Novel Barbiturate-Nitrate Compounds Inhibit the Upregulation of Matrix Metalloproteinase-9 Gene Expression in Intestinal Inflammation through a cGMP-Mediated Pathway


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Cell Culture
2.2. Gelatin Zymography
2.3. Quantitative PCR
2.4. Nitrate and Nitrite Quantification–Modified Griess Assay
2.5. NF-κB (p65) Binding Activity
2.6. Statistical Analysis
3. Results
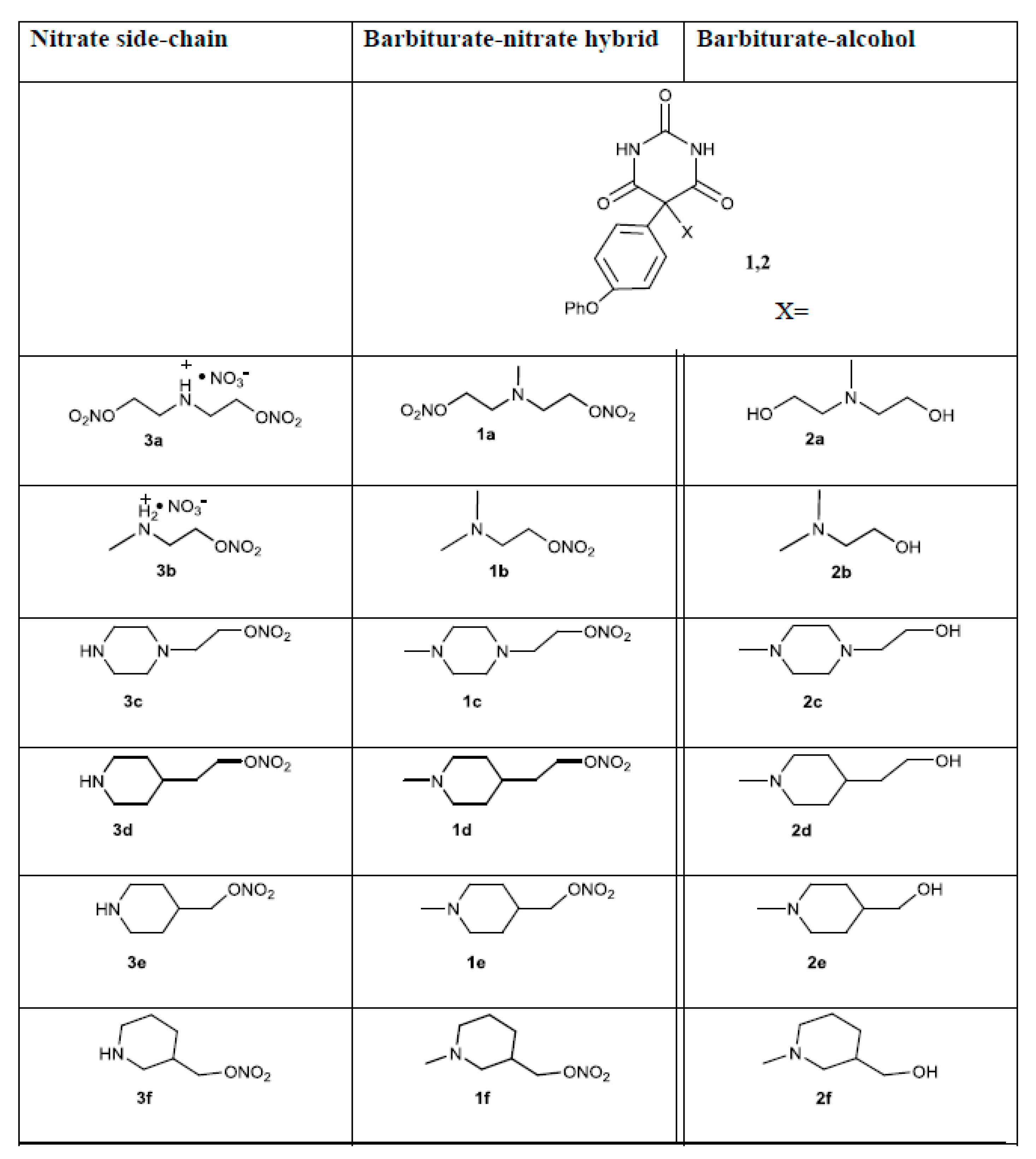
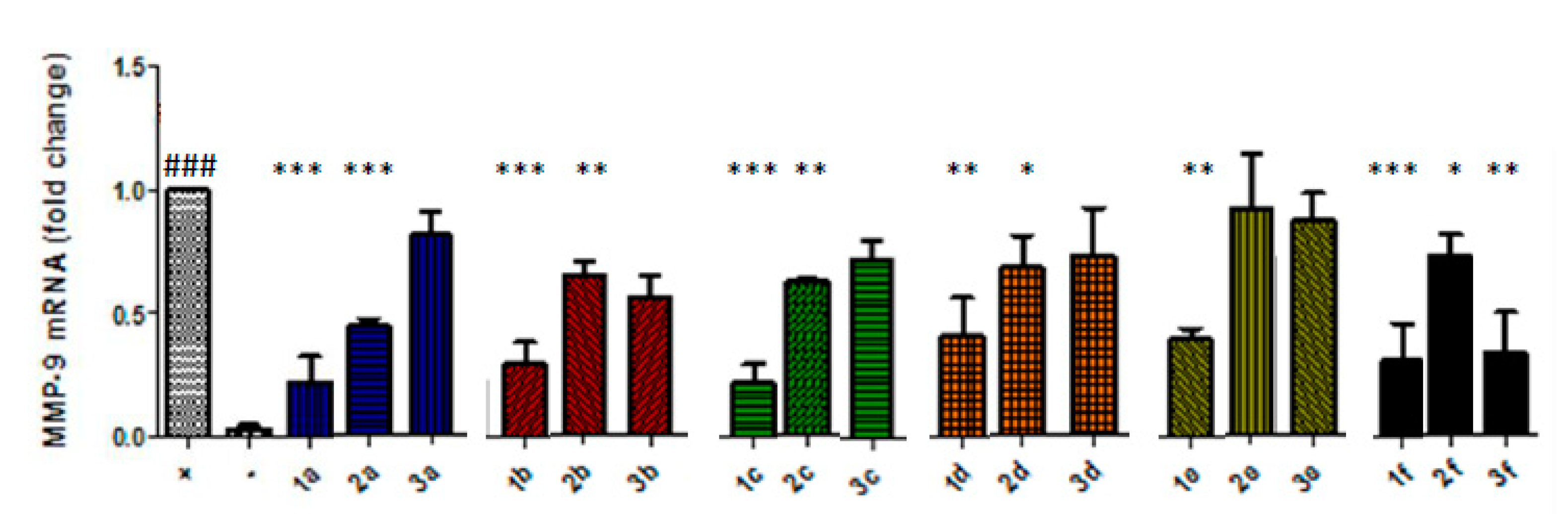
3.1. Barbiturate-Nitrate Hybrids Reduce MMP-9 Expression in Cytokine-Stimulated Caco-2 Cells
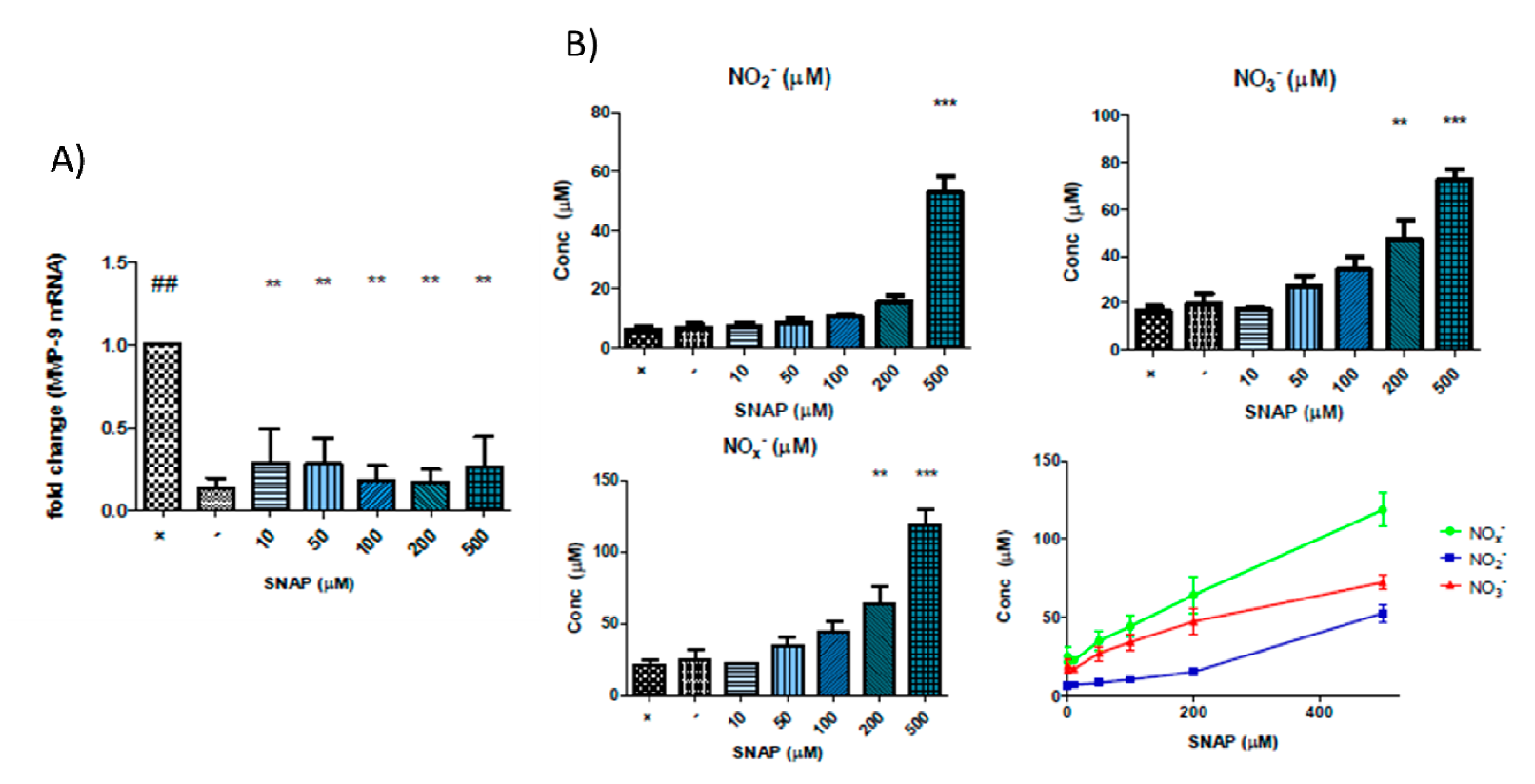
3.2. Nitric Oxide Donors Reduce MMP-9 mRNA Levels in Cytokine-Stimulated Caco-2 Cells
3.3. Barbiturate-Nitrate-Hybrids Inhibit MMP-9 Gene Expression in an NF-κB-Independent Manner
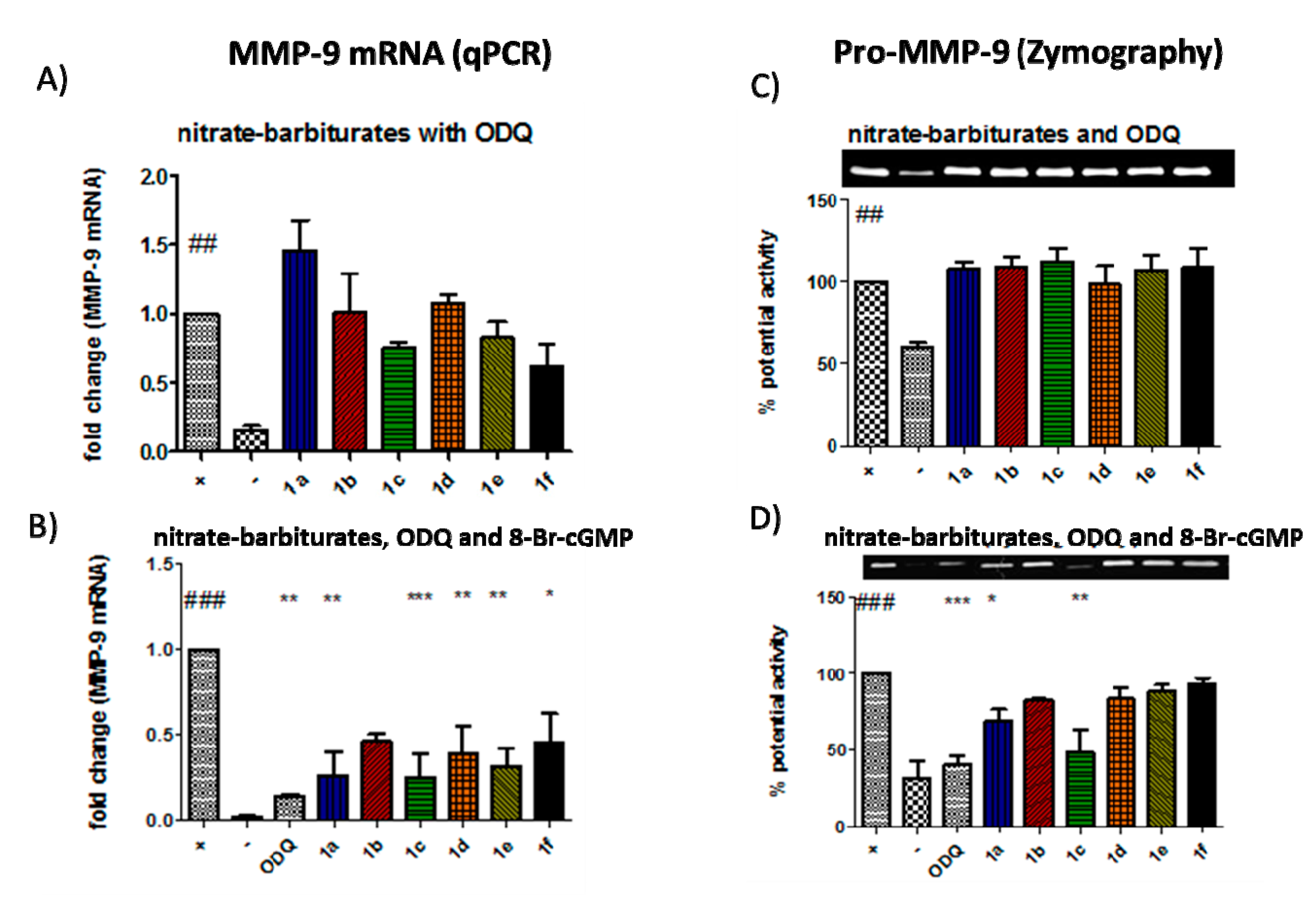
3.4. Inhibition of MMP-9 by the Barbiturate-Nitrate Hybrids Is Partly Mediated Through a sGC/cGMP Pathway
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Vandooren, J.; Steen, P.E.V.D.; Opdenakker, G. Biochemistry and molecular biology of gelatinase B or matrix metalloproteinase-9 (MMP-9): The next decade. Crit. Rev. Biochem. Mol. Biol. 2013, 48, 222–272. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, D.; Morrison, C.J.; Overall, C.M. Matrix metalloproteinases: What do they not do? New substrates and biological roles identified by murine models and proteomics. Biochim. Biophys. Acta 2010, 1803, 39–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehana, E.-S.E.; Khafaga, A.F.; El-Blehi, S.S.; El-Sayed, M.E.; Khafaga, A.F.; Samar, E.-B.S. The role of matrix metalloproteinases in osteoarthritis pathogenesis: An updated review. Life Sci. 2019, 234, 116786. [Google Scholar] [CrossRef] [PubMed]
- Medina, C.; Videla, S.; Radomski, A.; Radomski, M.W.; Antolín, M.; Guarner, F.; Vilaseca, J.; Salas, A.; Malagelada, J.-R. Increased activity and expression of matrix metalloproteinase-9 in a rat model of distal colitis. Am. J. Physiol. Liver Physiol. 2003, 284, G116–G122. [Google Scholar] [CrossRef]
- Medina, C.; Santana, A.; Paz, M.C.; Díaz-González, F.; Farre, E.; Salas, A.; Radomski, M.W.; Quintero, E. Matrix metalloproteinase-9 modulates intestinal injury in rats with transmural colitis. J. Leukoc. Biol. 2006, 79, 954–962. [Google Scholar] [CrossRef] [Green Version]
- Castaneda, F.E.; Walia, B.; Vijay–Kumar, M.; Patel, N.R.; Roser, S.; Kolachala, V.L.; Rojas, M.; Wang, L.; Oprea, G.; Garg, P.; et al. Targeted Deletion of Metalloproteinase 9 Attenuates Experimental Colitis in Mice: Central Role of Epithelial-Derived MMP. Gastroenterology 2005, 129, 1991–2008. [Google Scholar] [CrossRef]
- Al-Sadi, R.; Youssef, M.; Rawat, M.; Guo, S.; Dokladny, K.; Haque, M.; Watterson, M.D.; Ma, T.Y.; Watterson, D.M. MMP-9-induced increase in intestinal epithelial tight permeability is mediated by p38 kinase signaling pathway activation of MLCK gene. Am. J. Physiol. Liver Physiol. 2019, 316, G278–G290. [Google Scholar] [CrossRef]
- Opdenakker, G.; Steen, P.E.V.D.; Van Damme, J. Gelatinase B: A tuner and amplifier of immune functions. Trends Immunol. 2001, 22, 571–579. [Google Scholar] [CrossRef]
- Atkinson, J.J.; Senior, R.M. Matrix Metalloproteinase-9 in Lung Remodeling. Am. J. Respir. Cell Mol. Biol. 2003, 28, 12–24. [Google Scholar] [CrossRef]
- Mohan, R.; Chintala, S.K.; Jung, J.-C.; Villar, W.V.L.; McCabe, F.; Russo, L.A.; Lee, Y.; McCarthy, B.E.; Wollenberg, K.R.; Jester, J.V.; et al. Matrix Metalloproteinase Gelatinase B (MMP-9) Coordinates and Effects Epithelial Regeneration. J. Biol. Chem. 2001, 277, 2065–2072. [Google Scholar] [CrossRef] [Green Version]
- Fini, M.E.; Parks, W.C.; Rinehart, W.B.; Girard, M.T.; Matsubara, M.; Cook, J.R.; West-Mays, J.A.; Sadow, P.M.; Burgeson, R.E.; Jeffrey, J.J.; et al. Role of matrix metalloproteinases in failure to re-epithelialize after corneal injury. Am. J. Pathol. 1996, 149, 1287–1302. [Google Scholar] [PubMed]
- Hu, J.; Steen, P.E.V.D.; Sang, Q.-X.A.; Opdenakker, G. Matrix metalloproteinase inhibitors as therapy for inflammatory and vascular diseases. Nat. Rev. Drug Discov. 2007, 6, 480–498. [Google Scholar] [CrossRef] [PubMed]
- Winer, A.; Adams, S.; Mignatti, P. Matrix Metalloproteinase Inhibitors in Cancer Therapy: Turning Past Failures Into Future Successes. Mol. Cancer Ther. 2018, 17, 1147–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grams, F.; Brandstetter, H.; Dalò, S.; Geppert, D.; Krell, H.-W.; Leinert, H.; Livi, V.; Menta, E.; Oliva, A.; Zimmermann, G. Pyrimidine-2,4,6-Triones: A New Effective and Selective Class of Matrix Metalloproteinase Inhibitors. Biol. Chem. 2001, 382, 1277–1285. [Google Scholar] [CrossRef] [PubMed]
- Breyholz, H.-J.; Schäfers, M.; Wagner, S.; Höltke, C.; Faust, A.; Rabeneck, H.; Levkau, B.; Schober, O.; Kopka, K. C-5-Disubstituted Barbiturates as Potential Molecular Probes for Noninvasive Matrix Metalloproteinase Imaging. J. Med. Chem. 2005, 48, 3400–3409. [Google Scholar] [CrossRef] [PubMed]
- Foley, L.H.; Palermo, R.; Dunten, P.; Wang, P. Novel 5,5-disubstitutedpyrimidine-2,4,6-triones as selective MMP inhibitors. Bioorganic Med. Chem. Lett. 2001, 11, 969–972. [Google Scholar] [CrossRef]
- Wang, J.; O’Sullivan, S.; Harmon, S.; Keaveny, R.; Radomski, M.W.; Medina, C.; Gilmer, J.F. Design of Barbiturate–Nitrate Hybrids that Inhibit MMP-9 Activity and Secretion. J. Med. Chem. 2012, 55, 2154–2162. [Google Scholar] [CrossRef]
- O’Sullivan, S.; Medina, C.; Ledwidge, M.; Radomski, M.W.; Gilmer, J.F. Nitric oxide-matrix metaloproteinase-9 interactions: Biological and pharmacological significance. Biochim. Biophys. Acta 2014, 1843, 603–617. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, S.; Wang, J.; Pigott, M.T.; Docherty, N.; Boyle, N.; Lis, S.K.; Gilmer, J.F.; Medina, C. Inhibition of matrix metalloproteinase-9 by a barbiturate-nitrate hybrid ameliorates dextran sulphate sodium-induced colitis: Effect on inflammation-related genes. Br. J. Pharmacol. 2017, 174, 512–524. [Google Scholar] [CrossRef] [Green Version]
- Webb, D.J.; Megson, I. Nitric oxide donor drugs: Current status and future trends. Expert Opin. Investig. Drugs 2002, 11, 587–601. [Google Scholar] [CrossRef]
- Schrammel, A.; Behrends, S.; Schmidt, K.; Koesling, D.; Mayer, B. Characterization of 1H-(1,2,4)oxadiazolo(4,3-a)quinoxalin-1-one as a heme-site inhibitor of nitric oxide-sensitive guanylyl cyclase. Mol. Pharmacol. 1996, 50, 1–5. [Google Scholar] [PubMed]
- Garthwaite, J.; Southam, E.; Boulton, C.L.; Nielsen, E.B.; Schmidt, K.; Mayer, B. Potent and selective inhibition of nitric oxide-sensitive guanylyl cyclase by 1H-(1,2,4)oxadiazolo(4,3-a)quinoxalin-1-one. Mol. Pharmacol. 1995, 48, 184–188. [Google Scholar] [PubMed]
- Martin, M.D.; Matrisian, L.M. The other side of MMPs: Protective roles in tumor progression. Cancer Metastasis Rev. 2007, 26, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Juurikka, K.; Butler, G.S.; Salo, T.; Nyberg, P.; Åström, P. The Role of MMP8 in Cancer: A Systematic Review. Int. J. Mol. Sci. 2019, 20, 4506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dufour, A.; Overall, C.M. Missing the target: Matrix metalloproteinase antitargets in inflammation and cancer. Trends Pharmacol. Sci. 2013, 34, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Kobayashi, T.; D’Alessandro-Gabazza, C.N.; Toda, M.; Fujiwara, K.; Okano, T.; Fujimoto, H.; Asayama, K.; Takeshita, A.; Yasuma, T.; et al. Protective Role of Matrix Metalloproteinase-2 in Allergic Bronchial Asthma. Front. Immunol. 2019, 10, 1795. [Google Scholar] [CrossRef] [Green Version]
- Gong, Y.; Hart, E.; Shchurin, A.; Hoover-Plow, J. Inflammatory macrophage migration requires MMP-9 activation by plasminogen in mice. J. Clin. Investig. 2008, 118, 3012–3024. [Google Scholar] [CrossRef] [Green Version]
- Fayard, B.; Bianchi, F.; Dey, J.; Moreno, E.; Djaffer, S.; E Hynes, N.; Monard, D. The Serine Protease Inhibitor Protease Nexin-1 Controls Mammary Cancer Metastasis through LRP-1-Mediated MMP-9 Expression. Cancer Res. 2009, 69, 5690–5698. [Google Scholar] [CrossRef] [Green Version]
- Dormán, G.; Kocsis-Szommer, K.; Spadoni, C.; Ferdinandy, P. MMP Inhibitors in Cardiac Diseases: An Update. Recent Patents Cardiovasc. Drug Discov. 2007, 2, 186–194. [Google Scholar] [CrossRef]
- Garcia, R.A.; Pantazatos, D.P.; Gessner, C.R.; Go, K.V.; Woods, V.L.; Villarreal, F.J. Molecular Interactions between Matrilysin and the Matrix Metalloproteinase Inhibitor Doxycycline Investigated by Deuterium Exchange Mass Spectrometry. Mol. Pharmacol. 2005, 67, 1128–1136. [Google Scholar] [CrossRef]
- Egeblad, M.; Werb, Z. New functions for the matrix metalloproteinases in cancer progression. Nat. Rev. Cancer 2002, 2, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Ramamurthy, N.S.; Rifkin, B.R.; Greenwald, R.A.; Xu, J.-W.; Liu, Y.; Turner, G.; Golub, L.M.; Vernillo, A. Inhibition of Matrix Metalloproteinase-Mediated Periodontal Bone Loss in Rats: A Comparison of 6 Chemically Modified Tetracyclines. J. Periodontol. 2002, 73, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Paemen, L.; Martens, E.; Norga, K.; Masure, S.; Roets, E.; Hoogmartens, J.; Opdenakker, G. The gelatinase inhibitory activity of tetracyclines and chemically modified tetracycline analogues as measured by a novel microtiter assay for inhibitors. Biochem. Pharmacol. 1996, 52, 105–111. [Google Scholar] [CrossRef]
- Sternlicht, M.D.; Werb, Z. How Matrix Metalloproteinases Regulate Cell Behavior. Annu. Rev. Cell Dev. Biol. 2001, 17, 463–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.-D.; Wang, W. Protective effect of ilomastat on trinitrobenzenesulfonic acid-induced ulcerative colitis in rats. World J. Gastroenterol. 2008, 14, 5683–5688. [Google Scholar] [CrossRef] [PubMed]
- St-Pierre, Y.; Couillard, J.; Van Themsche, C. Regulation of MMP-9 gene expression for the development of novel molecular targets against cancer and inflammatory diseases. Expert Opin. Ther. Targets 2004, 8, 473–489. [Google Scholar] [CrossRef]
- Eberhardt, W.; Huwiler, A.; Beck, K.F.; Walpen, S.; Pfeilschifter, J. Amplification of IL-1 beta-induced matrix metalloproteinase-9 expression by superoxide in rat glomerular mesangial cells is mediated by increased activities of NF-kappa B and activating protein-1 and involves activation of the mitogen-activated protein kinase pathways. J. Immunol. 2000, 165, 5788–5797. [Google Scholar]
- Bond, M.; Fabunmi, R.P.; Baker, A.H.; Newby, A. Synergistic upregulation of metalloproteinase-9 by growth factors and inflammatory cytokines: An absolute requirement for transcription factor NF-kappa B. FEBS Lett. 1998, 435, 435. [Google Scholar] [CrossRef] [Green Version]
- Bond, M.; Chase, A.J.; Baker, A.H.; Newby, A. Inhibition of transcription factor NF-kappaB reduces matrix metalloproteinase-1, -3 and -9 production by vascular smooth muscle cells. Cardiovasc. Res. 2001, 50, 556–565. [Google Scholar] [CrossRef]
- Chen, X.; Chang, L.; Li, X.; Huang, J.; Yang, L.; Lai, X.; Huang, Z.; Wang, Z.; Wu, X.; Zhao, J.; et al. Tc17/IL-17A Up-Regulated the Expression of MMP-9 via NF-κB Pathway in Nasal Epithelial Cells of Patients with Chronic Rhinosinusitis. Front. Immunol. 2018, 9, 2121. [Google Scholar] [CrossRef] [Green Version]
- Janssen-Heininger, Y.M.W.; E Poynter, M.; A Baeuerle, P. Recent advances towards understanding redox mechanisms in the activation of nuclear factor kappaB. Free. Radic. Biol. Med. 2000, 28, 1317–1327. [Google Scholar] [CrossRef]
- Dijkstra, G.; Moshage, H.; Jansen, P.L.M. Blockade of NF-κ B Activation and Donation of Nitric Oxide: New Treatment Options in Inflammatory Bowel Disease? Scand. J. Gastroenterol. 2002, 37, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Pfeilschifter, J.; Eberhardt, W.; Beck, K.-F. Regulation of gene expression by nitric oxide. Pflüger 2001, 442, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Ahern, G. cGMP and S-nitrosylation: Two routes for modulation of neuronal excitability by NO. Trends Neurosci. 2002, 25, 510–517. [Google Scholar] [CrossRef]
- Gudi, T.; Hong, G.K.-P.; Vaandrager, A.B.; Lohmann, S.M.; Pilz, R.B. Nitric oxide and cGMP regulate gene expression in neuronal and glial cells by activating type II cGMP-dependent protein kinase. FASEB J. 1999, 13, 2143–2152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huvar, I. Regulation of Gene Expression by cGMP-dependent Protein Kinase. J. Biol. Chem. 1996, 271, 4597–4600. [Google Scholar] [CrossRef] [Green Version]
- Gudi, T.; Lohmann, S.M.; Pilz, R.B. Regulation of gene expression by cyclic GMP-dependent protein kinase requires nuclear translocation of the kinase: Identification of a nuclear localization signal. Mol. Cell. Biol. 1997, 17, 5244–5254. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
O’Sullivan, S.; Wang, J.; Radomski, M.W.; Gilmer, J.F.; Medina, C. Novel Barbiturate-Nitrate Compounds Inhibit the Upregulation of Matrix Metalloproteinase-9 Gene Expression in Intestinal Inflammation through a cGMP-Mediated Pathway. Biomolecules 2020, 10, 808. https://doi.org/10.3390/biom10050808
O’Sullivan S, Wang J, Radomski MW, Gilmer JF, Medina C. Novel Barbiturate-Nitrate Compounds Inhibit the Upregulation of Matrix Metalloproteinase-9 Gene Expression in Intestinal Inflammation through a cGMP-Mediated Pathway. Biomolecules. 2020; 10(5):808. https://doi.org/10.3390/biom10050808
Chicago/Turabian StyleO’Sullivan, Shane, Jun Wang, Marek W. Radomski, John F. Gilmer, and Carlos Medina. 2020. "Novel Barbiturate-Nitrate Compounds Inhibit the Upregulation of Matrix Metalloproteinase-9 Gene Expression in Intestinal Inflammation through a cGMP-Mediated Pathway" Biomolecules 10, no. 5: 808. https://doi.org/10.3390/biom10050808