Immunoglobulin E-Binding Pattern of Canadian Peanut Allergic Children and Cross-Reactivity with Almond, Hazelnut and Pistachio


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Human Sera from Allergic and Nonatopic Subjects
2.3. Preparation of Defatted Peanut and Tree Nuts Flours
2.4. Preparation of Crude Protein Extracts from Peanut and Tree Nuts
2.5. Protein Quantitation
2.6. Protein Electrophoresis
2.7. Serum Immunoglobulin E (IgE) Indirect ELISA
2.8. Serum IgE Indirect Western Immunoblotting
2.9. Reciprocal IgE Inhibition ELISA and Immunoblotting
2.10. Statistical Analysis
3. Results and Discussion
3.1. Clinical Profiles of Peanut-Allergic Patients and their Sensitization Profiles to Tree Nuts by IgE ImmunoCAP Testing
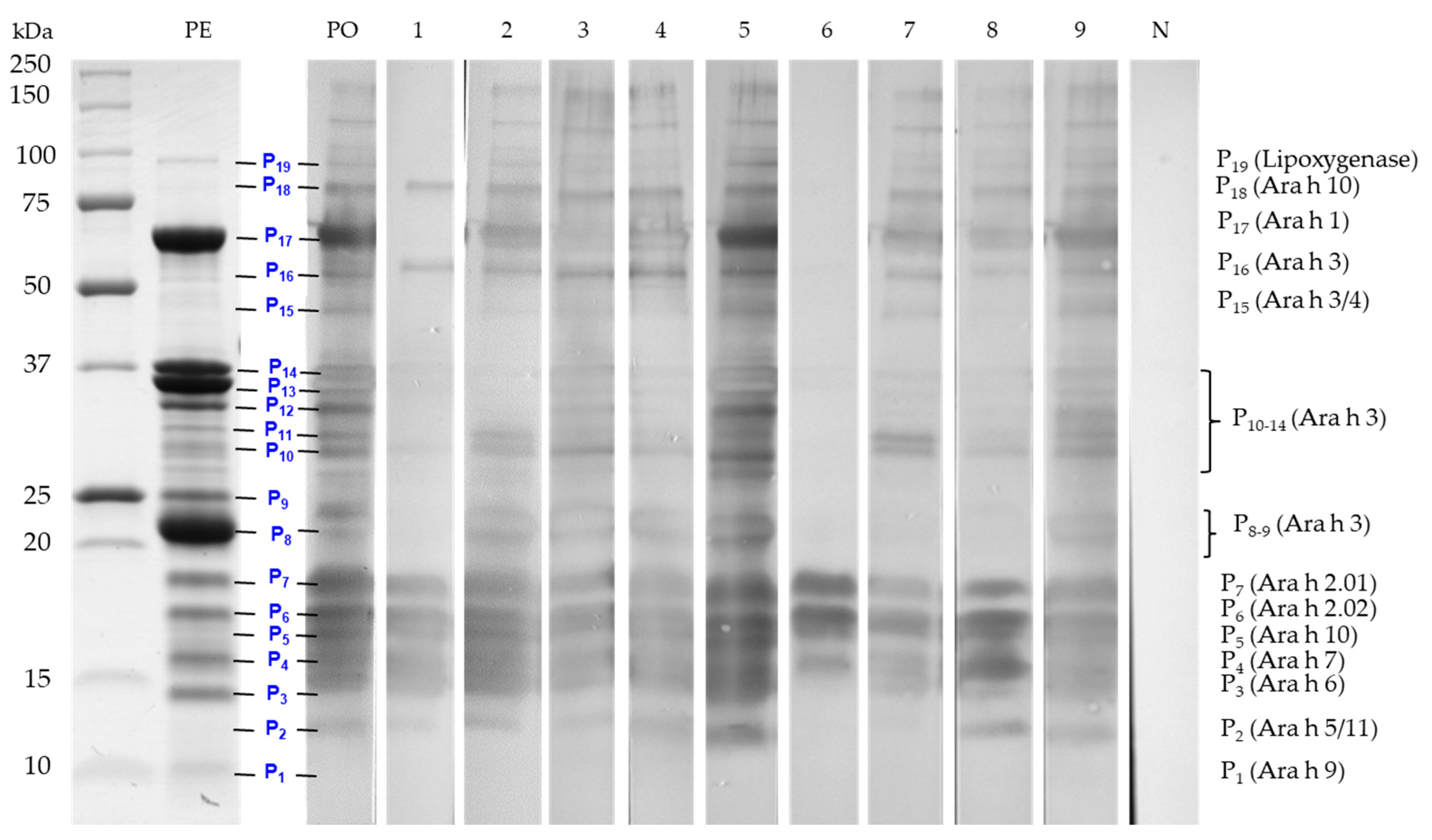
3.2. Electrophoretic Protein Profiles of Prepared Peanut and Selected Tree Nut Extracts and Correspondence to Known Allergens
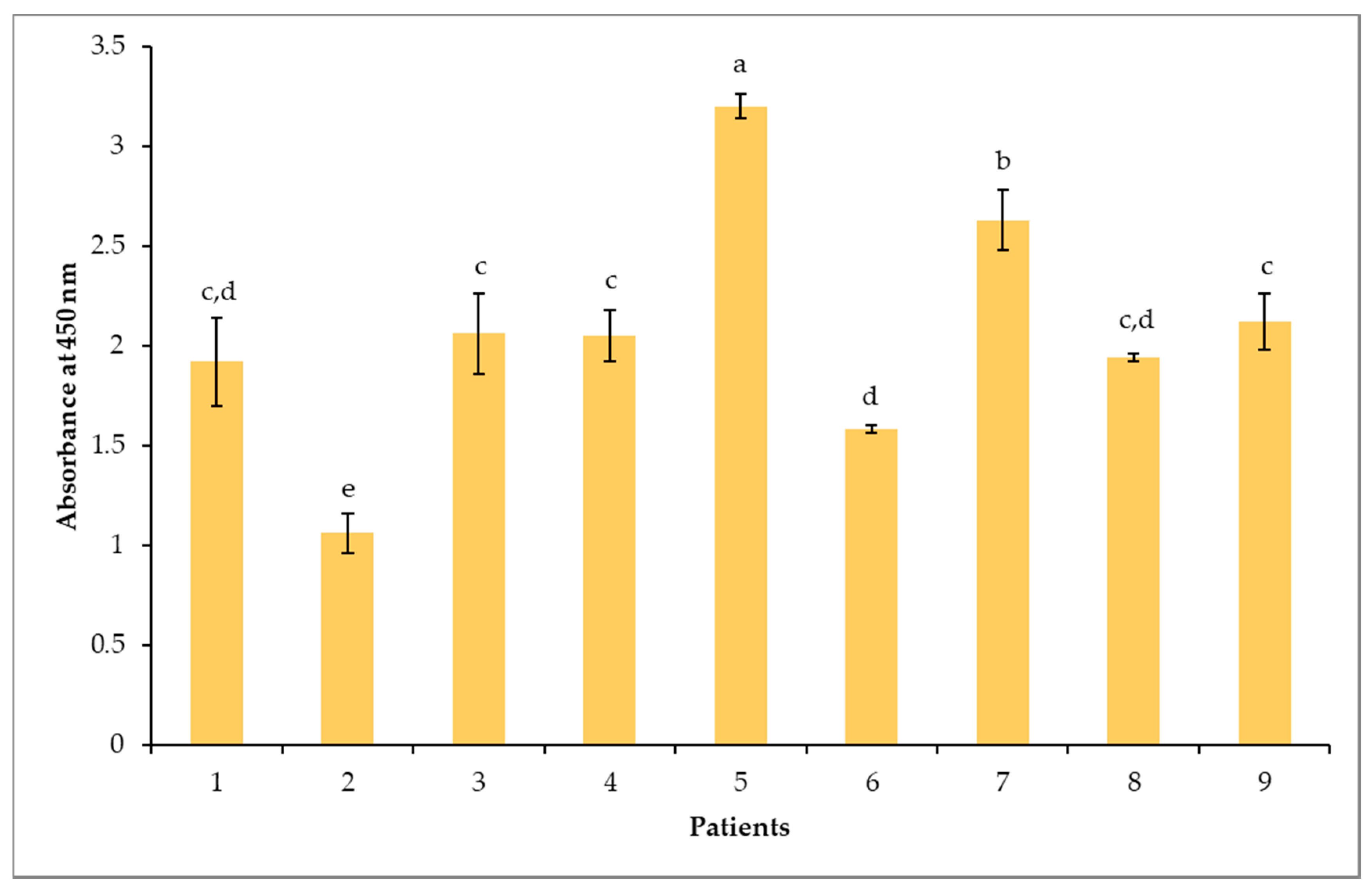
3.3. Indirect ELISA and Immunolotting for IgE Responses to Peanut Proteins
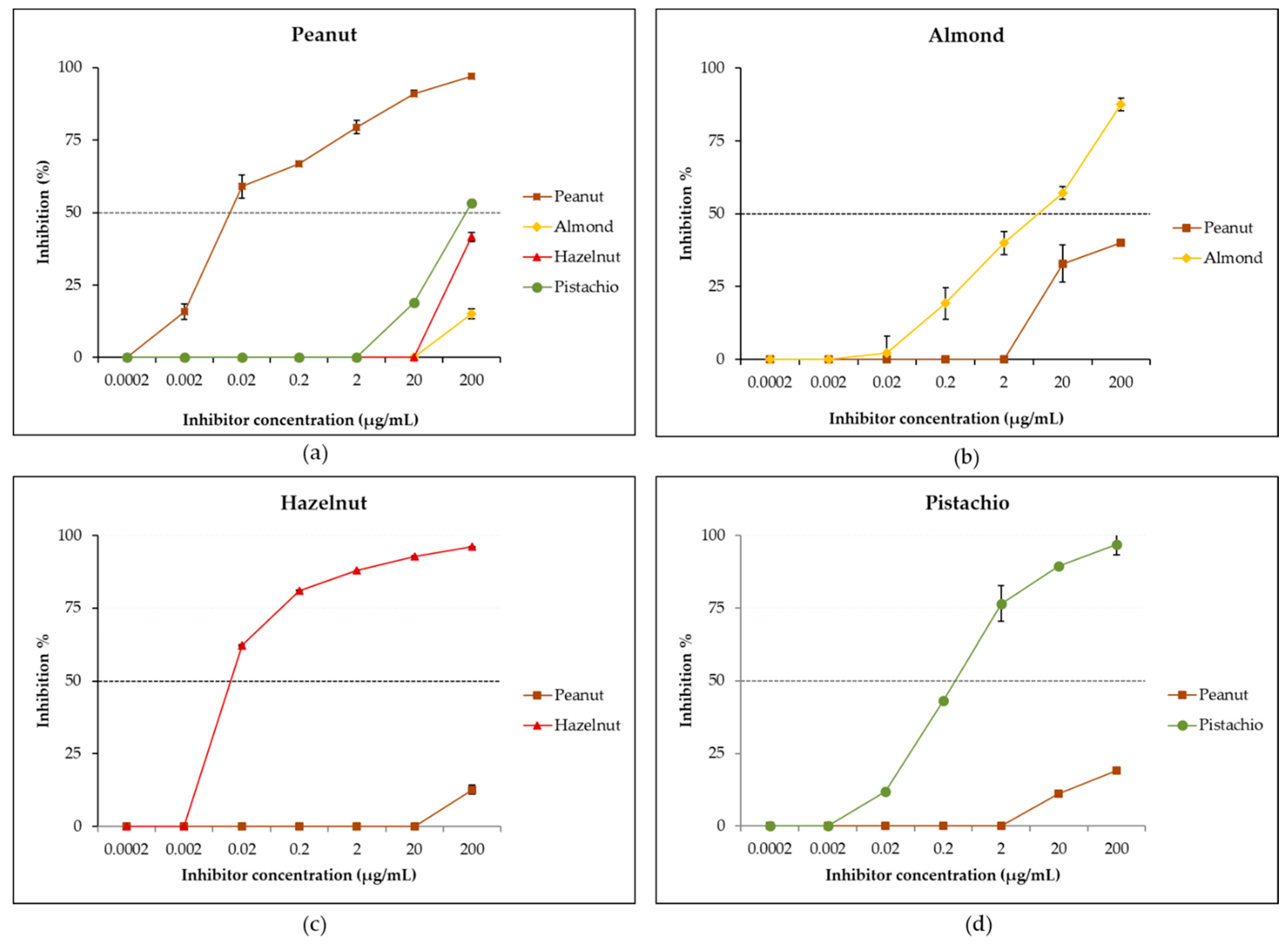
3.4. Evaluation of IgE Cross-Reactivity Between Peanut and Tree Nuts by Reciprocal Inhibition ELISA
3.5. Identification of IgE Cross-Reactive Proteins by Reciprocal Inhibition Immunoblotting
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gupta, R.S.; Springston, E.E.; Warrier, M.R.; Smith, B.; Kumar, R.; Pongracic, J.; Holl, J.L. The prevalence, severity, and distribution of childhood food allergy in the united states. Pediatrics 2011, 128, e9–e17. [Google Scholar] [CrossRef] [Green Version]
- Weinberger, T.; Sicherer, S. Current perspectives on tree nut allergy: A review. J. Asthma Allergy 2018, 11, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Shoshan, M.; Harrington, D.W.; Soller, L.; Fragapane, J.; Joseph, L.; St Pierre, Y.; Godefroy, S.B.; Elliot, S.J.; Clarke, A.E. A population-based study on peanut, tree nut, fish, shellfish, and sesame allergy prevalence in canada. J.Allergy Clin. Immunol. 2010, 125, 1327–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sicherer, S.H.; Sampson, H.A. Food allergy. J.Allergy Clin. Immunol. 2010, 125, S116–S125. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Warren, C.; Blumenstock, J.; Kotowska, J.; Mittal, K.; Smith, B. Or078 the prevalence of childhood food allergy in the united states: An update. Ann. Allergy Asthma Immunol. 2017, 119, S11. [Google Scholar] [CrossRef]
- Maloney, J.M.; Rudengren, M.; Ahlstedt, S.; Bock, S.A.; Sampson, H.A. The use of serum-specific ige measurements for the diagnosis of peanut, tree nut, and seed allergy. J. Allergy Clin. Immunol. 2008, 122, 145–151. [Google Scholar] [CrossRef]
- Matricardi, P.M.; Kleine-Tebbe, J.; Hoffmann, H.J.; Valenta, R.; Hilger, C.; Hofmaier, S.; Aalberse, R.C.; Agache, I.; Asero, R.; Ballmer-Weber, B.; et al. Eaaci molecular allergology user’s guide. Pediatric Allergy Immunol. 2016, 27, 1–250. [Google Scholar] [CrossRef]
- Aalberse, R.C. Assessment of allergen cross-reactivity. Clin. Mol. Allergy 2007, 5, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Bublin, M.; Breiteneder, H. Cross-reactivity of peanut allergens. Curr. Allergy Asthma Rep. 2014, 14. [Google Scholar] [CrossRef] [Green Version]
- Chan, E.S.; Greenhawt, M.J.; Fleischer, D.M.; Caubet, J.-C. Managing cross-reactivity in those with peanut allergy. J. Allergy Clin. Immunol. 2019, 7, 381–386. [Google Scholar] [CrossRef]
- Smeekens, J.M.; Bagley, K.; Kulis, M. Tree nut allergies: Allergen homology, cross-reactivity, and implications for therapy. Clin. Exp. Allergy 2018, 48, 762–772. [Google Scholar] [CrossRef] [PubMed]
- de Leon, M.P.; Glaspole, I.N.; Drew, A.C.; Rolland, J.M.; O’Hehir, R.E.; Suphioglu, C. Immunological analysis of allergenic cross-reactivity between peanut and tree nuts. Clin. Exp. Allergy 2003, 33, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Barre, A.; Jacquet, G.; Sordet, C.; Culerrier, R.; Rouge, P. Homology modelling and conformational analysis of ige-binding epitopes of ara h 3 and other legumin allergens with a cupin fold from tree nuts. Mol. Immunol. 2007, 44, 3243–3255. [Google Scholar] [CrossRef]
- Barre, A.; Sordet, C.; Culerrier, R.; Rance, F.; Didier, A.; Rouge, P. Vicilin allergens of peanut and tree nuts (walnut, Pistachio and cashew nut) share structurally related ige-binding epitopes. Mol. Immunol. 2008, 45, 1231–1240. [Google Scholar] [CrossRef]
- Aalberse, R.C. Structural biology of allergens. J. Allergy Clin. Immunol. 2000, 106, 228–238. [Google Scholar] [CrossRef]
- De Leon, M.P. Functional analysis of cross - reactive immunoglobulin e antibodies: Peanut-specific immunoglobulin e sensitizes basophils to tree nut allergens. Clin. Exp. Allergy 2005, 35, 1056. [Google Scholar] [CrossRef]
- de Leon, M.P.; Drew, A.C.; Glaspole, I.N.; Suphioglu, C.; O’Hehir, R.E.; Rolland, J.M. Ige cross-reactivity between the major peanut allergen ara h 2 and tree nut allergens. Mol. Immunol. 2007, 44, 463–471. [Google Scholar] [CrossRef]
- Eigenmann, P.A.; Lack, G.; Mazon, A.; Nieto, A.; Haddad, D.; Brough, H.A.; Caubet, J.-C. Managing nut allergy: A remaining clinical challenge. J. Allergy Clin. Immunol. 2017, 5, 296–300. [Google Scholar] [CrossRef]
- Elizur, A.; Appel, M.Y.; Nachshon, L.; Levy, M.B.; Epstein-Rigbi, N.; Golobov, K.; Goldberg, M.R. Nut co reactivity - acquiring knowledge for elimination recommendations (nut cracker) study. Allergy 2018, 73, 593–601. [Google Scholar] [CrossRef]
- Brough, H.A.; Caubet, J.-C. Selective nut-eating in peanut or tree nut allergic children—how can molecular allergology help? Clin. Exp. Allergy 2018, 48, 618–619. [Google Scholar] [CrossRef] [Green Version]
- Lieberman, J.A. Severity of peanut allergy and the unmet gaps in care: A call to action. Am. J. Manag. Care 2018, 24, S412–S418. [Google Scholar]
- Sampson, H.A. Food allergy. Part 2: Diagnosis and management. J. Allergy Clin. Immunol. 1999, 103, 981–989. [Google Scholar] [CrossRef]
- L’Hocine, L.; Pitre, M. Quantitative and qualitative optimization of allergen extraction from peanut and selected tree nuts. Part 2. Optimization of buffer and ionic strength using a full factorial experimental design. Food Chem. 2016, 194, 820–827. [Google Scholar]
- Venkatachalam, M.; Sathe, S.K. Chemical composition of selected edible nut seeds. J. Agric. Food Chem. 2006, 54, 4705–4714. [Google Scholar] [CrossRef]
- Sampson, H.A. Utility of food-specific ige concentrations in predicting symptomatic food allergy. J. Allergy Clin. Immunol. 2001, 107, 891–896. [Google Scholar] [CrossRef]
- Roberts, G.; Lack, G. Diagnosing peanut allergy with skin prick and specific ige testing. J. Allergy Clin. Immunol. 2005, 115, 1291–1296. [Google Scholar] [CrossRef]
- Clark, A.T.; Ewan, P.W. The development and progression of allergy to multiple nuts at different ages. Pediatr Allergy Immunol. 2005, 16, 507–511. [Google Scholar] [CrossRef]
- Glaspole, I.N.; de Leon, M.P.; Prickett, S.R.; O’Hehir, R.E.; Rolland, J.M. Clinical allergy to hazelnut and peanut: Identification of t cell cross-reactive allergens. Int. Arch. Allergy Immunol. 2011, 155, 345–354. [Google Scholar] [CrossRef]
- Breiteneder, H.; Mills, E.N.C. Food allergens: Molecular and immunological characteristics. In Food Allergy: Adverse Reactions to Foods and Food Additives, 4th ed.; Metcalfe, D.D.H.A., Sampson, H.A., Simon, R.A., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2009; pp. 43–61. [Google Scholar]
- Scheurer, S.; Toda, M.; Vieths, S. What makes an allergen? Clin. Exp. Allergy 2015, 45, 1150–1161. [Google Scholar] [CrossRef]
- International Union of Immunological Societies. Allergen Nomenclature. Available online: http://allergen.org (accessed on 23 May 2020).
- Allergome Platform. Available online: http://www.allergome.org/index.php (accessed on 11 May 2020).
- Lehmann, K.; Schweimer, K.; Reese, G.; Randow, S.; Suhr, M.; Becker, W.M.; Vieths, S.; Rosch, P. Structure and stability of 2s albumin-type peanut allergens: Implications for the severity of peanut allergic reactions. Biochem. J. 2006, 395, 463–472. [Google Scholar] [CrossRef]
- Chatel, J.M.; Bernard, H.; Orson, F.M. Isolation and characterization of two complete ara h 2 isoforms cdna. Int. Arch. Allergy Immunol. 2003, 131, 14–18. [Google Scholar] [CrossRef]
- Restani, P.; Ballabio, C.; Corsini, E.; Fiocchi, A.; Isoardi, P.; Magni, C.; Poiesi, C.; Terracciano, L.; Duranti, M. Identification of the basic subunit of ara h 3 as the major allergen in a group of children allergic to peanuts. Ann. Allergy Asthma Immunol. 2005, 94, 262–266. [Google Scholar] [CrossRef]
- Boldt, A.; Fortunato, D.; Conti, A.; Petersen, A.; Ballmer-Weber, B.; Lepp, U.; Reese, G.; Wolf-Meinhard, B. Analysis of the composition of an immunoglobulin e reactive high molecular weight protein complex of peanut extract containing ara h 1 and ara h 3/4. PROTEOMICS 2005, 5, 675–686. [Google Scholar] [CrossRef]
- Burks, A.W.; Williams, L.W.; Helm, R.M.; Connaughton, C.; Cockrell, G.; O’Brien, T. Identification of a major peanut allergen, ara h 1, in patients with atopic dermatitis and positive peanut challenges. J. Allergy Clin. Immunol. 1991, 88, 172–179. [Google Scholar] [CrossRef]
- Burks, A.W.; Cockrell, G.; Stanley, J.S.; Helm, R.M.; Bannon, G.A. Recombinant peanut allergen ara h 1 expression and ige binding in patients with peanut hypersensitivity. J. Clin. Investig. 1995, 96, 1715–1721. [Google Scholar] [CrossRef] [Green Version]
- Kottapalli, K.R.; Payton, P.; Rakwal, R.; Agrawal, G.K.; Shibato, J.; Burow, M.; Puppala, N. Proteomics analysis of mature seed of four peanut cultivars using two-dimensional gel electrophoresis reveals distinct differential expression of storage, anti-nutritional, and allergenic proteins. Plant Sci. 2008, 175, 321–329. [Google Scholar] [CrossRef]
- Sathe, S.K.; Wolf, W.J.; Roux, K.H.; Teuber, S.S.; Venkatachalam, M.; Sze-Tao, K.W. Biochemical characterization of amandin, the major storage protein in almond (prunus dulcis l.). J. Agric. Food Chem. 2002, 50, 4333–4341. [Google Scholar] [CrossRef]
- UniProt Consortium. Uniprotkb. Available online: https://www.uniprot.org/ (accessed on 23 May 2020).
- Buhler, S.; Tedeschi, T.; Faccini, A.; Garino, C.; Arlorio, M.; Dossena, A.; Sforza, S. Isolation and full characterisation of a potentially allergenic lipid transfer protein (ltp) in almond. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2015, 32, 648–656. [Google Scholar]
- Poltronieri, P.; Cappello, M.S.; Dohmae, N.; Conti, A.; Fortunato, D.; Pastorello, E.A.; Ortolani, C.; Zacheo, G. Identification and characterisation of the ige-binding proteins 2s albumin and conglutin gamma in almond (prunus dulcis) seeds. Int. Arch. Allergy Immunol. 2002, 128, 97–104. [Google Scholar] [CrossRef]
- Abolhassani, M.; Roux, K.H. Cdna cloning, expression and characterization of an allergenic 60s ribosomal protein of almond (prunus dulcis). Iran. J. Allergy Asthma Immunol. 2009, 8, 77–84. [Google Scholar]
- Chen, L.; Zhang, S.; Illa, E.; Song, L.; Wu, S.; Howad, W.; Arús, P.; van de Weg, E.; Chen, K.; Gao, Z. Genomic characterization of putative allergen genes in peach/almond and their synteny with apple. BMC Genom. 2008, 9, 543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Che, H.; Zhang, Y.; Jiang, S.; Jin, T.; Lyu, S.-C.; Nadeau, K.C.; McHugh, T. Almond (prunus dulcis) allergen pru du 8, the first member of a new family of food allergens. J. Agric. Food Chem. 2019, 67, 8626–8631. [Google Scholar] [CrossRef] [PubMed]
- Rigby, N.M.; Marsh, J.; Sancho, A.I.; Wellner, K.; Akkerdaas, J.; van Ree, R.; Knulst, A.; Fernández-Rivas, M.; Brettlova, V.; Schilte, P.P.; et al. The purification and characterisation of allergenic hazelnut seed proteins. Mol. Nutr. Food Res. 2008, 52, S251–S261. [Google Scholar]
- Geiselhart, S.; Hoffmann-Sommergruber, K.; Bublin, M. Tree nut allergens. Mol. Immunol. 2018, 100, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Parra, F.M.; Cuevas, M.; Lezaun, A.; Alonso, M.D.; Beristain, A.M.; Losada, E. Pistachio nut hypersensitivity: Identification of pistachio nut allergens. Clin. Exp. Allergy 1993, 23, 996–1001. [Google Scholar] [CrossRef] [PubMed]
- Fernández, F.A.; Fiandor, A.; Martinez-Garate, A.; Martinez Quesada, J. Allergy to pistachio: Crossreactivity between pistachio nut and other anacardiaceae. Clin. Exp. Allergy 1995, 25, 1254–1259. [Google Scholar]
- Garcia, F.; Moneo, I.; Fernandez, B.; Garcia-Menaya, J.M.; Blanco, J.; Juste, S.; Gonzalo, M.A. Allergy to anacardiaceae: Description of cashew and pistachio nut allergens. J. Investig. Allergol. Clin. Immunol. 2000, 10, 173–177. [Google Scholar]
- Ahn, K.; Bardina, L.; Grishina, G.; Beyer, K.; Sampson, H.A. Identification of two pistachio allergens, pis v 1 and pis v 2, belonging to the 2s albumin and 11s globulin family. Clin. Exp. Allergy 2009, 39, 926–934. [Google Scholar] [CrossRef]
- Vicente, F.; Sanchiz, A.; Rodríguez-Pérez, R.; Pedrosa, M.; Quirce, S.; Haddad, J.; Besombes, C.; Linacero, R.; Allaf, K.; Cuadrado, C. Influence of instant controlled pressure drop (dic) on allergenic potential of tree nuts. Molecules 2020, 25. [Google Scholar] [CrossRef] [Green Version]
- Sanchiz, A.; Cuadrado, C.; Dieguez, M.C.; Ballesteros, I.; Rodríguez, J.; Crespo, J.F.; de las Cuevas, N.; Rueda, J.; Linacero, R.; Cabanillas, B.; et al. Thermal processing effects on the ige-reactivity of cashew and pistachio. Food Chem. 2018, 245, 595–602. [Google Scholar] [CrossRef]
- Krause, S.; Reese, G.; Randow, S.; Zennaro, D.; Quaratino, D.; Palazzo, P.; Ciardiello, M.A.; Petersen, A.; Becker, W.-M.; Mari, A. Lipid transfer protein (ara h 9) as a new peanut allergen relevant for a mediterranean allergic population. J. Allergy Clin. Immunol. 2009, 124, 771–778.e775. [Google Scholar] [CrossRef]
- Iqbal, A.; Shah, F.; Hamayun, M.; Ahmad, A.; Hussain, A.; Waqas, M.; Kang, S.-M.; Lee, I.-J. Allergens of arachis hypogaea and the effect of processing on their detection by elisa. Food Nutr. Res. 2016, 60, 28945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, H.; Krause, S.; Gelhaus, C.; Petersen, A.; Janssen, O.; Becker, W.-M. Detection and structural characterization of natural ara h 7, the third peanut allergen of the 2s albumin family. J. Proteome Res. 2010, 9, 3701–3709. [Google Scholar] [CrossRef] [PubMed]
- Pons, L.; Chery, C.; Romano, A.; Namour, F.; Artesani, M.C.; Gueant, J.L. The 18 kda peanut oleosin is a candidate allergen for ige-mediated reactions to peanuts. Allergy 2002, 57 (Suppl. 72), 88–93. [Google Scholar] [CrossRef]
- Hales, B.J.; Bosco, A.; Mills, K.L.; Hazell, L.A.; Loh, R.; Holt, P.G.; Thomas, W.R. Isoforms of the major peanut allergen ara h 2: Ige binding in children with peanut allergy. Int. Arch. Allergy Immunol. 2004, 135, 101–107. [Google Scholar] [CrossRef]
- Piersma, S.R.; Gaspari, M.; Hefle, S.L.; Koppelman, S.J. Proteolytic processing of the peanut allergen ara h 3. Mol. Nutr. Food Res. 2005, 49, 744–755. [Google Scholar] [CrossRef]
- L’Hocine, L.; Pitre, M. Quantitative and qualitative optimization of allergen extraction from peanut and selected tree nuts. Part 1. Screening of optimal extraction conditions using a d-optimal experimental design. Food Chem. 2016, 194, 780–786. [Google Scholar]
- Jin, T.; Guo, F.; Chen, Y.-W.; Howard, A.; Zhang, Y. Crystal structure of ara h 3, a major allergen in peanut. Mol. Immunol. 2009, 46, 1796–1804. [Google Scholar] [CrossRef]
- Yusnawan, E.; Marquis, C.P.; Lee, N.A. Purification and characterization of ara h 1 and ara h 3 from four peanut market types revealed higher order oligomeric structures. J. Agric.Food Chem. 2012, 60, 10352–10358. [Google Scholar] [CrossRef]
- Garino, C.; De Paolis, A.; Coïsson, J.D.; Arlorio, M. Pru du 2s albumin or pru du vicilin? Comput. Biol. Chem. 2015, 56, 30–32. [Google Scholar] [CrossRef]
- Tawde, P.; Venkatesh, Y.P.; Wang, F.; Teuber, S.S.; Sathe, S.K.; Roux, K.H. Cloning and characterization of profilin (pru du 4), a cross-reactive almond (prunus dulcis) allergen. J. Allergy Clin. Immunol. 2006, 118, 915–922. [Google Scholar] [CrossRef]
- Mandalari, G.; Mackie, A.R. Almond allergy: An overview on prevalence, thresholds, regulations and allergen detection. Nutrients 2018, 10, 1706. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Jin, T. Almond allergens: Update and perspective on identification and characterization. J. Sci. Food Agric. 2020. [Google Scholar] [CrossRef]
- Albillos, S.M.; Jin, T.; Howard, A.; Zhang, Y.; Kothary, M.H.; Fu, T.J. Purification, crystallization and preliminary x-ray characterization of prunin-1, a major component of the almond (prunus dulcis) allergen amandin. J. Agric. Food Chem. 2008, 56, 5352–5358. [Google Scholar] [CrossRef]
- Dreveny, I.; Gruber, K.; Glieder, A.; Thompson, A.; Kratky, C. The hydroxynitrile lyase from almond: A lyase that looks like an oxidoreductase. Structure 2001, 9, 803–815. [Google Scholar] [CrossRef] [Green Version]
- Garino, C.; Zuidmeer, L.; Marsh, J.; Lovegrove, A.; Morati, M.; Versteeg, S.; Schilte, P.; Shewry, P.; Arlorio, M.; van Ree, R. Isolation, cloning, and characterization of the 2s albumin: A new allergen from hazelnut. Mol. Nutr. Food Res. 2010, 54, 1257–1265. [Google Scholar] [CrossRef]
- Hansen, K.S.; Ballmer-Weber, B.K.; Sastre, J.; Lidholm, J.; Andersson, K.; Oberhofer, H.; Lluch-Bernal, M.; Östling, J.; Mattsson, L.; Schocker, F.; et al. Component-resolved in vitro diagnosis of hazelnut allergy in europe. J. Allergy Clin. Immunol. 2009, 123, 1134–1141.e1133. [Google Scholar] [CrossRef]
- Zuidmeer-Jongejan, L.; Fernández-Rivas, M.; Winter, M.G.; Akkerdaas, J.H.; Summers, C.; Lebens, A.; Knulst, A.C.; Schilte, P.; Briza, P.; Gadermaier, G.; et al. Oil body-associated hazelnut allergens including oleosins are underrepresented in diagnostic extracts but associated with severe symptoms. Clin. Transl. Allergy 2014, 4, 4. [Google Scholar] [CrossRef] [Green Version]
- Pastorello, E.A.; Stefan, V.; Valerio, P.; Laura, F.; Chiara, T.; Donatella, F.; Dirk, L.t.; Marianna, C.; Raffaella, A.; Joseph, S.; et al. Identification of hazelnut major allergens in sensitive patients with positive double-blind, placebo-controlled food challenge results. J. Allergy Clin. Immunol. 2002, 109, 563–570. [Google Scholar] [CrossRef]
- Guo, F.; Kothary, M.H.; Wang, Y.; Yu, X.; Howard, A.J.; Fu, T.J.; Zhang, Y.Z. Purification and crystallization of cor a 9, a major hazelnut allergen. Acta Crystallogr. Sec. F 2009, 65, 42–46. [Google Scholar] [CrossRef] [Green Version]
- Lauer, I.; Foetisch, K.; Kolarich, D.; Ballmer-Weber, B.K.; Conti, A.; Altmann, F.; Vieths, S.; Scheurer, S. Hazelnut (corylus avellana) vicilin cor a 11: Molecular characterization of a glycoprotein and its allergenic activity. Biochem. J. 2004, 383, 327–334. [Google Scholar] [CrossRef] [Green Version]
- Gruehn, S.; Suphioglu, C.; O’Hehir, R.E.; Volkmann, D. Molecular cloning and characterization of hazel pollen protein (70 kd) as a luminal binding protein (bip): A novel cross-reactive plant allergen. Int. Arch. Allergy Immunol. 2003, 131, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Noorbakhsh, R.; Mortazavi, S.A.; Sankian, M.; Shahidi, F.; Assarehzadegan, M.A.; Varasteh, A. Cloning, expression, characterization, and computational approach for cross-reactivity prediction of manganese superoxide dismutase allergen from pistachio nut. Allergol. Int. 2010, 59, 295–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willison, L.N.; Tawde, P.; Robotham, J.M.; Penney, R.M.t.; Teuber, S.S.; Sathe, S.K.; Roux, K.H. Pistachio vicilin, pis v 3, is immunoglobulin e-reactive and cross-reacts with the homologous cashew allergen, ana o 1. Clin. Exp. Allergy 2008, 38, 1229–1238. [Google Scholar] [CrossRef] [PubMed]
- van Erp, F.C.; Knol, E.F.; Pontoppidan, B.; Meijer, Y.; van der Ent, C.K.; Knulst, A.C. The ige and basophil responses to ara h 2 and ara h 6 are good predictors of peanut allergy in children. J. Allergy Clin. Immunol. 2017, 139, 358–360.e358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyer, K.; Morrowa, E.; Li, X.-M.; Bardina, L.; Bannon, G.A.; Burks, A.W.; Sampson, H.A. Effects of cooking methods on peanut allergenicity. J. Allergy Clin. Immunol. 2001, 107, 1077–1081. [Google Scholar] [CrossRef]
- Cabanillas, B.; Jappe, U.; Novak, N. Allergy to peanut, soybean, and other legumes: Recent advances in allergen characterization, stability to processing and ige cross-reactivity. Mol. Nutr. Food Res. 2018, 62, 1700446. [Google Scholar] [CrossRef]
- Ballmer-Weber, B.K.; Lidholm, J.; Fern#xE1;ndez-Rivas, M.; Seneviratne, S.; Hanschmann, K.-M.; Vogel, L.; Bures, P.; Fritsche, P.; Summers, C.; Knulst, A.C.; et al. Ige recognition patterns in peanut allergy are age dependent: Perspectives of the europrevall study. Allergy 2015, 70, 391–407. [Google Scholar] [CrossRef]
- Vereda, A.; van Hage, M.; Ahlstedt, S.; Ibañez, M.D.; Cuesta-Herranz, J.; van Odijk, J.; Wickman, M.; Sampson, H.A. Peanut allergy: Clinical and immunologic differences among patients from 3 different geographic regions. J. Allergy Clin. Immunol. 2011, 127, 603–607. [Google Scholar] [CrossRef]
- Garcia-Blanca, A.; Aranda, A.; Blanca-Lopez, N.; Perez, D.; Gomez, F.; Mayorga, C.; Torres, M.J.; Diaz-Perales, A.; Perkins, J.R.; Villalba, M.; et al. Influence of age on ige response in peanut-allergic children and adolescents from the mediterranean area. Pediatric Allergy Immunol. 2015, 26, 497–502. [Google Scholar] [CrossRef] [Green Version]
- Namork, E.; Stensby, B.A. Peanut sensitization pattern in norwegian children and adults with specific ige to peanut show age related differences. Allergy Asthma Clin. Immunol. 2015, 11, 32. [Google Scholar] [CrossRef] [Green Version]
- Valcour, A.; Jones, J.E.; Lidholm, J.; Borres, M.P.; Hamilton, R.G. Sensitization profiles to peanut allergens across the united states. Ann.Allergy Asthma Immunol. 2017, 119, 262–266.e261. [Google Scholar] [CrossRef] [Green Version]
- Klemans, R.J.B.; van Os-Medendorp, H.; Blankestijn, M.; Bruijnzeel-Koomen, C.A.F.M.; Knol, E.F.; Knulst, A.C. Diagnostic accuracy of specific ige to components in diagnosing peanut allergy: A systematic review. Clin. Exp. Allergy 2015, 45, 720–730. [Google Scholar] [CrossRef] [PubMed]
- Barocci, F.; M, D.E.A.; Marseglia, G.L. Molecular evolution in food allergy diagnosis. Minerva pediatrica 2016, 68, 374–381. [Google Scholar]
- Noorbakhsh, R. Pistachio allergy - prevalence and in vitro cross - reactivity with other nuts. Allergol. Int. 2011, 60, 425–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akkerdaas, J.H.; Schocker, F.; Vieths, S.; Versteeg, S.; Zuidmeer, L.; Hefle, S.L.; Aalberse, R.C.; Richter, K.; Ferreira, F.; van Ree, R. Cloning of oleosin, a putative new hazelnut allergen, using a hazelnut cdna library. Mol. Nutr. Food Res. 2006, 50, 18–23. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient No. | Age (years) | Sex | Symptoms to Peanut 1 | SPT (mm) | Specific IgE (kU/L) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Negative Control | Histamine | Peanut Extract | Peanut | Almond | Hazelnut | Pistachio | ||||
| 1 | 3 | M | eczema | 0 | 4 | 8.0 | ˃100 | 0 | 0 | 0 |
| 2 | 4 | M | eczema | 0 | 4 | 8.5 | ˃100 | 1.13 | ˃100 | 4.36 |
| 3 | 3 | M | eczema | 0 | 3.5 | 5.0 | ˃100 | 1.44 | 3.32 | 13.4 |
| 4 | 12 | M | skin rash and eye swelling | 0 | 5 | 12.0 | ˃100 | 4.28 | 16.0 | 3.58 |
| 5 | 5 | M | urticaria | 0 | 3 | 14.5 | ˃100 | 8.73 | 60.2 | 3.95 |
| 6 | 12 | M | Urticaria and eye swelling | 0 | 5 | 15.0 | ˃100 | 0.67 | 0.48 | 0.21 |
| 7 | 13 | M | ocular pruritus, eye swelling, urticaria and rhinorrhea and, 2 hours later, cough, dyspnea and wheezing | 0 | 4 | 14.0 | ˃100 | ˂0.1 | 0.67 | 4.93 |
| 8 | 9 | M | skin rash, swelling, cough, dyspnea and vomiting | 0 | 3 | 8.5 | ˃100 | 0.534 | 0.878 | 12.6 |
| 9 | 9 | M | urticaria | 0 | 3 | 6.0 | ˃100 | 1.69 | 6.41 | 3.46 |
| Peak # | Allergen 1 | Protein Family | Molecular Weight on SDS-PAGE 2 (kDa) | Reference |
|---|---|---|---|---|
| Peanut (Arachis hypogea) | ||||
| P1 | Ara h 9 | Nonspecific lipid-transfer protein (LTP) | 9.8 | [55] |
| P2 | Ara h 5 | Profilin | 12-15 | [56] |
| P3 | Ara h 6 | Conglutin 2S albumin | 14.2 | [33] |
| P4 | Ara h 7 | Conglutin 2S albumin | 15.8 | [57] |
| P5 | Ara h 10 | Oleosin | 16 | [58] |
| P6 | Ara h 2.01 | Conglutin 2S albumin | 17 | [34] |
| P7 | Ara h 2.02 | Conglutin 2S albumin | 19 | [59] |
| P8/P9 | Ara h 3/4 | Legumin 11S globulin basic subunit | 23–25 | [60] |
| P10-P14 | Ara h 3 | Legumin 11S globulin acidic subunit | 30–37 | [32] |
| P15 | Ara h 3 | Legumin 11S globulin acidic subunit | 42–45 | [60,61] |
| P16 | Ara h 3 | Legumin 11S globulin | 60 | [62,63] |
| P17 | Ara h 1 | Vicilin 7S globulin | 64 | [38] |
| P18 | Ara h 10 | Oleosin oligomers | 67–85 | [58] |
| P19 | Lipoxygenase | 95–100 | [39] | |
| Almond (Prunus dulcis) | ||||
| A1 | Pru du 5 | Ribosomal protein | 10 | [44] |
| A2 | Pru du 2S albumin | Conglutin 2S albumin | 12 | [64] |
| A3 | Pru du 4 | Profilin | 14 | [65] |
| A4 | Pru du 1 | PR-10 protein | 17 | [32] |
| A5 | Pru du 6 | Legumin 11 S globulin (Amandin) | 20–22 | [66] |
| A6 | Pru du 2 | PR-5 protein (Thaumatin-like protein) | 26 | [67] |
| A7 | Pru du 8 | Antimicrobial seed storage protein | 31 | [46] |
| A8 | Pru du 6 | Legumin 11 S globulin (Amandin) | 40 | [68] |
| A9 | Pru du γ-conglutin | 7S vicilin storage protein | 45 | [67] |
| A11 | Pru du 10 | Mandelonitrile lyase 2 | 60 | [69] |
| Hazelnut (Corylus avellana) | ||||
| H1 | Cor a 8 | Nonspecific lipid-transfer protein (LTP) | 9 | [47] |
| H2 | Cor a 14 | Conglutin 2S albumin | 11 | [70] |
| H3 | Cor a 2 | Profilin | 14 | [71] |
| H4 | Cor a 13 | Oleosin | 14–16 | [72] |
| H5 | Cor a 12 | Oleosin | 17 | [72] |
| H6 | Cor a 1 | Pathogenesis-related protein | 17 | [73] |
| H7-H10 | Cor a 9 | Legumin 11S globulin | 21–25; 31–35 | [47,74] |
| H11 | Cor a 11 | Vicilin 7S globulin | 48 | [47] |
| H12 | Cor a 11 | Vicilin 7S globulin | 55 | [75] |
| H13 | Cor a 10 | Luminal binding protein | 70 | [76] |
| Pistachio (Pistachia vera) | ||||
| Pi1/Pi2 | Pis v 1 | Conglutin 2S albumin | 7 (17) | [48] |
| Pi3 | Pis v 4 | Mn superoxide dismutase | 26 | [77] |
| Pi4 | Pis v 2 | Legumin 11S globulin | 32 | [52] |
| Pi5 | Pis v 5 | Legumin 11S globulin | 36 | |
| Pi6/Pi7 | Pis v 2 | Legumin 11S globulin | 53 | [52] |
| Pi8 | Pis v 3 | Vicilin 7S globulin | 55 (59) | [78] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pitre, M.; L’Hocine, L.; Achouri, A.; Blaquière, M.; Des Roches, A. Immunoglobulin E-Binding Pattern of Canadian Peanut Allergic Children and Cross-Reactivity with Almond, Hazelnut and Pistachio. Biomolecules 2020, 10, 1091. https://doi.org/10.3390/biom10081091
Pitre M, L’Hocine L, Achouri A, Blaquière M, Des Roches A. Immunoglobulin E-Binding Pattern of Canadian Peanut Allergic Children and Cross-Reactivity with Almond, Hazelnut and Pistachio. Biomolecules. 2020; 10(8):1091. https://doi.org/10.3390/biom10081091
Chicago/Turabian StylePitre, Mélanie, Lamia L’Hocine, Allaoua Achouri, Martin Blaquière, and Anne Des Roches. 2020. "Immunoglobulin E-Binding Pattern of Canadian Peanut Allergic Children and Cross-Reactivity with Almond, Hazelnut and Pistachio" Biomolecules 10, no. 8: 1091. https://doi.org/10.3390/biom10081091