scholar] [CrossRef] [PubMed]
- Daher, R.; Karim, Z. Iron metabolism: State of the art. Transfus Clin. Biol. 2017, 24, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Giansanti, F.; Panella, G.; Leboffe, L.; Antonini, G. Lactoferrin from Milk: Nutr. aceutical and Pharmacol.ogical Properties. Pharmaceuticals 2016, 9, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nairz, M.; Theurl, I.; Swirski, F.K.; Weiss, G. “Pumping iron”—How macrophages handle iron at the systemic, microenvironmental, and cellular levels. Pflugers Arch. 2017, 469, 397–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukhbaatar, N.; Weichhart, T. Iron Regulation: Macrophages in Control. Pharmaceuticals 2018, 11, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolas, G.; Bennoun, M.; Devaux, I.; Beaumont, C.; Grandchamp, B.; Kahn, A.; Vaulont, S. Lack of hepcidin gene expression and severe tissue iron overload in upstream stimulatory factor 2 (USF2) knockout mice. Proc. Natl. Acad. Sci. USA 2001, 98, 8780–8785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pigeon, C.; Ilyin, G.; Courselaud, B.; Leroyer, P.; Turlin, B.; Brissot, P.; Loréal, O. A new mouse liver-specific gene, encoding a protein homologous to human antimicrobial peptide hepcidin, is overexpressed during Iron overload. J. Biol. Chem. 2001, 276, 7811–7819. [Google Scholar] [CrossRef] [Green Version]
- Barrett, J.F.; Whittaker, P.G.; Williams, J.G.; Lind, T. Absorption of non-haem iron from food during normal pregnancy. BMJ 1994, 309, 79–82. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganze, T.; Kaplan, J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science 2004, 306, 2090–2093. [Google Scholar] [CrossRef] [Green Version]
- Wada, H.G.; Hass, P.E.; Sussman, H.H. Transferrin receptor in human placental brush border membranes. J. Biol. Chem. 1979, 254, 12629–12635. [Google Scholar]
- Müller, R.; Verma, I.M.; Adamson, E.D. Expression of c-onc genes: C-fos transcripts accumulate to high levels during development of mouse placenta, yolk sac and amnion. EMBO J. 1983, 2, 679–684. [Google Scholar] [CrossRef] [PubMed]
- Gambling, L.; Danzeisen, R.; Gair, S.; Lea, R.G.; Charania, Z.; Solanky, N.; Joory, K.D.; Srai, S.K.; McArdle, H.J. Effect of iron deficiency on placental transfer of iron and expression of iron transport proteins in vivo and in vitro. Biochem. J. 2001, 356 (Pt 3), 883–889. [Google Scholar] [CrossRef]
- Bastin, J.; Drakesmith, H.; Rees, M.; Sargent, I.; Townsend, A. Localisation of proteins of iron metabolism in the human placenta and liver. Br. J. Haematol. 2006, 134, 532–543. [Google Scholar] [CrossRef] [PubMed]
- Ohgami, R.S.; Campagna, D.R.; McDonald, A.; Fleming, M.D. The Steap proteins are metalloreductases. Blood 2006, 108, 1388–1394. [Google Scholar] [CrossRef] [PubMed]
- Georgieff, M.K.; Wobken, J.K.; Welle, J.; Burdo, J.R.; Connor, J.R. Identification and localization of divalent metal transporter-1 (DMT-1) in term human placenta. Placenta 2000, 21, 799–804. [Google Scholar] [CrossRef] [PubMed]
- McArdle, H.J.; Gambling, L.; Kennedy, C. Iron deficiency during pregnancy: The consequences for placental function and fetal outcome. Proc Nutr. Soc. 2014, 73, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Gunshin, H.; Fujiwara, Y.; Custodio, A.O.; Direnzo, C.; Robine, S.; Andrews, N.C. Slc11a2 is required for intestinal iron absorption and erythropoiesis but dispensable in placenta and liver. J. Clin. Invest. 2005, 115, 1258–1266. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.-Y.; Jenkitkasemwong, S.; Duarte, S.; Sparkman, B.K.; Shawki, A.; Mackenzie, B.; Knutson, M.D. ZIP8 is an iron and zinc transporter whose cell-surface expression is up-regulated by cellular iron loading. J. Biol. Chem. 2012, 287, 34032–34043. [Google Scholar] [CrossRef] [Green Version]
- Cao, C.; Fleming, M.D. The placenta: The forgotten essential organ of iron transport. Nutr. Rev. 2016, 74, 421–431. [Google Scholar] [CrossRef] [Green Version]
- Sangkhae, V.; Nemeth, E. Placental iron transport: The mechanism and regulatory circuits. Free Radic Biol. Med. 2019, 133, 254–261. [Google Scholar] [CrossRef]
- Kohgo, Y.; Ikuta, K.; Ohtake, T.; Torimoto, Y.; Kato, J. Body iron metabolism and pathophysiology of iron overload. Int. J. Hematol. 2008, 88, 7–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, D.F. The Regulation of Iron Absorption and Homeostasis. Clin. Biochem. Rev. 2016, 37, 51–62. [Google Scholar] [PubMed]
- Baker, L.B. Physiology of sweat gland function: The roles of sweating and sweat composition in human health. Temp. Multidiscip Biomed. J. 2019, 6, 211–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gulec, S.; Anderson, G.J.; Collins, J.F. Mechanistic and regulatory aspects of intestinal iron absorption. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bothwell, T.H. Iron requirements in pregnancy and strategies to meet them. Am. J. Clin. Nutr. 2000, 72, 257S–264S. [Google Scholar] [CrossRef]
- Johnson-Wimbley, T.D.; Graham, D.Y. Diagnosis and management of iron deficiency anemia in the 21st century. Therap. Adv. Gastroenterol. 2011, 4, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Soppi, E.T. Iron deficiency without anemia—A clinical challenge. Clin. Case Rep. 2018, 6, 1082–1086. [Google Scholar] [CrossRef]
- Rüfer, A.; Criblez, D.; Wuillemin, W.A. Iron-deficiency anemia and gastrointestinal bleeding. Ther. Umschau. 2006, 63, 339–343. [Google Scholar] [CrossRef]
- Spottiswoode, N.; Duffy, P.E.; Drakesmith, H. Iron, anemia and hepcidin in malaria. Front Pharmacol. 2014, 5, 125. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.L.; Brooker, S. Impact of hookworm infection and deworming on anaemia in non-pregnant populations: A systematic review. Trop. Med. Int. Health 2010, 15, 776–795. [Google Scholar] [CrossRef]
- Miller, J.L. Iron deficiency anemia: A common and curable disease. Cold Spring Harb. Perspect. Med. 2013, 3, a011866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartnikas, T.B. Known and potential roles of transferrin in iron biology. Biometals 2012, 25, 677–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naoum, F.A. Iron deficiency in cancer patients. Rev. Bras. Hematol. Hemoter. 2016, 38, 325–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.I.; Okam, M.M. Anemia in pregnancy. Hematol. Oncol Clin. N. Am. 2011, 25, 241–259. [Google Scholar] [CrossRef] [PubMed]
- Charzewska, J. Human NutritionBasics of Nutrition Science; Gawęcki, J., Ed.; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2000; pp. 481–492. [Google Scholar]
- Loréal, O.; Cavey, T.; Bardou-Jacquet, E.; Guggenbuhl, P.; Ropert, M.; Brissot, P. Iron, hepcidin, and the metal connection. Front. Pharmacol. 2014, 5, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menon, A.V.; Chang, J.; Kim, J. Mechanisms of Divalent Metal Toxicity in Affective Disorders. Toxicology 2016, 339, 58–72. [Google Scholar] [CrossRef] [Green Version]
- Hubbard, A.C.; Bandyopadhyay, S.; Wojczyk, B.S.; Spitalnik, S.L.; Hod, E.A.; Prestia, K.A. Effect of dietary iron on fetal growth in pregnant mice. Comp. Med. 2013, 63, 127–135. [Google Scholar]
- Woodman, A.G.; Mah, R.; Keddie, D.; Noble, R.M.N.; Panahi, S.; Gragasin, F.S.; Lemieux, H.; Bourque, S.L. Prenatal iron deficiency causes sex-dependent mitochondrial dysfunction and oxidative stress in fetal rat kidneys and liver. FASEB J. 2018, 32, 3254–3263. [Google Scholar] [CrossRef] [Green Version]
- Harrison-Findik, D.D. Gender-related variations in iron metabolism and liver diseases. World J. Hepatol. 2010, 2, 302–310. [Google Scholar] [CrossRef]
- Dignass, A.; Farrag, K.; Stein, J. Limitations of serum ferritin in diagnosing iron deficiency in inflammatory conditions. Int. J. Chronic Dis. 2018, 2018, 9394060. [Google Scholar] [CrossRef] [Green Version]
- Hallberg, L. Vitamins and Minerals in Pregnancy and Lactation; Berger, H., Ed.; Raven Press: New York, NY, USA, 1988; pp. 115–127. [Google Scholar]
- Seu, M.; Mose, J.C.; Panigoro, R.; Sahiratmadja, E. Anemia prevalence after iron supplementation among pregnant women in midwifes practice of primary health care facilities in eastern indonesia. Anemia 2019, 2019, 1413906. [Google Scholar] [CrossRef] [PubMed]
- McMahon, L.P. Iron deficiency in pregnancy. Obstet. Med. 2010, 3, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Tangeda, P.R.; Patil, S.; Shastri, N.; Noorali, S.N. Maternal myocardial performance in second trimester of pregnancy with iron deficiency anaemia. J. Clin. Diagn Res. 2016, 10, CC16. [Google Scholar] [CrossRef]
- Woodman, A.G.; Care, A.S.; Mansour, Y.; Cherak, S.J.; Panahi, S.; Gragasin, F.S.; Bourque, S.L. Modest and severe maternal iron deficiency in pregnancy are associated with fetal anaemia and organ-specific hypoxia in rats. Sci. Rep. 2017, 7, 46573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tayyebi, A.; Poursadeghfard, M.; Nazeri, M.; Pousadeghfard, T. Is there any correlation between migraine attacks and iron deficiency anemia? A case-control study. Int. J. Hematol. Stem Cell Res. 2019, 13, 164–171. [Google Scholar]
- Abu-Ouf, N.M.; Jan, M.M. The impact of maternal iron deficiency and iron deficiency anemia on child’s health. Saudi Med. J. 2015, 36, 146–149. [Google Scholar] [CrossRef]
- Lumish, R.A.; Young, S.L.; Lee, S.; Cooper, E.; Pressman, E.; Guillet, R.; O’Brien, K.O. Gestational iron deficiency is associated with pica behaviors in adolescents. J. Nutr. 2014, 144, 1533–1539. [Google Scholar] [CrossRef] [Green Version]
- Breymann, C. Iron deficiency anemia in pregnancy. Semin Hematol. 2015, 52, 339–347. [Google Scholar] [CrossRef]
- Milman, N.; Taylor, C.L.; Merkel, J.; Brannon, P.M. Iron status in pregnant women and women of reproductive age in Europe. Am. J. Clin. Nutr. 2017, 106, 1655S–1662S. [Google Scholar] [CrossRef]
- Tandon, R.; Jain, A.; Malhotra, P. Management of iron deficiency anemia in pregnancy in india. Indian J. Hematol. Blood Transfus. 2018, 34, 204. [Google Scholar] [CrossRef]
- Milman, N. Iron and pregnancy—A delicate balance. Ann. Hematol. 2006, 85, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.B.; Hurrell, R.F. Nutritional iron deficiency. Lancet 2007, 370, 511–520. [Google Scholar] [CrossRef]
- Black, R.E. Micronutrients in pregnancy. Br. J. Nutr. 2001, 85, S193–S197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueiredo, A.C.M.G.; Gomes-Filho, I.S.; Silva, R.B.; Pereira, P.P.S.; Mata, F.A.F.; Da Lyrio, A.O.; Souza, E.S.; Cruz, S.S.; Pereira, M.G. Maternal anemia and low birth weight: A systematic review and meta-analysis. Nutrients 2018, 10, 601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ru, Y.; Pressman, E.K.; Cooper, E.M.; Guillet, R.; Katzman, P.J.; Kent, T.R.; Bacak, S.J.; O’Brien, K.O. Iron deficiency and anemia are prevalent in women with multiple gestations. Am. J. Clin. Nutr. 2016, 104, 1052–1060. [Google Scholar] [CrossRef]
- Kaiser, L.; Allen, L.H.; American Dietetic Association. Position of the American Dietetic Association: Nutrition and lifestyle for a healthy pregnancy outcome. J. Am. Diet. Assoc. 2008, 108, 553–561. [Google Scholar]
- Khayat, S.; Fanaei, H.; Ghanbarzehi, A. Minerals in pregnancy and lactation: A review Article. J. Clin. Diagn Res. 2017, 11, QE01–QE05. [Google Scholar] [CrossRef]
- Cogswell, M.E.; Parvanta, I.; Ickes, L.; Yip, R.; Brittenham, G.M. Iron supplementation during pregnancy, anemia, and birth weight: A randomized controlled trial. Am. J. Clin. Nutr. 2003, 78, 773–781. [Google Scholar] [CrossRef] [Green Version]
- Allen, L.H. Biological mechanisms that might underlie iron’s effects on fetal growth and preterm birth. J. Nutr. 2001, 131, 581S–589S. [Google Scholar] [CrossRef] [Green Version]
- Peña-Rosas, J.P.; De-Regil, L.M.; Garcia-Casal, M.N.; Dowswell, T. Daily oral iron supplementation during pregnancy. Cochrane Database Syst Rev. 2015, 7, CD004736. [Google Scholar] [CrossRef] [Green Version]
- Abioye, A.I.; Aboud, S.; Premji, Z.; Etheredge, A.J.; Gunaratna, N.S.; Sudfeld, C.R.; Mongi, R.; Meloney, L.; Darling, A.M.; Noor, R.A.; et al. Iron supplementation affects hematologic biomarker concentrations and pregnancy outcomes among iron-deficient Tanzanian women. J. Nutr. 2016, 146, 1162–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, G.; Xu, G.; Zhou, M.; Jiang, Y.; Richards, B.; Clark, K.M.; Kaciroti, N.; Georgieff, M.K.; Zhang, Z.; Tardif, T.; et al. Prenatal iron supplementation reduces maternal anemia, iron deficiency, and iron deficiency anemia in a randomized clinical trial in rural china, but iron deficiency remains widespread in mothers and neonates. J. Nutr. 2015, 145, 1916–1923. [Google Scholar] [CrossRef] [PubMed]
- Peña-Rosas, J.P.; De-Regil, L.M.; Gomez, M.H.; Flores-Urrutia, M.C.; Dowswell, T. Intermittent oral iron supplementation during pregnancy. Cochrane Database Syst. Rev. 2015, CD009997. [Google Scholar] [CrossRef] [PubMed]
- Ziaei, S.; Rahman, A.; Raqib, R.; Lönnerdal, B.; Ekström, E.-C. A prenatal multiple micronutrient supplement produces higher maternal vitamin B-12 concentrations and similar folate, ferritin, and zinc concentrations as the standard 60-mg iron plus 400-μg folic acid supplement in rural bangladeshi women. J. Nutr. 2016, 146, 2520–2529. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.K.; Abbas, A.M.; Abdelmagied, A.M.; Mohammed, G.E.; Abdalmageed, O.S. A randomized clinical trial of the efficacy of single versus double-daily dose of oral iron for prevention of iron deficiency anemia in women with twin gestations. J. Matern. Fetal Neonatal Med. 2017, 30, 2884–2889. [Google Scholar] [CrossRef]
- Shinar, S.; Skornick-Rapaport, A.; Maslovitz, S. Iron supplementation in twin pregnancy—The benefit of doubling the iron dose in iron deficient pregnant women: A randomized controlled trial. Twin Res. Hum. Genet. 2017, 20, 419–424. [Google Scholar] [CrossRef] [Green Version]
- Skoczynska, A.; Kwiecinska, D.; Kielbinski, M.; Lukaszewski, M. Acute iron poisoning in adult female. Hum. Exp. Toxicol. 2007, 26, 663–666. [Google Scholar] [CrossRef]
- Wood, J.C. Guidelines for quantifying iron overload. ASH Educ. Progr. B 2014, 2014, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Brissot, P.; Ropert, M.; Le Lan, C.; Loréal, O. Non-transferrin bound iron: A key role in iron overload and iron toxicity. Biochim. Biophys Acta Gen. Subj. 2012, 1820, 403–410. [Google Scholar] [CrossRef]
- Aydemir, T.B.; Cousins, R.J. The Multiple Faces of the Metal Transporter ZIP14 (SLC39A14). J. Nutr. 2018, 148, 174–184. [Google Scholar] [CrossRef]
- Fibach, E.; Rachmilewitz, E.A. Iron overload in hematological disorders. Presse Med. 2017, 46, e296–e305. [Google Scholar] [CrossRef] [PubMed]
- Pietrangelo, A. Iron and the liver. Liver Int. 2016, 36, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Silver, M.M.; Beverley, D.W.; Valberg, L.S.; Cutz, E.; Phillips, M.J.; Shaheed, W.A. Perinatal hemochromatosis. Clinical, morphologic, and quantitative iron studies. Am. J. Pathol. 1987, 128, 538. [Google Scholar] [PubMed]
- Prus, E.; Fibach, E. Effect of iron chelators on labile iron and oxidative status of thalassaemic erythroid cells. Acta Haematol. 2010, 123, 14–20. [Google Scholar] [CrossRef]
- Menon, R. Oxidative stress damage as a detrimental factor in preterm birth pathology. Front. Immunol. 2014, 5, 567. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.F. Copper. In Modern Nutrition in Health and Disease, 11th ed.; Ross, A.C., Caballero, B., Cousins, R.J., Tucker, K.L., Ziegler, T.R., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2014; p. 206. [Google Scholar]
- Gibson, R.S. Principles of Nutritional Assessment; Oxford University Press: Oxford, UK, 2005; 908p. [Google Scholar]
- Łanocha-Arendarczyk, N.; Kosik-Bogacka, D.I.; Copper, C. Mammals and Birds as Bioindicators of Trace Element Contaminations in Terrestrial Environments: An Ecotoxicological Assessment of the Northern Hemisphere; Kalisińska, E., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 125–161. [Google Scholar]
- Uauy, R.; Olivares, M.; Gonzalez, M. Essentiality of copper in humans. Am. J. Clin. Nutr. 1998, 67, 952S–959S. [Google Scholar] [CrossRef]
- Cetin, I.; Berti, C.; Calabrese, S. Role of micronutrients in the periconceptional period. Hum. Reprod Update. 2010, 16, 80–95. [Google Scholar] [CrossRef] [Green Version]
- Lurie, D.G.; Holden, J.M.; Schubert, A.; Wolf, W.R.; Miller-Ihli, N.J. The copper content of foods based on a critical evaluation of published analytical data. J. Food Compos. Anal. 1989, 2, 298–316. [Google Scholar] [CrossRef]
- Squitti, R.; Siotto, M.; Polimanti, R. Low-copper diet as a preventive strategy for Alzheimer’s disease. Neurobiol. Aging 2014, 35, S40–S50. [Google Scholar] [CrossRef] [Green Version]
- Peña, M.M.O.; Lee, J.; Thiele, D.J. A delicate balance: Homeostatic control of copper uptake and distribution. J. Nutr. 1999, 129, 1251–1260. [Google Scholar] [CrossRef] [Green Version]
- Krzeptowski, W.; Pierzchała, O.; Lenartowicz, M. Copper metabolism and characteristic of inherited metabolic syndromes caused by copper deficiency and lack of TP7A protein activity. Kosmos 2014, 63, 395–413. [Google Scholar]
- Wang, F.; Jiao, P.; Qi, M.; Frezza, M.; Dou, Q.P.; Yan, B. Turning tumor-promoting copper into an anti-cancer weapon via high-throughput chemistry. Curr Med. Chem. 2010, 17, 2685–2698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisses, J.F.; Kaplan, J.H. The mechanism of copper uptake mediated by human CTR1, a mutational analysis. J. Biol. Chem. 2005, 280, 37159–37168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunshin, H.; Mackenzie, B.; Berger, U.V.; Gunshin, Y.; Romero, M.F.; Boron, W.F.; Nussberger, S.; Gollan, J.L.; Hediger, M.A. Cloning and characterization of a proton-coupled mammalian metal-ion transporter. Nature 1997, 388, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Wu, X.; Lee, J. SLC31 (CTR) family of copper transporters in health and disease. Mol. Aspects Med. 2013, 34, 561–570. [Google Scholar] [CrossRef] [Green Version]
- Sharp, P.A. Ctr1 and its role in body copper homeostasis. Int. J. Biochem. Cell Biol. 2003, 35, 288–291. [Google Scholar] [CrossRef]
- Wee, N.K.Y.; Weinstein, D.C.; Fraser, S.T.; Assinder, S.J. The mammalian copper transporters CTR1 and CTR2 and their roles in development and disease. Int. J. Biochem. Cell Biol. 2013, 45, 960–963. [Google Scholar] [CrossRef]
- Kuo, Y.M.; Zhou, B.; Cosco, D.; Gitschier, J. The copper transporter CTR1 provides an essential function in mammalian embryonic development. Proc. Natl. Acad. Sci. USA 2001, 98, 6836–6841. [Google Scholar] [CrossRef] [Green Version]
- Kelner, G.S.; Lee, M.; Clark, M.E.; Maciejewski, D.; McGrath, D.; Rabizadeh, S.; Lyons, T.; Bredesen, D.; Jenner, P.; Maki, R.A. The copper transport protein Atox1 promotes neuronal survival. J. Biol. Chem. 2000, 275, 580–584. [Google Scholar] [CrossRef] [Green Version]
- Shike, M. Copper in Parenteral Nutrition. Gastroenterology 2009, 137, S13–S17. [Google Scholar] [CrossRef] [Green Version]
- Kunachowicz, H.; Przygoda, B.; Nadolna, I.K. Food Composition and Nutrition Tables; PZWL: Warszawa, Poland, 2017. [Google Scholar]
- McArdle, H.J.; Bingham, M.J.; Summer, K.; Ong, T.J. Cu metabolism in the liver. Adv. Exp. Med. Biol. 1999, 448, 29–37. [Google Scholar] [PubMed]
- Pisu, P.; Bellovino, D.; Gaetani, S. Copper regulated synthesis, secretion and degradation of ceruloplasmin in a mouse immortalized hepatocytic cell line. Cell Mol. Biol. 2005, 51, OL859–OL867. [Google Scholar] [PubMed]
- Boyer, J.L. Bile Formation and Secretion. Compr. Physiol. 2013, 3, 1035. [Google Scholar]
- Moore, S.D.; Cox, D.W. Expression in Mouse Kidney of Membrane Copper Transporters Atp7a and Atp7b. Nephron 2002, 92, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Frankel, D.A. Supplementation of trace elements in parenteral nutrition: Rationale and recommendations. Nutr. Res. 1993, 13, 583–596. [Google Scholar] [CrossRef]
- Bartee, M.Y.; Lutsenko, S. Hepatic copper-transporting ATPase ATP7B: Function and inactivation at the molecular and cellular level. Biometals 2007, 20, 627–637. [Google Scholar] [CrossRef]
- Lee, J.; Prohaska, J.R.; Thiele, D.J. Essential role for mammalian copper transporter Ctr1 in copper homeostasis and embryonic development. Proc. Natl. Acad. Sci. USA 2001, 98, 6842–6847. [Google Scholar] [CrossRef] [Green Version]
- Hardman, B.; Manuelpillai, U.; Wallace, E.M.; van de Waasenburg, S.; Cater, M.; Mercer, J.F.B.; Ackland, M.L. Expression and localization of Menkes and Wilson copper transporting ATPases in human placenta. Placenta 2004, 25, 512–517. [Google Scholar] [CrossRef] [Green Version]
- Hardman, B.; Michalczyk, A.; Greenough, M.; Camakaris, J.; Mercer, J.F.B.; Ackland, M.L. Hormonal regulation of the Menkes and Wilson copper-transporting ATPases in human placental Jeg-3 cells. Biochem. J. 2007, 402, 241–250. [Google Scholar] [CrossRef]
- McArdle, H.J.; Andersen, H.S.; Jones, H.; Gambling, L. Copper and iron transport across the placenta: Regulation and Interactions. J. Neuroendocrinol. 2008, 20, 427–431. [Google Scholar] [CrossRef]
- Collins, J.F.; Klevay, L.M. Copper. Adv. Nutr. 2011, 2, 520–522. [Google Scholar] [CrossRef] [PubMed]
- Wazir, S.M.; Ghobrial, I. Copper deficiency, a new triad: Anemia, leucopenia, and myeloneuropathy. J. Community Hosp. Intern. Med. Perspect. 2017, 7, 265–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myint, Z.W.; Oo, T.H.; Thein, K.Z.; Tun, A.M.; Saeed, H. Copper deficiency anemia: Review article. Ann. Hematol. 2018, 97, s00277–s018. [Google Scholar] [CrossRef] [PubMed]
- Bolognin, S.; Pasqualetto, F.; Mucignat-Caretta, C.; Scancar, J.; Milacic, R.; Zambenedetti, P.; Cozzi, B.; Zatta, P. Effects of a copper-deficient diet on the biochemistry, neural morphology and behavior of aged mice. PLoS ONE 2012, 7, e47063. [Google Scholar] [CrossRef] [PubMed]
- Zatta, P.; Frank, A. Copper deficiency and neurological disorders in man and animals. Brain Res. Rev. 2007, 54, 19–33. [Google Scholar] [CrossRef]
- Chen, Y.; Saari, J.T.; Kang, Y.J. Weak antioxidant defenses make the heart a target for damage in copper-deficient rats. Free Radic Biol. Med. 1994, 17, 529–536. [Google Scholar] [CrossRef]
- Qu, X.; He, Z.; Qiao, H.; Zhai, Z.; Mao, Z.; Yu, Z.; Dai, K. Serum copper levels are associated with bone mineral density and total fracture. J. Orthop. Transl. 2018, 14, 34. [Google Scholar] [CrossRef]
- Wang, H.W.; Zhou, B.H.; Zhang, S.; Guo, H.W.; Zhang, J.L.; Zhao, J.; Tian, E.J. Reproductive toxicity in male mice after exposure to high molybdenum and low copper concentrations. Toxicol. Ind. Health 2016, 32, 1598–1606. [Google Scholar] [CrossRef]
- Li, Y.; Wang, L.; Schuschke, D.A.; Zhou, Z.; Saari, J.T.; Kang, Y.J. Marginal dietary copper restriction induces cardiomyopathy in rats. J. Nutr. 2005, 135, 2130–2136. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Reynolds, C.; Xiao, C.; Feng, W.; Zhou, Z.; Rodriguez, W.; Tyagi, S.C.; Eaton, J.W.; Saari, J.T.; Kang, Y.J. Dietary copper supplementation reverses hypertrophic cardiomyopathy induced by chronic pressure overload in mice. J. Exp. Med. 2007, 204, 657. [Google Scholar] [CrossRef]
- WHO. Copper in Drinking-Water; WHO/SDE/WSH/03.04/88:2004; World Health Organization: Geneva, Switzerland, 2004. [Google Scholar]
- Danks, D.M. Copper Deficiency in Humans. Annu. Rev. Nutr. 1988, 8, 235–257. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-Z.; Li, M.; Huang, C.-Y.; Zhang, Y.-Z. Study of effects of copper deficiency on internal organ through a copper deficiency model in rat. Zhonghua Yu Fang Yi Xue Za Zhi 2007, 41, 127–130. [Google Scholar] [PubMed]
- Kiilholma, P.; Grönroos, M.; Erkkola, R.; Pakarinen, P.; Näntö, V. The role of calcium, copper, iron and zinc in preterm delivery and premature rupture of fetal membranes. Gynecol. Obstet. Invest. 1984, 17, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Kardos, J.; Héja, L.; Simon, Á.; Jablonkai, I.; Kovács, R.; Jemnitz, K. Copper signalling: Causes and consequences. Cell Commun. Signal. 2018, 16, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Liou, I.W.; Biggins, S.W.; Yeh, M.; Jalikis, F.; Chan, L.; Burkhead, J. Copper deficiency in liver diseases: A case series and pathophysiological considerations. Hepatol. Commun. 2019, 3, 1159. [Google Scholar] [CrossRef] [Green Version]
- Tümer, Z. An Overview and Update of ATP7A Mutations leading to Menkes disease and occipital horn syndrome. Hum. Mutat. 2013, 34, 417–429. [Google Scholar] [CrossRef]
- Tümer, Z.; Møller, L.B. Menkes disease. Eur. J. Hum. Genet. 2010, 18, 511–518. [Google Scholar] [CrossRef] [Green Version]
- Møller, L.; Lenartowicz, M.; Zabot, M.-T.; Josiane, A.; Burglen, L.; Bennett, C.; Riconda, D.; Fisher, R.; Janssens, S.; Mohammed, S.; et al. Clinical expression of Menkes disease in females with normal karyotype. Orphanet J. Rare Dis. 2012, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Gualandi, F.; Sette, E.; Fortunato, F.; Bigoni, S.; De Grandis, D.; Scotton, C.; Selvatici, R.; Neri, M.; Incensi, A.; Liguori, R.; et al. Report of a novel ATP7A mutation causing distal motor neuropathy. Neuromuscul. Disord. 2019, 29, 776–785. [Google Scholar] [CrossRef]
- Chiplunkar, S.; Bindu, P.S.; Nagappa, M.; Bineesh, C.; Govindaraj, P.; Gayathri, N.; Srinivas Bharath, M.M.; Arvinda, H.R.; Mathuranath, P.S.; Sinha, S.; et al. Huppke-Brendel syndrome in a seven months old boy with a novel 2-bp deletion in SLC33A1. Metab. Brain Dis. 2016, 31, 1195–1198. [Google Scholar] [CrossRef]
- Martinelli, D.; Dionisi-Vici, C. AP1S1 defect causing MEDNIK syndrome: A new adaptinopathy associated with defective copper metabolism. Ann. N. Y. Acad. Sci. 2014, 1314, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Kashanian, M.; Hadizadeh, H.; Faghankhani, M.; Nazemi, M.; Sheikhansari, N. Evaluating the effects of copper supplement during pregnancy on premature rupture of membranes and pregnancy outcome. J. Matern. Fetal Neonatal Med. 2018, 31, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Jankowski, M.A.; Uriu-Hare, J.Y.; Rucker, R.B.; Rogers, J.M.; Keen, C.L. Maternal zinc deficiency, but not copper deficiency or diabetes, results in increased embryonic cell death in the rat: Implications for mechanisms underlying abnormal development. Teratology 1995, 51, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Kodama, H.; Fujisawa, C.; Bhadhprasit, W. Pathology, clinical features and treatments of congenital copper metabolic disorders—Focus on neurologic aspects. Brain Dev. 2011, 33, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Prohaska, J.R. Neurochemical roles of copper as antioxidant or prooxidant. In Metals and Oxidative Damage in Neurological Disorders; Springer: Boston, MA, USA, 1997; pp. 57–75. [Google Scholar]
- Lewandowska, M.; Sajdak, S.; Marciniak, W.; Lubiński, J. First trimester serum copper or zinc levels, and risk of pregnancy-induced hypertension. Nutrients 2019, 11, 2479. [Google Scholar] [CrossRef] [Green Version]
- Prohaska, J.R.; Brokate, B. The timming of perinatal copper deficiency in mice influences offspring survival. J. Nutr. 2002, 132, 3142–3145. [Google Scholar] [CrossRef] [Green Version]
- Vukelić, J.; Kapamadžija, A.; Petrović, D.; Grujić, Z.; Novakov-mikić, A.; Kopitović, V.; Bjelica, A. Variations of serum copper values in pregnancy. Srp. Arh. za Celok. Lek. 2012, 140, 42–46. [Google Scholar] [CrossRef]
- Hao, Y.; Pang, Y.; Yan, H.; Zhang, Y.; Liu, J.; Jin, L.; Yan, L.; Chen, D.; Hu, L.; Qin, Y.; et al. Association of maternal serum copper during early pregnancy with the risk of spontaneous preterm birth: A nested case-control study in China. Environ. Int. 2019, 122, 237–243. [Google Scholar] [CrossRef]
- Li, P.; Yin, J.; Zhu, Y.; Li, S.; Chen, S.; Sun, T.; Shan, Z.; Wang, J.; Shang, Q.; Li, X.; et al. Association between plasma concentration of copper and gestational diabetes mellitus. Clin. Nutr. 2019, 38, 2922–2927. [Google Scholar] [CrossRef]
- Gaetke, L.M.; Chow-Johnson, H.S.; Chow, C.K. Copper: Toxicological relevance and mechanisms. Arch. Toxicol. 2014, 88, 1929. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative stress: Harms and benefits for human health. Oxid. Med. Cell. Longev. 2017, 2017, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Rosencrantz, R.; Schilsky, M. Wilson Disease: Pathogenesis and clinical considerations in diagnosis and treatment. Semin. Liver Dis. 2011, 31, 245–259. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffenberger, J.; Beinhardt, S.; Gotthardt, D.N.; Haag, N.; Freissmuth, C.; Reuner, U.; Gauss, A.; Stremmel, W.; Schilsky, M.L.; Ferenci, P.; et al. Pregnancy in Wilson’s disease: Management and outcome. Hepatology 2018, 67, 1261–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reuner, U.; Dinger, J. Pregnancy and Wilson disease: Management and outcome of mother and newborns—Experiences of a perinatal centre. Ann. Transl. Med. 2019, 7, 56. [Google Scholar] [CrossRef]
- Russ, E.M.; Raymunt, J. Influence of estrogens on total serum copper and caeruloplasmin. Exp. Biol. Med. 1956, 92, 465–466. [Google Scholar] [CrossRef]
- Kambe, T.; Tsuji, T.; Hashimoto, A.; Itsumura, N. The physiological, biochemical, and molecular roles of zinc transporters in zinc homeostasis and metabolism. Physiol. Rev. 2015, 95, 749–784. [Google Scholar] [CrossRef]
- Rink, L.; Gabriel, P. Zinc and the immune system. Proc. Nutr. Soc. 2000, 59, 541–552. [Google Scholar] [CrossRef] [Green Version]
- Wessells, K.R.; Jorgensen, J.M.; Hess, S.Y.; Woodhouse, L.R.; Peerson, J.M.; Brown, K.H. Plasma zinc concentration responds rapidly to the initiation and discontinuation of short-term zinc supplementation in healthy men. J. Nutr. 2010, 140, 2128–2133. [Google Scholar] [CrossRef] [Green Version]
- Hess, S.Y.; Peerson, J.M.; King, J.C.; Brown, K.H. Use of serum zinc concentration as an indicator of population zinc status. Food Nutr. Bull. 2007, 28 (Suppl. 3), S403–S429. [Google Scholar] [CrossRef]
- Sekler, I.; Sensi, S.L.; Hershfinkel, M.; Silverman, W.F. Mechanism and regulation of cellular zinc transport. Mol. Med. 2007, 13, 337. [Google Scholar] [CrossRef]
- Jackson, M.J. Physiology of zinc: General aspects. In Zinc in Human Biology; Mills, C.F., Ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1989; pp. 1–14. [Google Scholar]
- Maret, W. Zinc Biochemistry, Physiology, and Homeostasis; Springer: Dordrecht, The Netherlands, 2001. [Google Scholar]
- Gower-Winter, S.D.; Levenson, C.W. Zinc in the central nervous system: From molecules to behavior. BioFactors 2012, 38, 186–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandstead, H.H.; Frederickson, C.J.; Penland, J.G. History of zinc as related to brain function. J. Nutr. 2000, 130, 496S–502S. [Google Scholar] [CrossRef] [PubMed]
- Merck, V.M. The Merck Veterinary Manual: A Handbook of Diagnosis, Therapy and Disease Prevention and Control for the Veterinarian, 6th ed.; Merck & Co., Inc.: Fort Kenalworth, NJ, USA, 1986. [Google Scholar]
- Barceloux, D.G. Zinc. J. Toxicol. Clin. Toxicol. 1999, 37, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Kay, A.R.; Toth, K. Is zinc a neuromodulator? Sci. Signal. 2008, 1, re3. [Google Scholar] [CrossRef]
- Mocchegiani, E.; Muzzioli, M.; Giacconi, R. Zinc, metallothioneins, immune responses, survival and ageing. Biogerontology 2000, 1, 133–143. [Google Scholar] [CrossRef]
- Solomons, N.W. Update on zinc biology. Ann. Nutr. Metab. 2013, 62, 8–17. [Google Scholar] [CrossRef]
- Hunt, C.D.; Johnson, P.E.; Herbel, J.; Mullen, L.K. Effects of dietary zinc depletion on seminal volume and zinc loss, serum testosterone concentrations, and sperm morphology in young men. Am. J. Clin. Nutr. 1992, 56, 148–157. [Google Scholar] [CrossRef]
- Dardenne, M.; Pleau, J.M.; Nabarra, B.; Lefrancier, P.; Derrien, M.; Choay, J.; Bach, J.F. Contribution of zinc and other metals to the biological activity of the serum thymic factor. Proc. Natl. Acad. Sci. USA 1982, 79, 5370–5373. [Google Scholar] [CrossRef] [Green Version]
- Gammoh, N.Z.; Rink, L. Zinc in Infection and Inflammation. Nutrients 2017, 9, 624. [Google Scholar] [CrossRef] [Green Version]
- Tainer, J.A.; Getzoff, E.D.; Richardson, J.S.; Richardson, D.C. Structure and mechanism of copper, zinc superoxide dismutase. Nature 1983, 306, 284–287. [Google Scholar] [CrossRef]
- Olechnowicz, J.; Tinkov, A.; Skalny, A.; Suliburska, J. Zinc status is associated with inflammation, oxidative stress, lipid, and glucose metabolism. J. Physiol. Sci. 2018, 68, 19–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boron, B.; Hupert, J.; Barch, D.H.; Fox, C.C.; Friedman, H.; Layden, T.J.; Mobarhan, S. Effect of zinc deficiency on hepatic enzymes regulating vitamin A status. J. Nutr. 1988, 118, 995–1001. [Google Scholar] [CrossRef]
- Zhai, Q.; Narbad, A.; Chen, W. Dietary strategies for the treatment of cadmium and lead toxicity. Nutrients 2015, 7, 552–571. [Google Scholar] [CrossRef] [Green Version]
- Gibson, R.S.; King, J.C.; Lowe, N. A review of dietary zinc recommendations. Food Nutr. Bull. 2016, 37, 443–460. [Google Scholar] [CrossRef] [Green Version]
- Maret, W. Zinc biochemistry: From a single zinc enzyme to a key element of life. Adv. Nutr. 2013, 4, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Zheng, Y.; Li, Y.; Shen, Z.; Tao, L.; Dou, X.; Qian, J.; Shen, H. Psychological stress induced zinc accumulation and up-regulation of ZIP14 and metallothionein in rat liver. BMC Gastroenterol. 2014, 14, 32. [Google Scholar] [CrossRef] [Green Version]
- Sas, B.; Bremner, I. Effect of acute stress on the absorption and distribution of zinc and on zn-metallothionein production in the liver of the chick. J. Inorg. Biochem. 1979, 11, 67–76. [Google Scholar] [CrossRef]
- Yonova, D.; Vazelov, E.; Tzatchev, K. Zinc status in patients with chronic renal failure on conservative and peritoneal dialysis treatment. Hippokratia 2012, 16, 356–359. [Google Scholar] [PubMed]
- Mohommad, M.K.; Zhou, Z.; Cave, M.; Barve, A.; McClain, C.J. Zinc and liver disease. Nutr. Clin. Pract. 2012, 27, 8. [Google Scholar] [CrossRef] [PubMed]
- Henderson, L.M.; Brewer, G.J.; Dressman, J.B.; Swidan, S.Z.; DuRoss, D.J.; Adair, C.H.; Barnett, J.L.; Berardi, R.R. Effect of intragastric pH on the absorption of oral zinc acetate and zinc oxide in young healthy volunteers. J. Parenter. Enteral Nutr. 1995, 19, 393–397. [Google Scholar] [CrossRef] [Green Version]
- Pallauf, J.; Kramer, K.; Markwitan, A.; Ebel, D. The effect of a supplement of citric acid on the bioavailability of zinc from corn germ. Z. Ernahr. 1990, 29, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Sandstrom, B.; Almgren, A.; Kivisto, B.; Cederblad, A. Effect of protein level and protein source on zinc absorption in humans. J. Nutr. 1989, 119, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Ghishan, F.K.; Stroop, S.; Meneely, R. The effect of lactose on the intestinal absorption of calcium and zinc in the rat during maturation. Pediatr. Res. 1982, 16, 566–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argiratos, V.; Samman, S. The effect of calcium carbonate and calcium citrate on the absorption of zinc in healthy female subjects. Eur. J. Clin. Nutr. 1994, 48, 198–204. [Google Scholar]
- Hambidge, K.M.; Miller, L.V.; Westcott, J.E.; Sheng, X.; Krebs, N.F. Zinc bioavailability and homeostasis. Am. J. Clin. Nutr. 2010, 91, 1478S–1483S. [Google Scholar] [CrossRef] [Green Version]
- Krittaphol, W.; Bailey, K.B.; Pongcharoen, T.; Winichagoon, P.; Gibson, R.S. Low zinc, iron, and calcium intakes of Northeast Thai school children consuming glutinous rice-based diets are not exacerbated by high phytate. Int. J. Food Sci. Nutr. 2006, 57, 520–528. [Google Scholar] [CrossRef]
- Sandström, B.; Davidsson, L.; Cederblad, Å.; Lönnerdal, B. Oral iron, dietary ligands and zinc absorption. J. Nutr. 1985, 115, 411–414. [Google Scholar] [CrossRef]
- Institute of Medicin (US) Panel on Micronutrients. Dietary reference intakes for vitamin, A.; vitamin, K. In Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc; National Academies Press (US): Washington, DC, USA, 2001. [Google Scholar]
- Giroux, E.; Schechter, P.J.; Schoun, J. Diminished albumin binding of zinc in serum of pregnant women. Clin. Sci. Mol. Med. 1976, 51, 545–549. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Food and Agriculture Organization of the United Nations, International Atomic Energy Agency. In Trace Elements in Human Nutrition and Health; World Health Organization: Geneva, Switzerland, 1996; 343p. [Google Scholar]
- King, J.C.; Shames, D.M.; Woodhouse, L.R. Zinc homeostasis in humans. J. Nutr. 2000, 130, 1360S–1366S. [Google Scholar] [CrossRef] [Green Version]
- Fukada, T.; Kambe, T. Molecular and genetic features of zinc transporters in physiology and pathogenesis. Met. Integr. Biomet. Sci. 2011, 3, 662–674. [Google Scholar] [CrossRef]
- Cousins, R.J.; McMahon, R.J. Integrative Aspects of Zinc Transporters. J. Nutr. 2000, 130, 1384S–1387S. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Chang, J.; Kim, J. Loss of divalent metal transporter 1 function promotes brain copper accumulation and increases impulsivity. J. Neurochem. 2016, 138, 918–928. [Google Scholar] [CrossRef] [PubMed]
- Turnlund, J.R.; King, J.C.; Keyes, W.R.; Gong, B.; Michel, M.C. A stable isotope study of zinc absorption in young men: Effects of phytate and a-cellulose. Am. J. Clin. Nutr. 1984, 40, 1071–1077. [Google Scholar] [CrossRef] [PubMed]
- Tapiero, H.; Tew, K.D. Trace elements in human physiology and pathology: Zinc and metallothioneins. Biomed. Pharmacother. 2003, 57, 399–411. [Google Scholar] [CrossRef]
- Smith, K.T.; Failla, M.L.; Cousins, R.J. Identification of albumin as the plasma carrier for zinc absorption by perfused rat intestine. Biochem. J. 1979, 184, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Gachot, B.; Tauc, M.; Morat, L.; Poujeol, P. Zinc uptake by proximal cells isolated from rabbit kidney: Effects of cysteine and histidine. Pflugers Arch. Eur. J. Physiol. 1991, 419, 583–587. [Google Scholar] [CrossRef]
- Aiken, S.P.; Horn, N.M.; Saunders, N.R. Effects of amino acids on zinc transport in rat erythrocytes. J. Physiol. 1992, 445, 69–80. [Google Scholar] [CrossRef]
- Coyle, P.; Philcox, J.C.; Carey, L.C.; Rofe, A.M. Metallothionein: The multipurpose protein. Cell Mol. Life Sci. 2002, 59, 627–647. [Google Scholar] [CrossRef]
- Drozd, A.; Wojewska, D.; Peris-Díaz, M.D.; Jakimowicz, P.; Krężel, A. Crosstalk of the structural and zinc buffering properties of mammalian metallothionein-2. Metallomics 2018, 10, 595–613. [Google Scholar] [CrossRef] [Green Version]
- Lynes, M.A.; Zaffuto, K.; Unfricht, D.W.; Marusov, G.; Samson, J.S.; Yin, X. The physiological roles of extracellular metallothionein. Exp. Biol. Med. 2006, 231, 1548–1554. [Google Scholar] [CrossRef]
- Ruttkay-Nedecky, B.; Nejdl, L.; Gumulec, J.; Zitka, O.; Masarik, M.; Eckschlager, T.; Stiborova, M.; Adam, V.; Kizek, R. The role of metallothionein in oxidative stress. Int. J. Mol. Sci. 2013, 14, 6044–6066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saad, A.A.; El-Sikaily, A.; Kassem, H. Metallothionein and glutathione content as biomarkers of metal pollution in mussels and local fishermen in Abu Qir Bay, Egypt. J. Heal. Pollut. 2016, 6, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, J.M.; Zweifach, A.; Lynes, M.A. Metallothionein regulates intracellular zinc signaling during CD4+ T cell activation. BMC Immunol. 2016, 17, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutherland, D.E.; Willans, M.J.; Stillman, M.J. Single domain metallothioneins: Supermetalation of human MT 1a. J. Am. Chem. Soc. 2012, 134, 3290–3299. [Google Scholar] [CrossRef]
- Krężel, A.; Maret, W. The functions of metamorphic metallothioneins in zinc and copper metabolism. Int. J. Mol. Sci. 2017, 18, 1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolas, G.; Bennoun, M.; Porteu, A.; Mativet, S.; Beaumont, C.; Grandchamp, B.; Sirito, M.; Sawadogo, M.; Kahn, A.; Vaulont, S. Severe iron deficiency anemia in transgenic mice expressing liver hepcidin. Proc. Natl. Acad. Sci. USA 2002, 99, 4596–4601. [Google Scholar] [CrossRef] [Green Version]
- Baltaci, A.K.; Yuce, K. Zinc transporter proteins. Neurochem. Res. 2018, 43, 517–530. [Google Scholar] [CrossRef]
- Liuzzi, J.P.; Cousins, R.J. Mammmalin zinc transporters. Annu. Rev. Nutr. 2004, 24, 151–172. [Google Scholar] [CrossRef]
- Huang, L.; Tepaamorndech, S. The SLC30 family of zinc transporters—A review of current understanding of their biological and pathophysiological roles. Mol. Aspects Med. 2013, 34, 548–560. [Google Scholar] [CrossRef]
- Grüngreiff, K.; Reinhold, D.; Wedemeyer, H. The role of zinc in liver cirrhosis. Ann. Hepatol. 2016, 15, 7–16. [Google Scholar] [CrossRef]
- Faa, G.; Nurchi, V.M.; Ravarino, A.; Fanni, D.; Nemolato, S.; Gerosa, C.; Eyken, P.V.; Geboes, K. Zinc in gastrointestinal and liver disease. Coord Chem. Rev. 2008, 252, 1257–1269. [Google Scholar] [CrossRef]
- Paterson, P.G.; Mas, A.; Sarkar, B.; Zlotkin, S.H. The influence of zinc-binding ligands in fetal circulation on zinc clearance across the in situ perfused guinea pig placenta. J. Nutr. 1991, 121, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Ford, D. Intestinal and placental zinc transport pathways. Proc. Nutr. Soc. 2004, 63, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Carey, L.C.; Coyle, P.; Philcox, J.C.; Rofe, A.M. Ethanol decreases zinc transfer to the fetus in normal but not metallothionein-null mice. Alcohol. Clin. Exp. Res. 2000, 24, 1236–1240. [Google Scholar] [CrossRef]
- Cousins, R.J.; Liuzzi, J.P.; Lichten, L.A. Mammalian zinc transport, trafficking, and signals. J. Biol. Chem. 2006, 281, 24085–24089. [Google Scholar] [CrossRef] [Green Version]
- Lewicka, I.; Kocyłowski, R.; Grzesiak, M.; Gaj, Z.; Oszukowski, P.; Suliburska, J. Selected trace elements concentrations in pregnancy and their possible role—Literature review. Ginekol. Pol. 2017, 88, 509–514. [Google Scholar] [CrossRef]
- Mocchegiani, E.; Romeo, J.; Malavolta, M.; Costarelli, L.; Giacconi, R.; Diaz, L.E.; Marcos, A. Zinc: Dietary intake and impact of supplementation on immune function in elderly. Age (Dordr) 2013, 35, 839–860. [Google Scholar] [CrossRef] [Green Version]
- Caulfield, L.E.; Zavaleta, N.; Shankar, A.H.; Merialdi, M. Potential contribution of maternal zinc supplementation during pregnancy to maternal and child survival. Am. J. Clin. Nutr. 1998, 68, 499S–508S. [Google Scholar] [CrossRef] [Green Version]
- Gapys, B.; Raszeja-Specht, A.; Bielarczyk, H. Role of zinc in physiological and pathological processes of the body. J. Lab. Diagn. Diagn Lab. 2014, 50, 45–52. [Google Scholar]
- Prasad, A.S. Discovery of human zinc deficiency: Its impact on human health and disease. Adv. Nutr. 2013, 4, 176–190. [Google Scholar] [CrossRef]
- Buamah, P.K.; Russell, M.; Bates, G.; Ward, A.M.; Skillen, A.W. Maternal zinc status: A determination of central nervous system malformation. BJOG An. Int. J. Obstet. Gynaecol. 1984, 91, 788–790. [Google Scholar] [CrossRef] [PubMed]
- Lehti, K.K. Stillbirth rates and folic acid and zinc status in low-socioeconomic pregnant women of brazilian amazon. Nutrition 1993, 9, 156–158. [Google Scholar] [PubMed]
- Scholl, T.O.; Hediger, M.L.; Schall, J.I.; Fischer, R.L.; Khoo, C.-S. Low zinc intake during pregnancy: Its association with preterm and very preterm delivery. Am. J. Epidemiol. 1993, 137, 1115–1124. [Google Scholar] [CrossRef]
- Siva, S.; Rubin, D.T.; Gulotta, G.; Wroblewski, K.; Pekow, J. Zinc deficiency is associated with poor clinical outcomes in patients with inflammatory bowel disease. Inflamm. Bowel Dis. 2017, 23, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osna, N.A.; Donohue, T.M., Jr.; Kharbanda, K.K. Alcoholic liver disease: Pathogenesis and current management. Alcohol Res. 2017, 38, 147. [Google Scholar]
- Mahajan, S.K. Zinc in kidney disease. J. Am. CollNutr. 1989, 8, 296–304. [Google Scholar] [CrossRef]
- Russell, R.M. Vitamin and mineral supplements in the management of liver disease. Med. Clin. North. Am. 1979, 63, 537–544. [Google Scholar] [CrossRef]
- Donangelo, C.M.; King, J.C. Maternal zinc intakes and homeostatic adjustments during pregnancy and lactation. Nutrients 2012, 4, 782–798. [Google Scholar] [CrossRef]
- Terrin, G.; Berni Canani, R.; Di Chiara, M.; Pietravalle, A.; Aleandri, V.; Conte, F.; Curtis, M.D. Zinc in early life: A key element in the fetus and preterm neonate. Nutrients 2015, 7, 10427–10446. [Google Scholar] [CrossRef] [Green Version]
- Karimi, A.; Bagheri, S.; Nematy, M.; Saeidi, M. Zinc deficiency in pregnancy and fetal—neonatal outcomes and impact of the supplements on pregnancy outcomes. MashhadUniv Med. Sci. 2012, 3, 77–83. [Google Scholar]
- Sultana, M.; Jahan, N.; Sultana, N.; Quraishi, S.B.; Chowdhury, T.R. Preterm delivery: Role of zinc. J. Bangladesh Soc. Physiol. 2010, 5, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Apgar, J. Effect of zinc deficiency on maintenance of pregnancy in the rat. J. Nutr. 1970, 100, 470–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKenzie, J.M.; Fosmire, G.J.; Sandstead, H.H. Zinc feficiency during the latter third of pregnancy: Effects on fetal rat brain, liver, and placenia. J. Nutr. 1975, 105, 1466–1475. [Google Scholar] [CrossRef] [PubMed]
- Golub, M.S.; Keen, C.L.; Gershwin, M.E.; Hendrickx, A.G. Developmental zinc deficiency and behavior. J. Nutr. 1995, 125, 2263S–2271S. [Google Scholar] [CrossRef] [PubMed]
- Nossier, S.A.; Naeim, N.E.; El-Sayed, N.A.; Abu Zeid, A.A. The effect of zinc supplementation on pregnancy outcomes: A double-blind, randomised controlled trial, Egypt. Br. J. Nutr. 2015, 114, 274–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prawirohartono, E.P.; Nyström, L.; Nurdiati, D.S.; Hakimi, M.; Lind, T. The impact of prenatal vitamin A and zinc supplementation on birth size and neonatal survival—A double-blind, randomized controlled trial in a rural area of Ind.onesia. Int. J. Vitam Nutr. Res. 2013, 83, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hu, Y.-F.; Hao, J.-H.; Chen, Y.-H.; Su, P.-Y.; Wang, Y.; Yu, Z.; Fu, L.; Xu, Y.Y.; Zhang, C.; et al. Maternal zinc deficiency during pregnancy elevates the risks of fetal growth restriction: A population-based birth cohort study. Sci. Rep. 2015, 5, 11262. [Google Scholar] [CrossRef] [Green Version]
- Merialdi, M.; Caulfield, L.E.; Zavaleta, N.; Figueroa, A.; Dominici, F.; DiPietro, J.A. Randomized controlled trial of prenatal zinc supplementation and the development of fetal heart rate. Am. J. Obstet. Gynecol. 2004, 190, 1106–1112. [Google Scholar] [CrossRef]
- Merialdi, M.; Caulfield, L.E.; Zavaleta, N.; Figueroa, A.; DiPietro, J.A. Adding zinc to prenatal iron and folate tablets improves fetal neurobehavioral development. Am. J. Obstet. Gynecol. 1999, 180, 483–490. [Google Scholar] [CrossRef]
- Merialdi, M.; Caulfield, L.E.; Zavaleta, N.; Figueroa, A.; Costigan, K.A.; Dominici, F.; Dipietro, J.A. Randomized controlled trial of prenatal zinc supplementation and fetal bone growth. Am. J. Clin. Nutr. 2004, 79, 826–830. [Google Scholar] [CrossRef] [Green Version]
- Wilson, R.L.; Leemaqz, S.Y.; Goh, Z.; McAninch, D.; Jankovic-Karasoulos, T.; Leghi, G.E.; Phillips, J.A.; Colafella, K.M.; Tran, C.; O’Leary, S.; et al. Zinc is a critical regulator of placental morphogenesis and maternal hemodynamics during pregnancy in mice. Sci. Rep. 2017, 7, 15137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, L.P.; Caton, J.S.; Redmer, D.A.; Grazul-Bilska, A.T.; Vonnahme, K.A.; Borowicz, P.P.; Luther, J.S.; Wallace, J.M.; Wu, G.; Spencer, T.E. Evidence for altered placental blood flow and vascularity in compromised pregnancies. J. Physiol. 2006, 572, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Ota, E.; Mori, R.; Middleton, P.; Tobe-Gai, R.; Mahomed, K.; Miyazaki, C.; Bhutta, Z.A. Zinc supplementation for improving pregnancy and infant outcome. Cochrane Database Syst. Rev. 2015, 2015, CD000230. [Google Scholar] [CrossRef] [PubMed]
- Karamali, M.; Heidarzadeh, Z.; Seifati, S.-M.; Samimi, M.; Tabassi, Z.; Talaee, N.; Bahardoost, H.; Asemi, Z. Zinc supplementation and the effects on pregnancy outcomes in gestational diabetes: A randomized, double-blind, placebo-controlled trial. Exp. Clin. Endocrinol. Diabetes 2015, 124, 28–33. [Google Scholar] [CrossRef]
- Zahiri sorouri, Z.; Sadeghi, H.; Pourmarzi, D. The effect of zinc supplementation on pregnancy outcome: A randomized controlled trial. J. Matern. Fetal Neonatal Med. 2016, 29, 2194–2198. [Google Scholar] [CrossRef]
- Chaffee, B.W.; King, J.C. Effect of zinc supplementation on pregnancy and infant outcomes: A systematic review. Paediatr. Perinat. Epidemiol. 2012, 26, 118–137. [Google Scholar] [CrossRef] [Green Version]
- Mori, R.; Ota, E.; Middleton, P.; Tobe-Gai, R.; Mahomed, K.; Bhutta, Z.A. Zinc supplementation for improving pregnancy and infant outcome. In Cochrane Database of Systematic Reviews; Mori, R., Ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2012. [Google Scholar]
- King, J.C. Determinants of maternal zinc status during pregnancy. Am. J. Clin. Nutr. 2000, 71, 1334S–1343S. [Google Scholar] [CrossRef] [Green Version]
- Goodnight, W.; Newman, R. Society of maternal-fetal medicine. Optimal nutrition for improved twin pregnancy outcome. Obstet. Gynecol. 2009, 114, 1121–1134. [Google Scholar] [CrossRef] [Green Version]
- Mahomed, K.; Bhutta, Z.A.; Middleton, P. Zinc supplementation for improving pregnancy and infant outcome. In Cochrane Database of Systematic Reviews; Mahomed, K., Ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2007. [Google Scholar]
- Campbell, D.M.; MacGillivray, I.; Tuttle, S. Maternal nutrition in twin pregnancy. Acta Genet. Med. Gemellol (Roma) 1982, 31, 221–227. [Google Scholar] [CrossRef]
- Fosmire, G.J. Zinc toxicity. Am. J. Clin. Nutr. 1990, 51, 225–227. [Google Scholar] [CrossRef]
- Plum, L.M.; Rink, L.; Haase, H. The essential toxin: Impact of zinc on human health. Int. J. Environ. Res. Public Health 2010, 7, 1342–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Environmental Health Criteria 221. Zinc; FIRST Draft Prepared; World Health Organization: Geneva, Switzerland, 2001.
- Scheers, N. Regulatory effects of Cu, Zn, and Ca on Fe absorption: The intricate play between nutrient transporters. Nutrients 2013, 5, 957–970. [Google Scholar] [CrossRef] [PubMed]
- Ha, J.-H.; Doguer, C.; Wang, X.; Flores, S.R.; Collins, J.F. High-iron consumption impairs growth and causes copper-deficiency anemia in weanling Sprague-Dawley rats. PLoS ONE 2016, 11, e0161033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Huang, G.; Su, T.; Gao, H.; Attieh, Z.K.; McKie, A.T.; Anderson, G.A.; Vulpe, C.D. Decreased hephaestin activity in the intestine of copper-deficient mice causes systemic iron deficiency. J. Nutr. 2006, 136, 1236–1241. [Google Scholar] [CrossRef] [Green Version]
- Reeves, P.G.; DeMars, L.C.S.; Johnson, W.T.; Lukaski, H.C. Dietary copper deficiency reduces iron absorption and duodenal enterocyte hephaestin protein in male and female Rats. J. Nutr. 2005, 135, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Muckenthaler, M.U.; Rivella, S.; Hentze, M.W.; Galy, B. A red carpet for iron metabolism. Cell 2017, 168, 344–361. [Google Scholar] [CrossRef] [Green Version]
- Toxqui, L.; Vaquero, M.P. Chronic iron deficiency as an emerging risk factor for osteoporosis: A hypothesis. Nutrients 2015, 7, 2324–2344. [Google Scholar] [CrossRef] [Green Version]
- Ha, J.-H.; Doguer, C.; Collins, J.F. Consumption of a high-iron diet disrupts homeostatic regulation of intestinal copper absorption in adolescent mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, G353. [Google Scholar] [CrossRef] [Green Version]
- Collins, J.F.; Prohaska, J.R.; Knutson, M.D. Metabolic crossroads of iron and copper. Nutr. Rev. 2010, 68, 133–147. [Google Scholar] [CrossRef] [Green Version]
- Chan, W.Y.; Rennert, O.M. The role of copper in iron metabolism. Ann. Clin. Lab. Sci. 1980, 10, 338–344. [Google Scholar]
- Gulec, S.; Collins, J.F. Molecular mediators governing iron-copper interactions. Annu. Rev. Nutr. 2014, 34, 95–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, E.D. The iron-copper connection: The link to ceruloplasmin grows stronger. Nutr. Rev. 1995, 53, 170–173. [Google Scholar] [CrossRef] [PubMed]
- Ha, J.-H.; Doguer, C.; Flores, S.R.L.; Wang, T.; Collins, J.F. Progressive increases in dietary iron are associated with the emergence of pathologic disturbances of copper homeostasis in growing rats. J. Nutr. 2018, 148, 373–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klevay, L.M. IHD from copper deficiency: A unified theory. Nutr. Res. Rev. 2016, 29, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Kedzierska, K.; Bober, J.; Ciechanowski, K.; Golembiewska, E.; Kwiatkowska, E.; Nocen, I.; Dołegowska, B.; Dutkiewicz, G.; Chlubek, D. Copper modifies the activity of sodium-transporting systems in erythrocyte membrane in patients with essential hypertension. Biol. Trace Elem. Res. 2005, 107, 021–032. [Google Scholar] [CrossRef]
- Broderius, M.; Mostad, E.; Wendroth, K.; Prohaska, J.R. Levels of plasma ceruloplasmin protein are markedly lower following dietary copper deficiency in rodents. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2010, 151, 473. [Google Scholar] [CrossRef] [Green Version]
- Fields, M.; Lewis, C.G. Hepatic iron overload may contribute to hypertriglyceridemia and hypercholesterolemia in copper-deficient rats. Metabolism 1997, 46, 377–381. [Google Scholar] [CrossRef]
- Zhao, M.; Matter, K.; Laissue, J.A.; Zimmermann, A. Copper/zinc and manganese superoxide dismutase immunoreactivity in hepatic iron overload diseases. Histol Histopathol. 1995, 10, 925–935. [Google Scholar]
- Patel, B.N.; David, S. A novel glycosylphosphatidylinositol-anchored form of ceruloplasmin is expressed by mammalian astrocytes. J. Biol. Chem. 1997, 272, 20185–20190. [Google Scholar] [CrossRef]
- Sebio, R.M.; Ferrarotti, N.; Lairion, F.; Magriñá, C.S.; Fuda, J.; Torti, H.; Boveris, A.; Repetto, M.G. Brain oxidative stress in rat with chronic iron or copper overload. J. Inorg. Biochem. 2019, 199, 110799. [Google Scholar] [CrossRef]
- Lan, A.P.; Chen, J.; Chai, Z.F.; Hu, Y. The neurotoxicity of iron, copper and cobalt in Parkinson’s disease through ROS-mediated mechanisms. BioMetals 2016, 29, 665–678. [Google Scholar] [CrossRef] [PubMed]
- Troost, F.J.; Brummer, R.-J.M.; Dainty, J.R.; Hoogewerff, J.A.; Bull, V.J.; Saris, W.H. Iron supplements inhibit zinc but not copper absorption in vivo in ileostomy subjects. Am. J. Clin. Nutr. 2003, 78, 1018–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christian, P.; Khatry, S.K.; LeClerq, S.C.; Ram Shrestha, S.; Kimbrough-Pradhan, E.; West, K.P. Iron and zinc interactions among pregnant Nepali women. Nutr. Res. 2001, 21, 141–148. [Google Scholar] [CrossRef]
- Saaka, M. Combined iron and zinc supplementation improves haematologic status of pregnant women in Upper West Region of Ghana. Ghana Med. J. 2012, 46, 225–233. [Google Scholar]
- Saaka, M.; Oosthuizen, J.; Beatty, S. Effect of prenatal zinc supplementation on birthweight. J. Health Popul. Nutr. 2009, 27, 619–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, L.J.; Dainty, J.R.; Hollands, W.J.; Bull, V.J.; Hoogewerff, J.A.; Foxall, R.J.; McAnena, L.; Strain, J.J.; Fairweather-Tait, S. Effect of high-dose iron supplements on fractional zinc absorption and status in pregnant women. Am. J. Clin. Nutr. 2007, 85, 131–136. [Google Scholar] [CrossRef]
- Peters, A.J.; Keen, C.L.; Lönnerdal, B.; Hurley, L.S. Zinc-vitamin a interaction in pregnant and fetal rats: Supplemental vitamin a does not prevent zinc-deficiency-induced teratogenesis. J. Nutr. 1986, 116, 1765–1771. [Google Scholar] [CrossRef]
- O’Brien, K.O.; Zavaleta, N.; Caulfield, L.E.; Wen, J.; Abrams, S.A. Prenatal iron supplements impair zinc absorption in pregnant Peruvian women. J. Nutr. 2000, 130, 2251–2255. [Google Scholar] [CrossRef] [Green Version]
- Whittaker, P. Iron and zinc interactions in humans. Am. J. Clin. Nutr. 1998, 68, 442S–446S. [Google Scholar] [CrossRef]
- Bodiga, S.; Krishnapillai, M.N. Concurrent repletion of iron and zinc reduces intestinal oxidative damage in iron- and zinc-deficient rats. World J. Gastroenterol. 2007, 13, 5707–5717. [Google Scholar] [CrossRef] [Green Version]
- Garrick, M.D.; Singleton, S.T.; Vargas, F.; Kuo, H.C.; Zhao, L.; Knöpfel, M.; Davidson, T.; Costa, M.; Paradkar, P.; Roth, J.A.; et al. DMT-1, Which metals does it transport? Biol. Res. 2006, 39, 79–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kordas, K.; Stoltzfus, R.J. New evidence of iron and zinc interplay at the enterocyte and neural tissues. J. Nutr. 2004, 134, 1295–1298. [Google Scholar] [CrossRef] [PubMed]
- Bowers, K.; Srai, S.K.S. The trafficking of metal ion transporters of the Zrt- and Irt-like protein family. Traffic 2018, 19, 813–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomons, N.W.; Jacob, R.A. Studies on the bioavailability of zinc in humans: Effects of heme and nonheme iron on the absorption of zinc. Am. J. Clin. Nutr. 1981, 34, 475–482. [Google Scholar] [CrossRef]
- Solomons, N.W.; Pineda, O.; Viteri, F.; Sandstead, H.H. Studies on the bioavailability of zinc in humans: Mechanism of the intestinal interaction of nonheme iron and zinc. J. Nutr. 1983, 113, 337–349. [Google Scholar] [CrossRef]
- Khoushabi, F.; Shadan, M.R.; Miri, A.; Sharifi-Rad, J. Determination of maternal serum zinc, iron, calcium and magnesium during pregnancy in pregnant women and umbilical cord blood and their association with outcome of pregnancy. Mater. Sociomed 2016, 28, 104–107. [Google Scholar] [CrossRef] [Green Version]
- Campbell-Brown, M.; Ward, R.J.; Haines, A.P.; North, W.R.; Abraham, R.; McFadyen, I.R.; Turnlund, J.R.; King, J.C. Zinc and copper in Asian pregnancies-is there evidence for a nutritional deficiency? Br. J. Obstet. Gynaecol. 1985, 92, 875–885. [Google Scholar] [CrossRef]
- Breskin, M.W.; Worthington-Roberts, B.S.; Knopp, R.H.; Brown, Z.; Plovie, B.; Mottet, N.K.; Mills, J.L. First trimester serum zinc concentrations in human pregnancy. Am. J. Clin. Nutr. 1983, 38, 943–953. [Google Scholar] [CrossRef] [Green Version]
- Ziaei, S.; Janghorban, R.; Shariatdoust, S.; Faghihzadeh, S. The effects of iron supplementation on serum copper and zinc levels in pregnant women with high-normal hemoglobin. Int. J. Gynecol. Obstet. 2008, 100, 133–135. [Google Scholar] [CrossRef]
- Andersen, H.S.; Gambling, L.; Holtrop, G.; McArdle, H.J. Effect of dietary copper deficiency on iron metabolism in the pregnant rat. Br. J. Nutr. 2007, 97, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Gambling, L.; Danzeisen, R.; Fosset, C.; Andersen, H.S.; Dunford, S.; Srai, S.K.S.; McArdle, H.J. Iron and copper interactions in development and the effect on pregnancy outcome. J. Nutr. 2003, 133, 1554S–1556S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shidfar, F.; Ameri, A.; Keshavarz, S.; Djalali, M. Effect of iron supplements on serum copper status in pregnant women in Islamshahr. Iran. J. Endocrinol. Metab. 2003, 5, 299–305. [Google Scholar]
- Donangelo, C.M.; Woodhouse, L.R.; King, S.M.; Viteri, F.E.; King, J.C. Supplemental zinc lowers measures of iron status in young women with low iron reserves. J. Nutr. 2002, 132, 1860–1864. [Google Scholar] [CrossRef]
- de Brito, N.J.N.; Rocha, É.D.; de Araújo Silva, A.; Costa, J.B.S.; França, M.C.; das Graças Almeida, M.; Brandão-Neto, J. Oral zinc supplementation decreases the serum iron concentration in healthy schoolchildren: A pilot study. Nutrients 2014, 6, 3460–3473. [Google Scholar] [CrossRef] [PubMed]
- Antunes, M.F.R.; Leite, L.D.; Rocha, É.D.M.; Brito, N.J.N.; França, M.C.; Silva, C.A.B.; Almeida, M.G.; de Rezende, A.A.; Marchini, J.S.; Brandão-Neto, J. Competitive interaction of zinc and iron after venous and oral zinc administration in eutrophic children. Trace Elem. Electrolytes. 2010, 27, 185–191. [Google Scholar] [CrossRef]
- Wieringa, F.T.; Berger, J.; Dijkhuizen, M.A.; Hidayat, A.; Ninh, N.X.; Utomo, B.; Wasantwisut, E.; Winichagoon, P. Combined iron and zinc supplementation in infants improved iron and zinc status, but interactions reduced efficacy in a multicountry trial in southeast Asia. J. Nutr. 2007, 137, 466–471. [Google Scholar]
- Holmes, J.B.; Kroeun, H.; Houghton, L.A.; Gibson, R.S.; Harding, K.B.; De-Regil, L.M.; Kraemer, K.; Barr, S.I.; Karakochuk, C.D. Including 60 mg elemental iron in a multiple micronutrient supplement blunts the increase in serum zinc after 12 weeks of daily supplementation in predominantly anemic, nonpregnant Cambodian women of reproductive age. J. Nutr. 2019, 149, 1503–1510. [Google Scholar] [CrossRef]
- Nguyen, P.; Grajeda, R.; Melgar, P.; Marcinkevage, J.; Flores, R.; Ramakrishnan, U.; Martorell, R. Effect of zinc on efficacy of iron supplementation in improving iron and zinc status in women. J. Nutr. Metab. 2012, 2012, 216179. [Google Scholar] [CrossRef] [Green Version]
- Davidsson, L.; Almgren, A.; Sandström, B.; Hurrell, R.F. Zinc absorption in adult humans: The effect of iron fortification. Br. J. Nutr. 1995, 74, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Shaw, C.F.; Savas, M.M.; Petering, D.H. Ligand substitution and sulfhydryl reactivity of metallothionein. Methods Enzymol. 1991, 205, 401–414. [Google Scholar]
- Merza, H.; Sood, N.; Sood, R. Idiopathic hyperzincemia with associated copper deficiency anemia: A diagnostic dilemma. Clin. Case Rep. 2015, 3, 819–822. [Google Scholar] [CrossRef] [PubMed]
- Sutton, L.; Vusirikala, M.; Chen, W. Hematogone hyperplasia in copper deficiency. Am. J. Clin. Pathol. 2009, 132, 191–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, R.; Hawthorne, B.; Durai, D.; McDowell, I. Zinc in denture adhesive: A rare cause of copper deficiency in a patient on home parenteral nutrition. BMJ Case Rep. 2015, 2015, bcr2015211390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deur, C.J.; Stone, M.J.; Frenkel, E.P. Trace metals in hematopoiesis. Am. J. Hematol. 1981, 11, 309–331. [Google Scholar] [CrossRef]
- Percival, S.S. Neutropenia caused by copper deficiency: Possible mechanisms of action. Nutr. Rev. 1995, 53, 59–66. [Google Scholar] [CrossRef]
- Paik, H.Y.; Joung, H.; Lee, J.Y.; Lee, H.K.; King, J.C.; Keen, C.L. Serum extracellular superoxide dismutase activity as an indicator of zinc status in humans. Biol. Trace Elem. Res. 1999, 69, 45–57. [Google Scholar] [CrossRef]
- Davis, C.D.; Milne, D.B.; Nielsen, F.H. Changes in dietary zinc and copper affect zinc-status indicators of postmenopausal women, notably, extracellular superoxide dismutase and amyloid precursor proteins. Am. J. Clin. Nutr. 2000, 71, 781–788. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, A.; Bhatt, M. Advances in treatment of Wilson disease. Tremor Other Hyperkinet. Mov. 2018, 8, 525. [Google Scholar] [CrossRef]
- Kathawala, M.; Hirschfield, G.M. Insights into the management of Wilson’s disease. Therap. Adv. Gastroenterol. 2017, 10, 889–905. [Google Scholar] [CrossRef]
- Wiggelinkhuizen, M.; Tilanus, M.E.C.; Bollen, C.W.; Houwen, R.H.J. Systematic review: Clinical efficacy of chelator agents and zinc in the initial treatment of Wilson disease. Aliment. Pharmacol. Ther. 2009, 29, 947–958. [Google Scholar] [CrossRef]
- Walshe, J.M.; Munro, N.A.R. Zinc-induced deterioration in Wilson’s disease aborted by treatment with penicillamine, dimercaprol, and a novel zero copper diet. Arch. Neurol. 1995, 52, 10–11. [Google Scholar] [CrossRef]
- Malik, A.; Khawaja, A.; Sheikh, L. Wilson’s disease in pregnancy: Case series and review of literature. BMC Res. Notes. 2013, 6, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Liu, Z.; Guo, J.; Wan, C.; Zhang, T.; Cui, H.; Yang, F.; Gao, X. Influence of dietary zinc and copper on apparent mineral retention and serum biochemical indicators in young male mink (Mustela vison). Biol. Trace Elem. Res. 2015, 165, 59–66. [Google Scholar] [CrossRef]
- Eckert, G.E.; Greene, L.W.; Carstens, G.E.; Ramsey, W.S. Copper status of ewes fed increasing amounts of copper from copper sulfate or copper proteinate. J. Anim Sci. 1999, 77, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Hemken, R.W.; Jackson, J.A.; Trammell, D.S. Utilization of copper in copper proteinate, copper lysine, and cupric sulfate using the rat as an experimental model. J. Anim Sci. 1996, 74, 1657–1667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zetzsche, A.; Schunter, N.; Zentek, J.; Pieper, R. Accumulation of copper in the kidney of pigs fed high dietary zinc is due to metallothionein expression with minor effects on genes involved in copper metabolism. J. Trace Elem. Med. Biol. 2016, 35, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Baecker, T.; Mangus, K.; Pfaender, S.; Chha1qbra, R.; Boeckers, T.M.; Grabrucker, A.M. Loss of COMMD1 and copper overload disrupt zinc homeostasis and influence an autism-associated pathway at glutamatergic synapses. BioMetals 2014, 27, 715–730. [Google Scholar] [CrossRef] [Green Version]
- Reinstein, N.H.; Lönnerdal, B.; Keen, C.L.; Hurley, L.S. Zinc-copper interactions in the pregnant rat: Fetal outcome and maternal and fetal zinc, copper and iron. J. Nutr. 1984, 114, 1266–1279. [Google Scholar] [CrossRef]
- Kinnamon, K.E. Some independent and combined effects of copper, molybdenum, and zinc on the placental transfer of zinc-65 in the Rat. J. Nutr. 1963, 81, 312–320. [Google Scholar] [CrossRef]
- Garg, H.K.; Singhal, K.C.; Arshad, Z. Effect of oral zinc supplementation on copper and haemoglobin levels in pregnant women. Indian J. Physiol. Pharmacol. 1994, 38, 272–276. [Google Scholar]


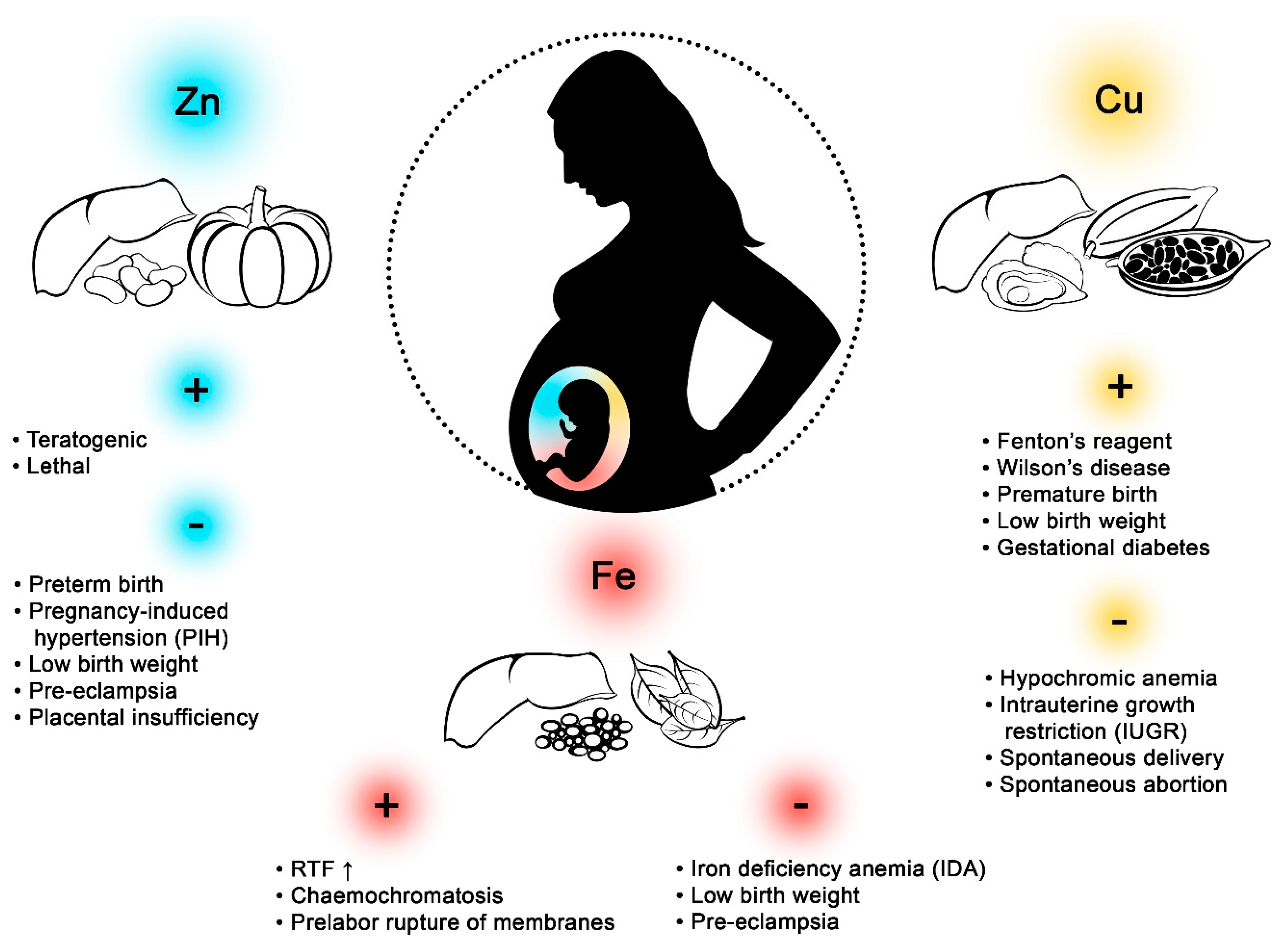{kind=link}
| Research Area | n | Mean Age(Years) | Concentration of | References | ||
|---|---|---|---|---|---|---|
| Fe | Cu | Zn | ||||
| Europe | ||||||
| Szczecin, Poland | 170 | 29 | P: 640.726 mg/kg dw | P: 6.013 mg/kg dw | P: 66.904 g/kg dw | [2] |
| FM: 640.726 mg/kg dw | FM: 8.906 mg/kg dw | FM: 62.788 g/kg dw | ||||
| C: 567.285 mg/kg dw | C: 4.320 mg/kg dw | C: 54.653 g/kg dw | ||||
| Poznan, Poland | 64 | 28.1 | MS: 1.08 μg/mL | MS: 0.63 μg/mL | MS: 1.91 μg/mL | [3] |
| CS (vein): 1.96 μg/mL | CS (vein): 0.65 μg/mL | CS (vein): 0.36 μg/mL | ||||
| CS (artery): 1.63 μg/mL | CS (artery): 0.65 μg/mL | CS (artery): 0.36 μg/mL | ||||
| Oleśnica, Poland | 64 | 27 | - | MP: 1.93 mg/L | MP: 0.58 mg/L | [4] |
| CP: 0.49 mg/L | CP: 0.82 mg/L | |||||
| Sverdlovsk region (non-industrial areas) andYekaterinburg (industrial city), Russian | Pregnant women from non-industrial areas (29) | Age range: 17–42 | - | MS: 5.44 mkg/mL P: 6.1 mkg/mL | - | [5] |
| Pregnant women from industrial city (127) | - | MS: 4.73 mkg/mL P: 13.34 mkg/mL | - | |||
| Moscow, Russian | (150) control | 33.1 | MS: 1.34 µg/L | MS: 1.15 µg/L | - | [6] |
| Pregnancy (169) | 33.4 | MS: 1.27 µg/L | MS:1.60 µg/L | - | ||
| Miscarriage (75) | 34.8 | MS: 1.43 µg/L | MS: 1.12 µg/L | - | ||
| Infertility (91) | 35.5 | MS: 1.29 µg/L | MS:1.04 µg/L | - | ||
| Barcelona, Spain | Appropriate for gestational age (96) | 32 | - | - | MS: 1181 µg /dL ww FS: 1518 µg /dL ww P: 8.4 µg /dL ww | [7] |
| Intrauterine growth restriction (49) | 32 | - | - | MS: 935 µg/dL ww FS: 935 µg/dL ww P: 8.5 µg/dL ww | ||
| Small for gestational age (33) | 30 | - | - | MS: 984 µg/dL ww FS: 1134 µg/dL ww P: 8.9 µg/dL ww | ||
| Africa | ||||||
| Mid-Western Region, Nigeria | 22 | - | P: 84.3 µg/gm dw | P: 6.3 µg/μm dw | P: 66.6 µg/μm dw | [8] |
| North America | ||||||
| New Hampshire, USA | 1159 | 31.2 | - | - | P: 10.26 µg/g ww | [9] |
| Las Vegas, Nevada, USA | 28 | 29.9 | P: 1185.18 µg/L dw | P: 7.81 µg/L dw | P: 63.59 µg/L dw | [10] |
| New York, USA | Women carrying multiples (101) | 30 | P: 26.63 µg/g dw | - | P: 5.9 µg/g dw | [11] |
| Women carrying singletons (132) | 17.4 | P: 17.99 µg/g dw | - | P: 2.6 µg/g dw | ||
| Chattanooga, USA | 374 | - | P: 503,200 µg/kg dw | P: 3889 µg/kg dw | P: 55,120 µg/kg dw | [12] |
| Pune, India | Normal pregnancies (47) | Age range: 19–35 | MB: 120.4 µg/dL | MB: 1.44 µg/dL | MB: 57.5 µg/dL | [13] |
| CB: 153.4 µg/dL | CB: 0.26 µg/dL | CB: 90.8 µg/dL | ||||
| Preeclamptic pregnancies (14) | MB: 96.3 µg/dL | MB: 1.58 µg/dL | MB: 49.2 µg/dL | |||
| CB: 118.6 µg/dL | CB: 0.81 µg/dL | CB:79.9 µg/dL | ||||
| Delhi, India | Females delivered full term babies (gestational age > 37 weeks) (50) | 25.54 | P: 58.94 µg/dL dw | P: 0.255 µg/dL dw | P: 18.28 µg/dL dw | [14] |
| Females delivered pre-term babies (gestational age <37 weeks)(30) | 24.63 | P: 50.60 µg/dL dw | P: 0.220 µg/dL dw | P: 17.26 µg/dL dw | ||
| Jaipur, India | Pregnant women (80) | 29.8 | P: 72.7 µg/dL | P: 187.3 µg/dL | P: 70.5 µg/dL | [15] |
| Non-pregnant women (20) | 20 | P: 106.9 µg/dL | P: 127.7 µg/dL | P: 100.9 µg/dL | ||
| Ahmedabad, India | Women with spontaneous abortion(159) | 24.85 | - | MS: 1.59 mg/L | MS: 1.43 mg/L | [16] |
| Woman without spontaneous abortion (118) | 23.65 | - | MS: 1.81 mg/L | MS: 1.46 mg/L | ||
| Hyderabad, India | Pregnant women from rural area (30) | 21.1 | MS: 201.6μg/dL | MS: 166.6μg/dL | MS: 205.1μg/dL | [17] |
| CS: 279.8 μg/dL | CS: 92.03μg/dL | CS: 128.2 μg/dL | ||||
| P: 1159.5 μg/g | P: 78.4 μg/g | P: 49.6 μg/g | ||||
| Pregnant women from urban area (30) | 22.2 | MS: 128.6 μg/dL | MS: 150.7μg/dL | MS: 245.6μg/dL | ||
| CS: 200.3 μg/dL | CS:83.5μg/dL | CS: 122.1μg/dL | ||||
| P: 1458.2 μg/g | P: 61.4μg/g | P: 51.5 μg/g | ||||
| Jakarta, Indonesia | Pregnant womens ≥37 weeks of gestational age for the term group (25) | 27.68 | P: 252.16 µg/g dw | P: 2.96 µg/g dw | P: 58.34 µg/g dw | [18] |
| CB: 212.00 µg/dL dw | CB: 32.20 µg/dL dw | CB: 293.80 µg/dL dw | ||||
| MS: 77 µg/dL dw | MS: 222.65 µg/dL dw | MS: 45.16 µg/dL dw | ||||
| Pregnant womens preterm birth in 26–36 weeks of gestational age (26) | 24.0 | P: 78.45 µg/g dw | P: 1.62 µg/g dw | P: 28.41 µg/g dw | ||
| CB: 236.50 µg/dL dw | CB: 20.60 µg/dL dw | CB: 321.43 µg/dL dw | ||||
| MS: 71.50 µg/dL dw | MS: 215.35 µg/dL dw | MS: 40.26 µg/dL dw | ||||
| Fukuoka, Japan | 48 | 29.3 | - | P: 3910 ng/g dw | P: 48,100 ng/g dw | [19] |
| C: 2960 ng/g dw | C: 35,700 ng/g dw | |||||
| Shanghai, China | 1568 | 26.4 | MS: 8.1 mmol/L | MS: 23.43 µmol/L | MS: 87.32 µmol/L | [20] |
| Amman, Jordan | 92 | 27 | CB: 116 µg/dL | CB: 49 µg/dL | CB: 114 µg/dL | [21] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grzeszczak, K.; Kwiatkowski, S.; Kosik-Bogacka, D. The Role of Fe, Zn, and Cu in Pregnancy. Biomolecules 2020, 10, 1176. https://doi.org/10.3390/biom10081176
Grzeszczak K, Kwiatkowski S, Kosik-Bogacka D. The Role of Fe, Zn, and Cu in Pregnancy. Biomolecules. 2020; 10(8):1176. https://doi.org/10.3390/biom10081176
Chicago/Turabian StyleGrzeszczak, Konrad, Sebastian Kwiatkowski, and Danuta Kosik-Bogacka. 2020. "The Role of Fe, Zn, and Cu in Pregnancy" Biomolecules 10, no. 8: 1176. https://doi.org/10.3390/biom10081176