Role of RadA and DNA Polymerases in Recombination-Associated DNA Synthesis in Hyperthermophilic Archaea

Abstract
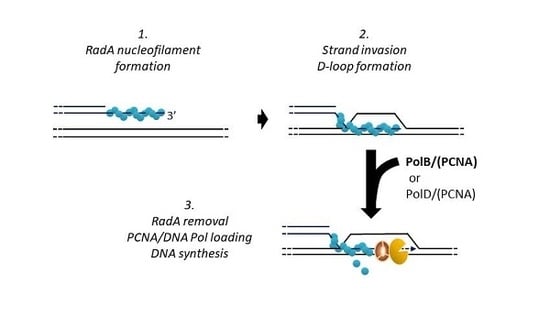
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Protein Expression and Purification
2.1.1. RadA
2.1.2. PCNA (Proliferating Cell Nuclear Antigen)
2.1.3. PolB
2.1.4. PolD
2.2. DNA Substrates
2.2.1. Circular D-Loop
2.2.2. Linear D-Loop
2.2.3. Linear Primer/Template
2.3. Enzymatic Assays
2.3.1. D-Loop Formation Assays
2.3.2. Circular D-Loop Extension Assays
2.3.3. Linear D-Loop Extension Assays
2.3.4. Strand Displacement Assays
3. Results and Discussion
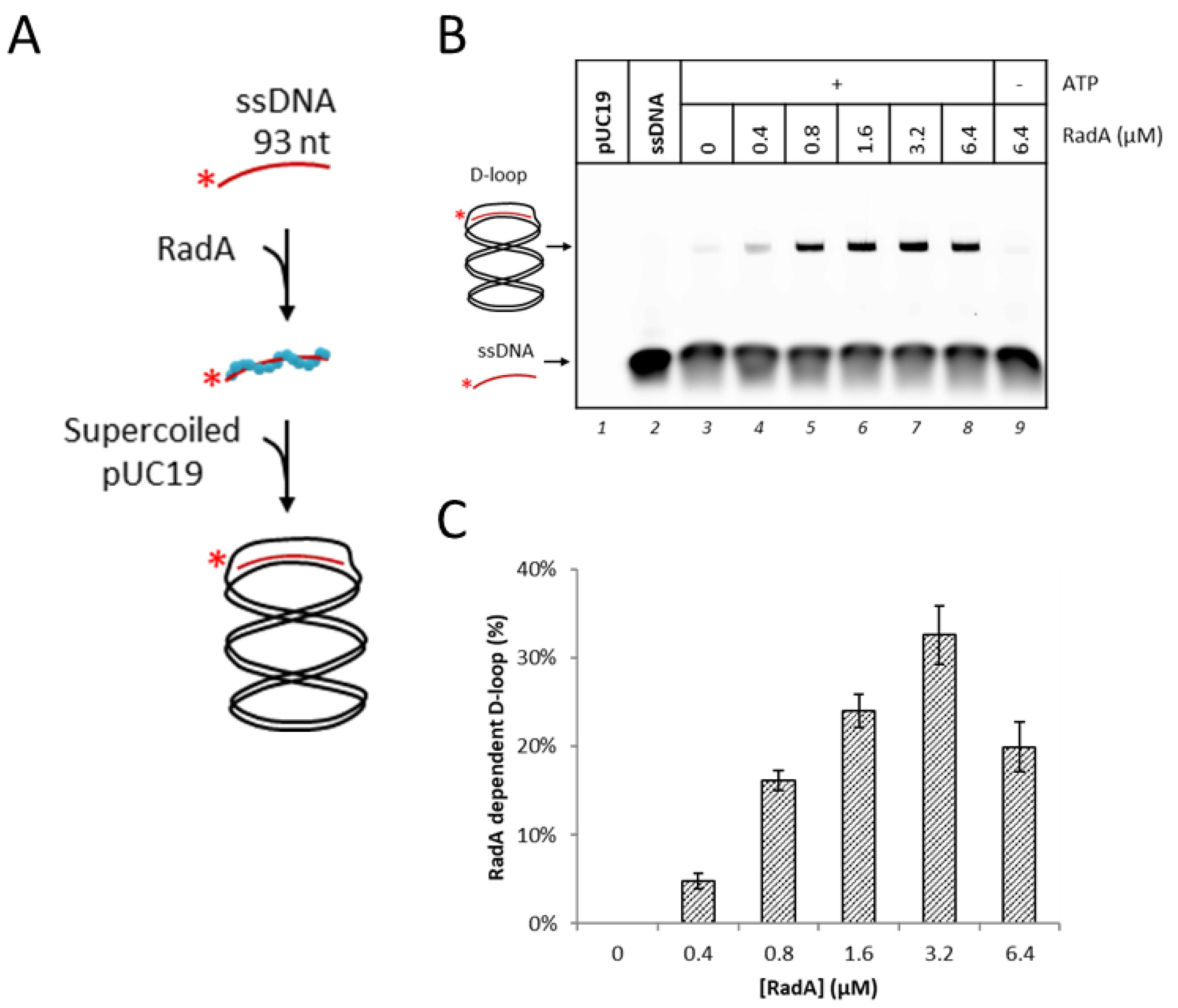
3.1. RadA Catalyzes D-Loop Formation on a Circular DNA Substrate
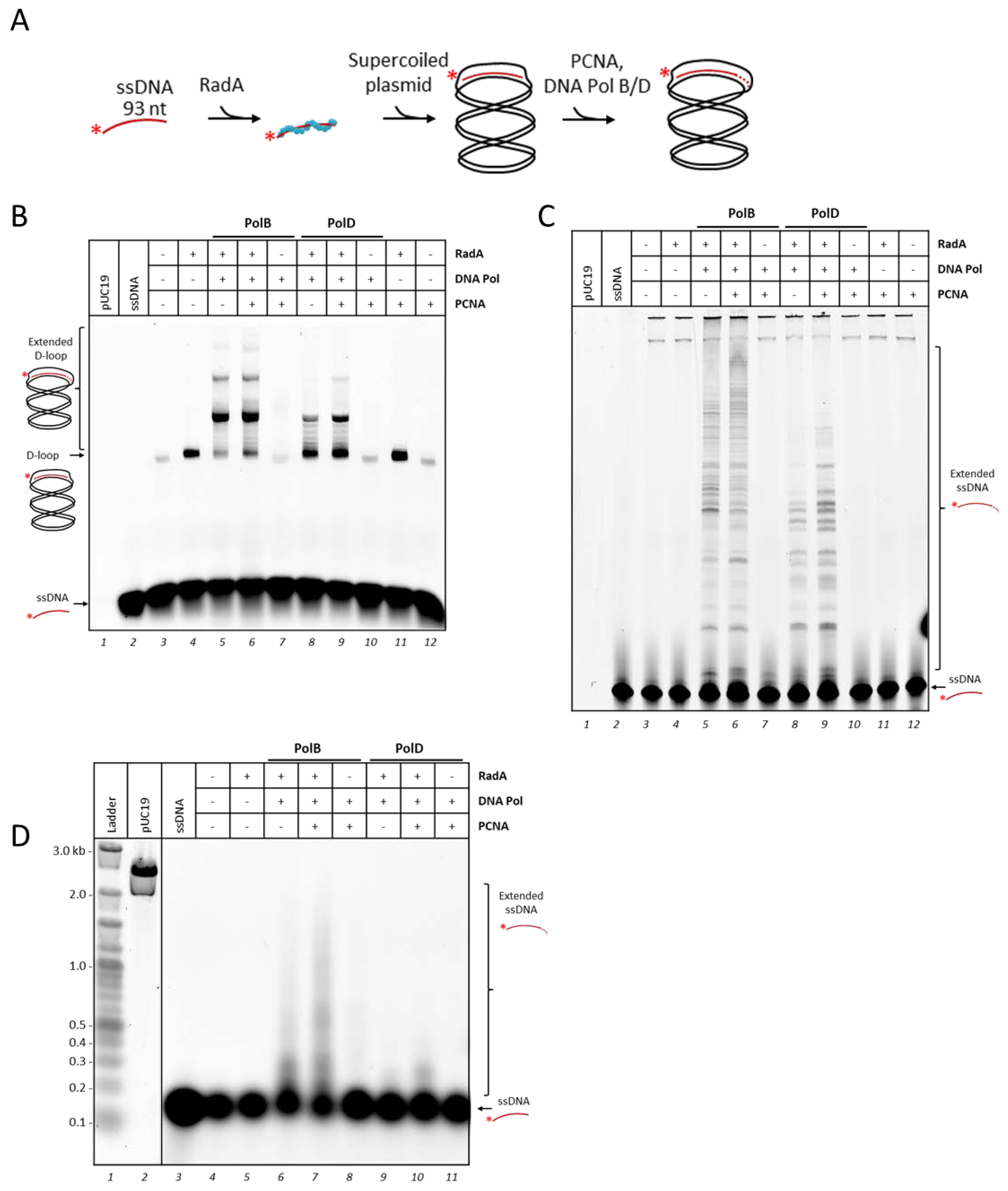
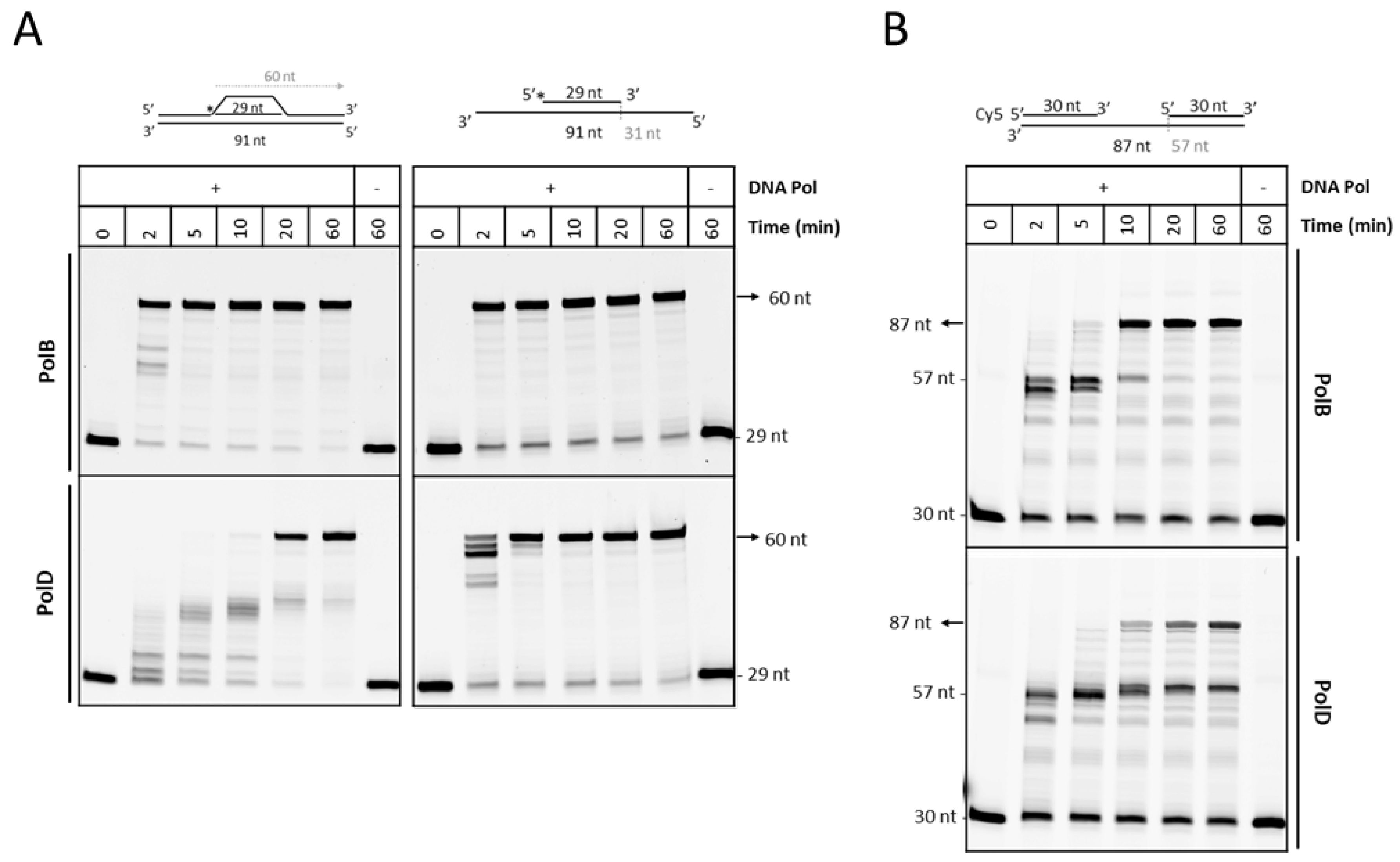
3.2. P. abyssi DNA Polymerases Extend RadA-Dependent D-Loops
3.3. PolD is Less Efficient Compare to PolB to Extend D-loop Like Substrate
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wright, W.D.; Shah, S.S.; Heyer, W.-D. Homologous recombination and the repair of DNA double-strand breaks. J. Biol. Chem. 2018, 293, 10524–10535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pannunzio, N.R.; Watanabe, G.; Lieber, M.R. Nonhomologous DNA end-joining for repair of DNA double-strand breaks. J. Biol. Chem. 2017, 293, 10512–10523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sallmyr, A.; Tomkinson, A.E. Repair of DNA double-strand breaks by mammalian alternative end-joining pathways. J. Biol. Chem. 2018, 293, 10536–10546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, M.F.; Allers, T. DNA repair in the archaea—An emerging picture. FEMS Microbiol. Rev. 2018, 42, 514–526. [Google Scholar] [CrossRef]
- Fujikane, R.; Ishino, S.; Ishino, Y.; Forterre, P. Genetic analysis of DNA repair in the hyperthermophilic archaeon, Thermococcus kodakaraensis. Genes Genet. Syst. 2010, 85, 243–257. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Tian, B.; Li, S.; Ao, X.; Dalgaard, K.; Gokce, S.; Liang, Y.; She, Q. Genetic manipulation in Sulfolobus islandicus and functional analysis of DNA repair genes. Biochem. Soc. Trans. 2013, 41, 405–410. [Google Scholar] [CrossRef] [Green Version]
- Huang, Q.; Liu, L.; Liu, J.; Ni, J.; She, Q.; Shen, Y. Efficient 5′-3′ DNA end resection by HerA and NurA is essential for cell viability in the crenarchaeon Sulfolobus islandicus. BMC Mol. Biol. 2015, 16, 2. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, M.; Malla, S.; Blythe, M.J.; Nieduszynski, C.A.; Allers, T. Accelerated growth in the absence of DNA replication origins. Nature 2013, 503, 544–547. [Google Scholar] [CrossRef]
- Gehring, A.M.; Astling, D.P.; Matsumi, R.; Burkhart, B.W.; Kelman, Z.; Reeve, J.N.; Jones, K.L.; Santangelo, T.J. Genome Replication in Thermococcus kodakarensis Independent of Cdc6 and an Origin of Replication. Front. Microbiol. 2017, 8, 2084. [Google Scholar] [CrossRef]
- Komori, K.; Miyata, T.; Daiyasu, H.; Toh, H.; Shinagawa, H.; And, Y.I. Domain Analysis of an Archaeal RadA Protein for the Strand Exchange Activity. J. Biol. Chem. 2000, 275, 33791–33797. [Google Scholar] [CrossRef] [Green Version]
- Komori, K.; Miyata, T.; DiRuggiero, J.; Holley-Shanks, R.; Hayashi, I.; Cann, I.K.O.; Mayanagi, K.; Shinagawa, H.; Ishino, Y. Both RadA and RadB are involved in homologous recombination inPyrococcus furiosus. J. Biol. Chem. 2000, 275, 33782–33790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renkawitz, J.; Lademann, C.A.; Jentsch, S. Mechanisms and principles of homology search during recombination. Nat. Rev. Mol. Cell Biol. 2014, 15, 369–383. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-W.; Ko, T.-P.; Lee, C.-D.; Chang, Y.-C.; Lin, K.-A.; Chang, C.-S.; Wang, A.H.-J.; Wang, T.-F. Three new structures of left-handed RadA helical filaments: Structural flexibility of N-Terminal domain is critical for recombinase activity. PLoS ONE 2009, 4, e4890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McRobbie, A.-M.; Carter, L.G.; Kerou, M.; Liu, H.; McMahon, S.A.; Johnson, K.A.; Oke, M.; Naismith, J.H.; White, M.F. Structural and Functional Characterisation of a Conserved Archaeal RadA Paralog with Antirecombinase Activity. J. Mol. Boil. 2009, 389, 661–673. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Lipfert, J.; Sánchez, H.; Wyman, C.; Dekker, N.H. Structural and torsional properties of the RAD51-dsDNA nucleoprotein filament. Nucleic Acids Res. 2013, 41, 7023–7230. [Google Scholar] [CrossRef]
- Short, J.M.; Liu, Y.; Chen, S.; Soni, N.; Madhusudhan, M.S.; Shivji, M.K.; Venkitaraman, A.R. High-resolution structure of the presynaptic RAD51 filament on single-stranded DNA by electron cryo-microscopy. Nucleic Acids Res. 2016, 44, 9017–9030. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Zhao, L.; Xu, Y.; Zhao, W.; Sung, P.; Wang, H.-W. Cryo-EM structures of human RAD51 recombinase filaments during catalysis of DNA-strand exchange. Nat. Struct. Mol. Biol. 2016, 24, 40–46. [Google Scholar] [CrossRef] [Green Version]
- Brouwer, I.; Moschetti, T.; Candelli, A.; Garcin, E.B.; Modesti, M.; Pellegrini, L.; Wuite, G.J.L.; Peterman, E.J.G. Two distinct conformational states define the interaction of human RAD 51-ATP with single-stranded DNA. EMBO J. 2018, 37, e98162. [Google Scholar] [CrossRef]
- Shin, D.S.; Pellegrini, L.; Daniels, D.; Yelent, B.; Craig, L.; Bates, D.; Yu, D.S.; Shivji, M.K.; Hitomi, C.; Arvai, A.S.; et al. Full-length archaeal Rad51 structure and mutants: Mechanisms for RAD51 assembly and control by BRCA2. EMBO J. 2003, 22, 4566–4576. [Google Scholar] [CrossRef] [Green Version]
- Pomerantz, R.T.; Goodman, M.F.; O’Donnell, M. DNA polymerases are error-prone at RecA-mediated recombination intermediates. Cell Cycle 2013, 12, 2558–2563. [Google Scholar] [CrossRef] [Green Version]
- Pomerantz, R.T.; Kurth, I.; Goodman, M.F.; E O’Donnell, M.; O’Donnell, M. Preferential D-loop extension by a translesion DNA polymerase underlies error-prone recombination. Nat. Struct. Mol. Biol. 2013, 20, 748–755. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Marians, K.J. A Dynamic RecA Filament Permits DNA Polymerase-catalyzed Extension of the Invading Strand in Recombination Intermediates. J. Biol. Chem. 2002, 277, 14321–14328. [Google Scholar] [CrossRef] [Green Version]
- McIlwraith, M.; Vaisman, A.; Liu, Y.; Fanning, E.; Woodgate, R.; West, S. Human DNA Polymerase η Promotes DNA Synthesis from Strand Invasion Intermediates of Homologous Recombination. Mol. Cell 2005, 20, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Stith, C.M.; Burgers, P.M.; Heyer, W.-D. PCNA Is Required for Initiation of Recombination-Associated DNA Synthesis by DNA Polymerase δ. Mol. Cell 2009, 36, 704–713. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Holzschu, D.L.; Sugiyama, T. PCNA is efficiently loaded on the DNA recombination intermediate to modulate polymerase δ, η, and ζ activities. Proc. Natl. Acad. Sci. USA 2013, 110, 7672–7677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sneeden, J.L.; Grossi, S.M.; Tappin, I.; Hurwitz, J.; Heyer, W.-D. Reconstitution of recombination-associated DNA synthesis with human proteins. Nucleic Acids Res. 2013, 41, 4913–4925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolinjivadi, A.M.; Sannino, V.; De Antoni, A.; Zadorozhny, K.; Kilkenny, M.; Técher, H.; Baldi, G.; Shen, R.; Ciccia, A.; Pellegrini, L.; et al. Smarcal1-mediated fork reversal triggers Mre11-dependent degradation of nascent DNA in the absence of Brca2 and stable Rad51 Nucleofilaments. Mol. Cell 2017, 67, 867–881.e7. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Ira, G.; Tercero, J.A.; Holmes, A.M.; Diffley, J.F.X.; Haber, J.E. Role of DNA Replication Proteins in Double-Strand Break-Induced Recombination in Saccharomyces cerevisiae. Mol. Cell. Biol. 2004, 24, 6891–6899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganai, R.A.; Zhang, X.-P.; Heyer, W.-D.; Johansson, E. Strand displacement synthesis by yeast DNA polymerase ε. Nucleic Acids Res. 2016, 44, 8229–8240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raia, P.; Delarue, M.; Sauguet, L. An updated structural classification of replicative DNA polymerases. Biochem. Soc. Trans. 2019, 47, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Cooper, C.D. Archaeal DNA polymerases: New frontiers in DNA replication and repair. Emerg. Top. Life Sci. 2018, 2, 503–516. [Google Scholar] [CrossRef] [Green Version]
- Cann, I.K.O.; Ishino, S.; Hayashi, I.; Komori, K.; Toh, H.; Morikawa, K.; Ishino, Y. Functional Interactions of a Homolog of Proliferating Cell Nuclear Antigen with DNA Polymerases inArchaea. J. Bacteriol. 1999, 181, 6591–6599. [Google Scholar] [CrossRef] [Green Version]
- Gouge, J.; Ralec, C.; Henneke, G.; Delarue, M. molecular recognition of canonical and deaminated bases by P. abyssi family B DNA Polymerase. J. Mol. Biol. 2012, 423, 315–336. [Google Scholar] [CrossRef] [PubMed]
- Uemori, T.; Sato, Y.; Kato, I.; Doi, H.; Ishino, Y. A novel DNA polymerase in the hyperthermophilic archaeon, Pyrococcus furiosus: Gene cloning, expression, and characterization. Genes Cells 1997, 2, 499–512. [Google Scholar] [CrossRef]
- Raia, P.; Carroni, M.; Henry, E.; Pehau-Arnaudet, G.; Brûlé, S.; Béguin, P.; Henneke, G.; Lindahl, E.; Delarue, M.; Sauguet, L. Structure of the DP1–DP2 PolD complex bound with DNA and its implications for the evolutionary history of DNA and RNA polymerases. PLoS Biol. 2019, 17, e3000122. [Google Scholar] [CrossRef] [PubMed]
- Boudsocq, F. Sulfolobus solfataricus P2 DNA polymerase IV (Dpo4): An archaeal DinB-like DNA polymerase with lesion-bypass properties akin to eukaryotic poleta. Nucleic Acids Res. 2001, 29, 4607–4616. [Google Scholar] [CrossRef] [Green Version]
- Le Breton, M.; Henneke, G.; Norais, C.; Flament, D.; Myllykallio, H.; Querellou, J.; Raffin, J.-P. The Heterodimeric Primase from the Euryarchaeon Pyrococcus abyssi: A Multifunctional Enzyme for Initiation and Repair? J. Mol. Biol. 2007, 374, 1172–1185. [Google Scholar] [CrossRef] [Green Version]
- Jozwiakowski, S.K.; Keith, B.J.; Gilroy, L.; Doherty, A.J.; Connolly, B.A. An archaeal family-B DNA polymerase variant able to replicate past DNA damage: Occurrence of replicative and translesion synthesis polymerases within the B family. Nucleic Acids Res. 2014, 42, 9949–9963. [Google Scholar] [CrossRef] [Green Version]
- Lemor, M.; Kong, Z.; Henry, E.; Brizard, R.; Laurent, S.; Bossé, A.; Henneke, G. Differential activities of DNA Polymerases in processing Ribonucleotides during DNA synthesis in Archaea. J. Mol. Biol. 2018, 430, 4908–4924. [Google Scholar] [CrossRef]
- Killelea, T.; Palud, A.; Akcha, F.; Lemor, M.; L’Haridon, S.; Godfroy, A.; Henneke, G. The interplay at the replisome mitigates the impact of oxidative damage on the genetic integrity of hyperthermophilic Archaea. Elife 2019, 8, e45320. [Google Scholar] [CrossRef]
- Gueguen, Y.; Rolland, J.-L.; Lecompte, O.; Azam, P.; Le Romancer, G.; Flament, D.; Raffin, J.-P.; Dietrich, J. Characterization of two DNA polymerases from the hyperthermophilic euryarchaeonPyrococcus abyssi. Eur. J. Biochem. 2001, 268, 5961–5969. [Google Scholar] [CrossRef] [PubMed]
- Rouillon, C.; Henneke, G.; Flament, D.; Querellou, J.; Raffin, J.-P. DNA polymerase switching on Homotrimeric PCNA at the replication fork of the Euryarchaea Pyrococcus abyssi. J. Mol. Biol. 2007, 369, 343–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henneke, G.; Flament, D.; Hübscher, U.; Querellou, J.; Raffin, J.-P. The Hyperthermophilic Euryarchaeota Pyrococcus abyssi Likely Requires the Two DNA Polymerases D and B for DNA Replication. J. Mol. Biol. 2005, 350, 53–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palud, A.; Villani, G.; L’Haridon, S.; Querellou, J.; Henneke, G.; Raffin, J.-P. Intrinsic properties of the two replicative DNA polymerases ofPyrococcus abyssiin replicating abasic sites: Possible role in DNA damage tolerance? Mol. Microbiol. 2008, 70, 746–761. [Google Scholar] [CrossRef] [Green Version]
- Ralec, C.; Henry, E.; Lemor, M.; Killelea, T.; Henneke, G. Calcium-driven DNA synthesis by a high-fidelity DNA polymerase. Nucleic Acids Res. 2017, 45, 12425–12440. [Google Scholar] [CrossRef] [Green Version]
- Greenough, L.; Kelman, Z.; Gardner, A.F. The roles of family B and D DNA polymerases in Thermococcus species 9° N Okazaki fragment maturation. J. Biol. Chem. 2015, 290, 12514–12522. [Google Scholar] [CrossRef] [Green Version]
- Henneke, G. In vitro reconstitution of RNA primer removal in Archaea reveals the existence of two pathways. Biochem. J. 2012, 447, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Ishino, S.; Ishino, Y. Comprehensive Search for DNA Polymerase in the Hyperthermophilic Archaeon, Pyrococcus furiosus. Nucleosides Nucleotides Nucleic Acids 2006, 25, 681–691. [Google Scholar] [CrossRef]
- Kushida, T.; Narumi, I.; Ishino, S.; Ishino, Y.; Fujiwara, S.; Imanaka, T.; Higashibata, H. PolB, a Family B DNA Polymerase, in Thermococcus kodakarensis is Important for DNA Repair, but not DNA Replication. Microbes Environ. 2019, 34, 316–326. [Google Scholar] [CrossRef] [Green Version]
- Sarmiento, F.; Mrázek, J.; Whitman, W.B. Genome-scale analysis of gene function in the hydrogenotrophic methanogenic archaeon Methanococcus maripaludis. Proc. Natl. Acad. Sci. USA 2013, 110, 4726–4731. [Google Scholar] [CrossRef] [Green Version]
- Čuboňová, L.; Richardson, T.; Burkhart, B.W.; Kelman, Z.; Connolly, B.A.; Reeve, J.N.; Santangelo, T.J. Archaeal DNA Polymerase D but Not DNA Polymerase B Is Required for Genome Replication in Thermococcus kodakarensis. J. Bacteriol. 2013, 195, 2322–2328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raynard, S.; Sung, P. Assay for human Rad51-mediated DNA displacement loop formation. Cold Spring Harb. Protoc. 2009, 2009, 5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. ISBN 978-1-58829-343-5. [Google Scholar]
- Meslet-Cladière, L.; Norais, C.; Kuhn, J.; Briffotaux, J.; Sloostra, J.W.; Ferrari, E.; Hübscher, U.; Flament, D.; Myllykallio, H. A Novel Proteomic Approach Identifies New Interaction Partners for Proliferating Cell Nuclear Antigen. J. Mol. Biol. 2007, 372, 1137–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauguet, L.; Raia, P.; Henneke, G.; Delarue, M. Shared active site architecture between archaeal PolD and multi-subunit RNA polymerases revealed by X-ray crystallography. Nat. Commun. 2016, 7, 12227. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; McCorvie, T.J.; Yates, L.A.; Zhang, X. Structural basis of homologous recombination. Cell. Mol. Life Sci. 2019, 77, 3–18. [Google Scholar] [CrossRef] [Green Version]
- Seitz, E.M.; Brockman, J.P.; Sandler, S.J.; Clark, A.J.; Kowalczykowski, S.C. RadA protein is an archaeal RecA protein homolog that catalyzes DNA strand exchange. Genes Dev. 1998, 12, 1248–1253. [Google Scholar] [CrossRef]
- Del Val, E.; Nasser, W.; Abaibou, H.; Reverchon, S. RecA and DNA recombination: A review of molecular mechanisms. Biochem. Soc. Trans. 2019, 47, 1511–1531. [Google Scholar] [CrossRef]
- Atwell, S.X.; Migliozzi, D.; Dupont, A.; Viovy, J.-L.; Cappello, G. Structural transitions and mechanochemical coupling in the nucleoprotein filament explain homology selectivity and Rad51 protein cooperativity in cellular DNA repair. Phys. Rev. E 2020, 101, 032407. [Google Scholar] [CrossRef] [PubMed]
- De Vlaminck, I.; Van Loenhout, M.T.; Zweifel, L.; Blanken, J.D.; Hooning, K.; Hage, S.; Kerssemakers, J.; Dekker, C. Mechanism of Homology Recognition in DNA Recombination from Dual-Molecule Experiments. Mol. Cell 2012, 46, 616–624. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Sneeden, J.; Heyer, W.-D. In vitro assays for DNA pairing and recombination-associated DNA synthesis. In DNA Recombination; Humana Press: Totowa, NJ, USA, 2011; Volume 745, pp. 363–383. [Google Scholar]
- Ito, K.; Murayama, Y.; Takahashi, M.; Iwasaki, H. Two three-strand intermediates are processed during Rad51-driven DNA strand exchange. Nat. Struct. Mol. Biol. 2017, 25, 29–36. [Google Scholar] [CrossRef]
- Sheng, D.; Zhu, S.; Wei, T.; Ni, J.; Shen, Y. The in vitro activity of a Rad55 homologue from Sulfolobus tokodaii, a candidate mediator in RadA-catalyzed homologous recombination. Extremophiles 2007, 12, 147–157. [Google Scholar] [CrossRef]
- Graham, W.J.; Rolfsmeier, M.L.; Haseltine, C. An archaeal RadA paralog influences presynaptic filament formation. DNA Repair 2013, 12, 403–413. [Google Scholar] [CrossRef] [Green Version]
- Rolfsmeier, M.L.; Haseltine, C. The RadA Recombinase and Paralogs of the Hyperthermophilic Archaeon Sulfolobus solfataricus. Methods Enzymol. 2018, 600, 255–284. [Google Scholar] [CrossRef] [PubMed]
- Tavares, E.M.; Wright, W.D.; Heyer, W.-D.; Le Cam, E.; Dupaigne, P. In vitro role of Rad54 in Rad51-ssDNA filament-dependent homology search and synaptic complexes formation. Nat. Commun. 2019, 10, 4058. [Google Scholar] [CrossRef] [Green Version]
- Knadler, C.; Rolfsmeier, M.; Vallejo, A.; Haseltine, C. Characterization of an archaeal recombinase paralog that exhibits novel anti-recombinase activity. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2020, 821, 111703. [Google Scholar] [CrossRef] [PubMed]
- Wardell, K.; Haldenby, S.; Jones, N.; Liddell, S.; Ngo, G.; Allers, T. RadB acts in homologous recombination in the archaeon Haloferax volcanii, consistent with a role as recombination mediator. DNA Repair 2017, 55, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Andriuskevicius, T.; Kotenko, O.; Makovets, S. Putting together and taking apart: Assembly and disassembly of the Rad51 nucleoprotein filament in DNA repair and genome stability. Cell Stress 2018, 2, 96–112. [Google Scholar] [CrossRef] [PubMed]
- Hogrel, G.; Lu, Y.; Laurent, S.; Henry, E.; Etienne, C.; Phung, D.K.; Dulermo, R.; Bossé, A.; Pluchon, P.-F.; Clouet-D’Orval, B.; et al. Physical and functional interplay between PCNA DNA clamp and Mre11–Rad50 complex from the archaeon Pyrococcus furiosus. Nucleic Acids Res. 2018, 46, 5651–5663. [Google Scholar] [CrossRef]
- Formosa, T.; Alberts, B.M. DNA synthesis dependent on genetic recombination: Characterization of a reaction catalyzed by purified bacteriophage T4 proteins. Cell 1986, 47, 793–806. [Google Scholar] [CrossRef]
- Patel, M.; Jiang, Q.; Woodgate, R.; Cox, M.M.; Goodman, M.F. A new model for SOS-induced mutagenesis: How RecA protein activates DNA polymerase V. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 171–184. [Google Scholar] [CrossRef] [Green Version]
- Madru, C.; Henneke, G.; Raia, P.; Hugonneau-Beaufet, I.; Pehau-Arnaudet, G.; England, P.; Lindahl, E.; Delarue, M.; Carroni, M.; Sauguet, L. Structural basis for the increased processivity of D-family DNA polymerases in complex with PCNA. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreuzer, K.N.; Brister, J.R. Initiation of bacteriophage T4 DNA replication and replication fork dynamics: A review in the Virology Journal series on bacteriophage T4 and its relatives. Virol. J. 2010, 7, 358. [Google Scholar] [CrossRef] [PubMed] [Green Version]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hogrel, G.; Lu, Y.; Alexandre, N.; Bossé, A.; Dulermo, R.; Ishino, S.; Ishino, Y.; Flament, D. Role of RadA and DNA Polymerases in Recombination-Associated DNA Synthesis in Hyperthermophilic Archaea. Biomolecules 2020, 10, 1045. https://doi.org/10.3390/biom10071045
Hogrel G, Lu Y, Alexandre N, Bossé A, Dulermo R, Ishino S, Ishino Y, Flament D. Role of RadA and DNA Polymerases in Recombination-Associated DNA Synthesis in Hyperthermophilic Archaea. Biomolecules. 2020; 10(7):1045. https://doi.org/10.3390/biom10071045
Chicago/Turabian StyleHogrel, Gaëlle, Yang Lu, Nicolas Alexandre, Audrey Bossé, Rémi Dulermo, Sonoko Ishino, Yoshizumi Ishino, and Didier Flament. 2020. "Role of RadA and DNA Polymerases in Recombination-Associated DNA Synthesis in Hyperthermophilic Archaea" Biomolecules 10, no. 7: 1045. https://doi.org/10.3390/biom10071045