Different Impacts of MucR Binding to the babR and virB Promoters on Gene Expression in Brucella abortus 2308

 ,
, 
{kind=link}
{kind=link}
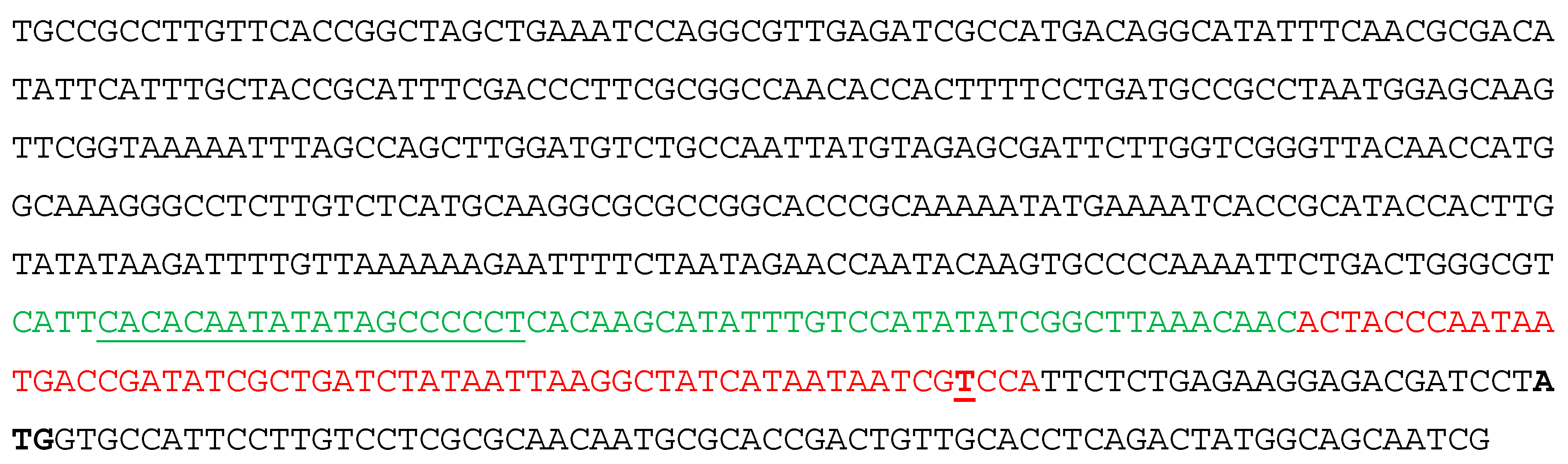{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cloning, Protein Expression and Purification
2.2. Electrophoretic Mobility Shift Assay
2.3. RT-qPCR
2.4. Isolation of B. abortus Field Strains and Genomic Sequencing
3. Results
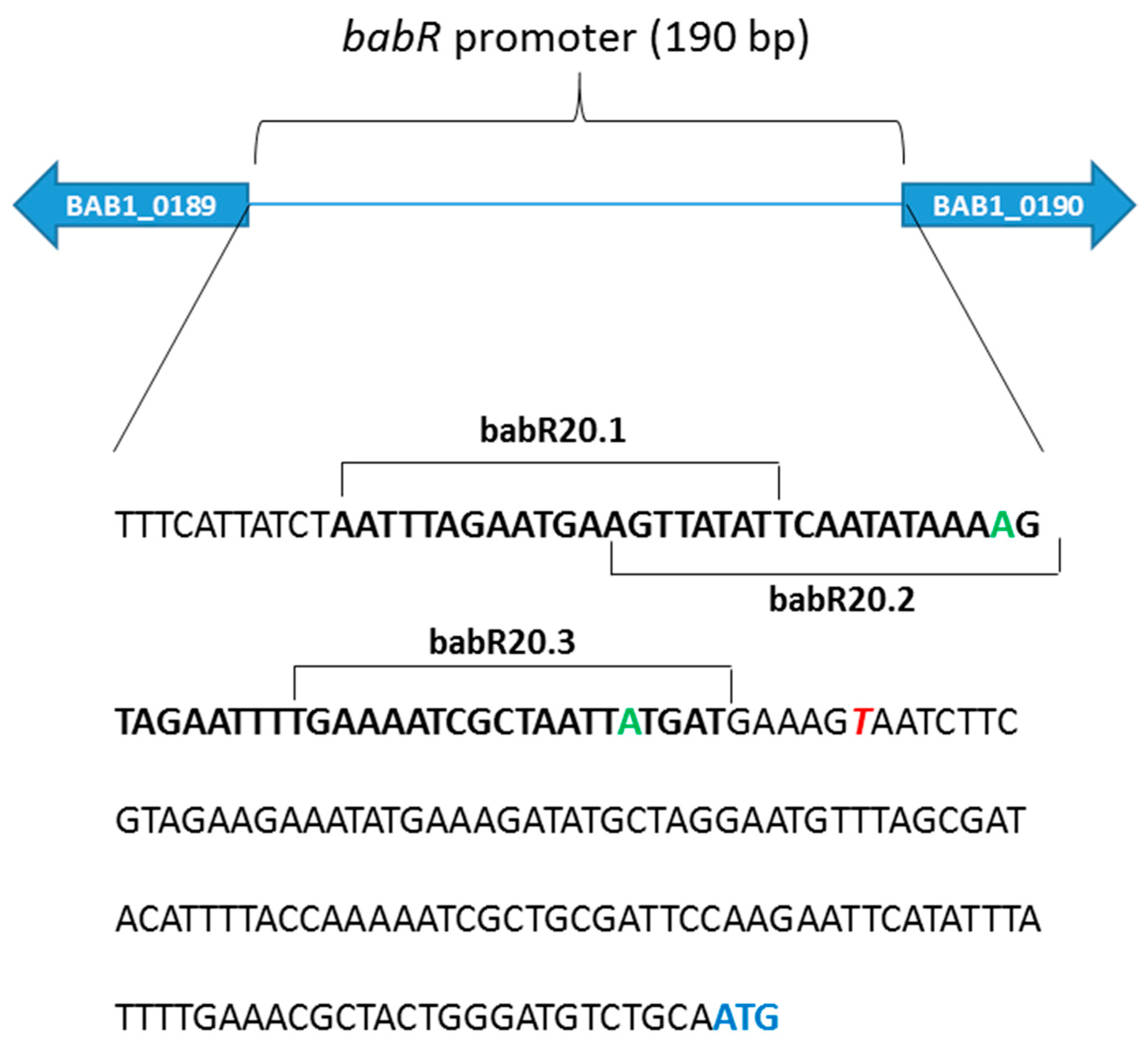
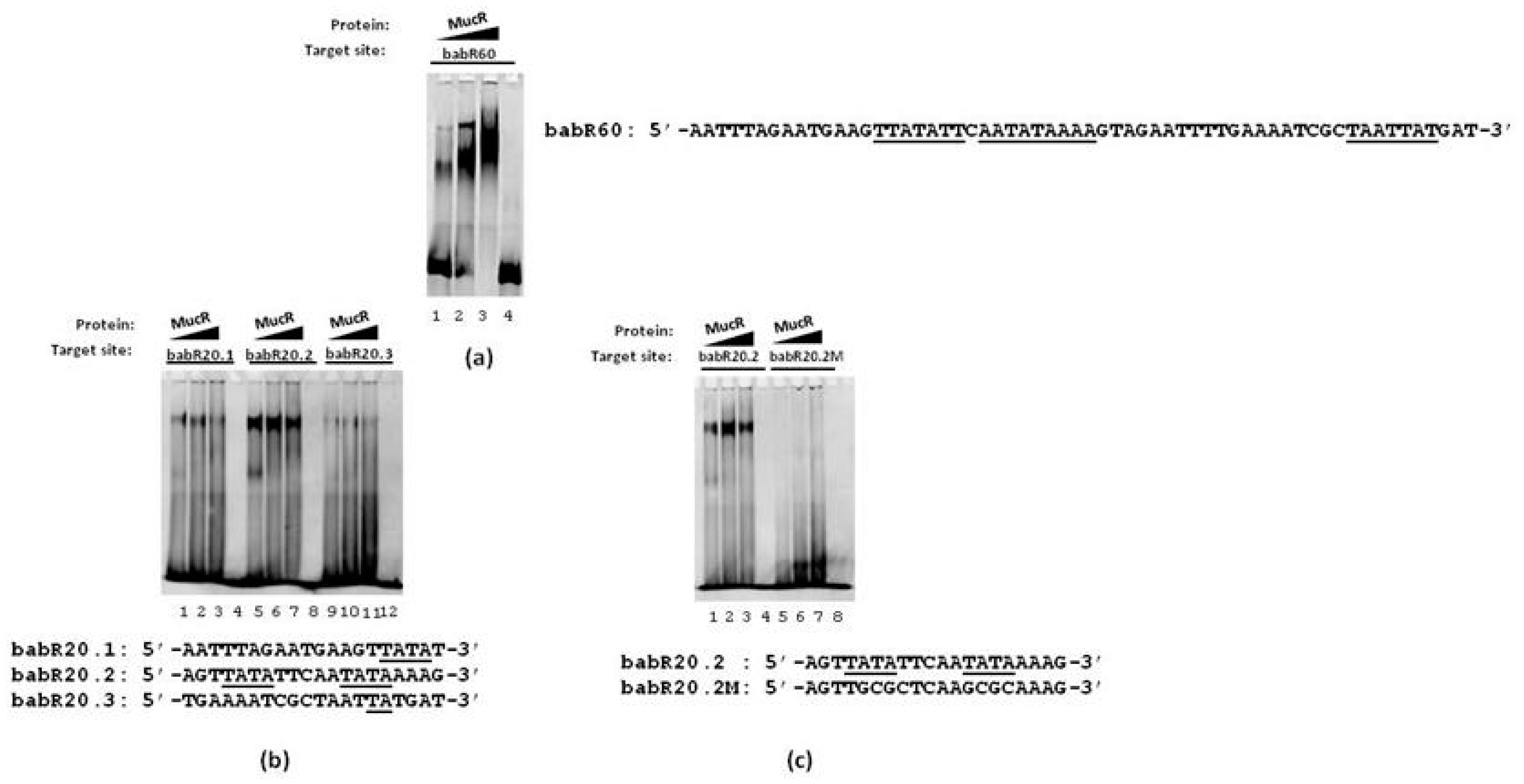
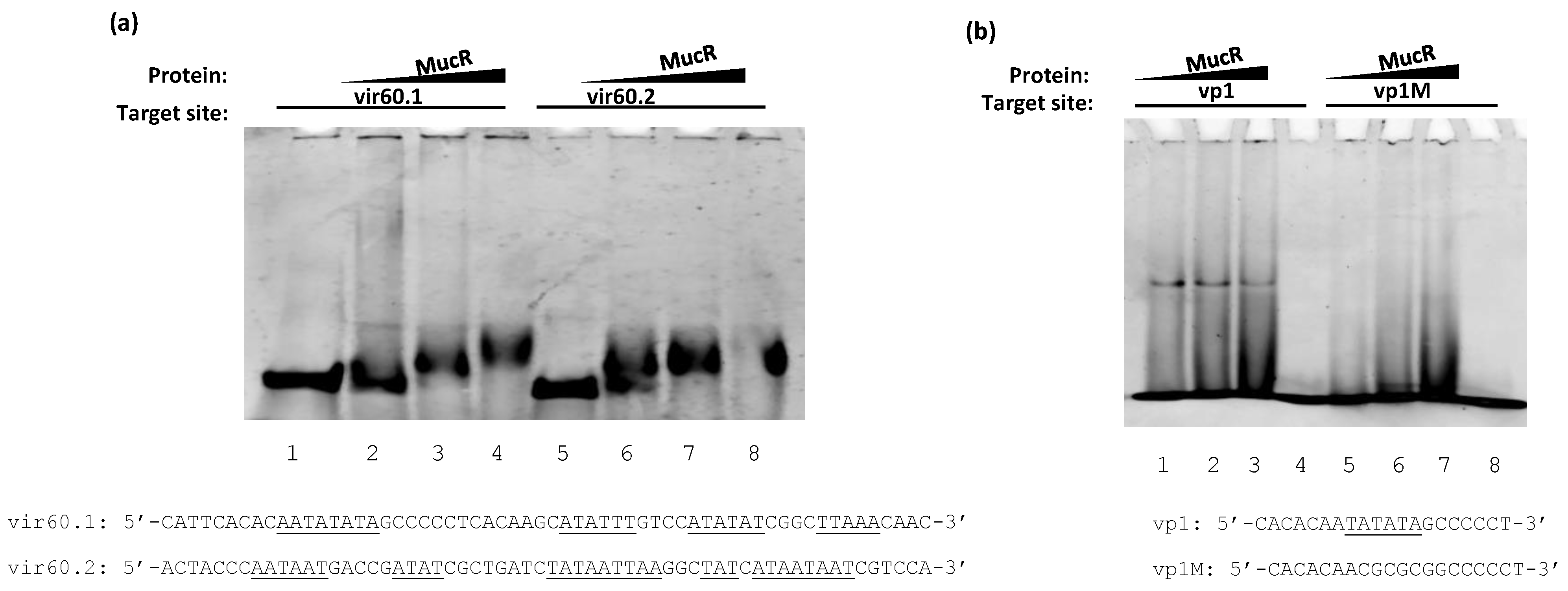
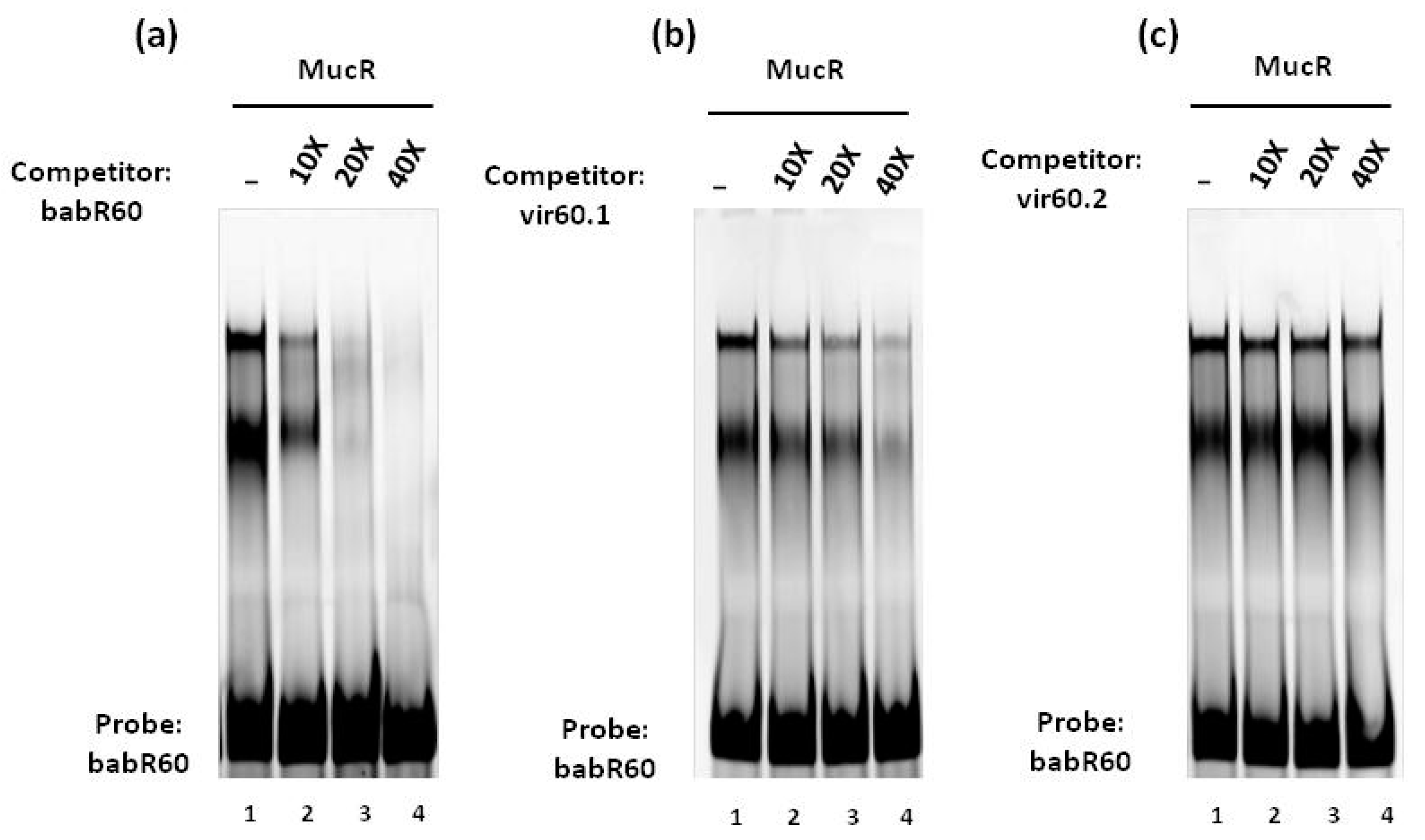
3.1. MucR Binds the Promoters of babR and virB Genes
3.2. MucR Has Minimal Impact on the Expression of the virB Genes in B. abortus 2308
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pappas, G.; Akritidis, N.; Bosilkovski, M.; Tsianos, E. Brucellosis. N. Engl. J. Med. 2005, 352, 2325–2336. [Google Scholar] [CrossRef] [PubMed]
- Cossaboom, C.M.; Kharod, G.A.; Salzer, J.S.; Tiller, R.V.; Campbell, L.P.; Wu, K.; Negrón, M.E.; Ayala, N.; Evert, N.; Radowicz, J.; et al. Notes from the Field: Brucella abortus Vaccine Strain RB51 Infection and Exposures Associated with Raw Milk Consumption—Wise County, Texas, 2017. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longo, M.; Mallardo, K.; Montagnaro, S.; De Martino, L.; Gallo, S.; Fusco, G.; Galiero, G.; Guarino, A.; Pagnini, U.; Iovane, G. Shedding of Brucella abortus rough mutant strain RB51 in milk of water buffalo (Bubalus bubalis). Prev. Vet. Med. 2009, 90, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Caswell, C.C.; Elhassanny, A.E.; Planchin, E.E.; Roux, C.M.; Weeks-Gorospe, J.N.; Ficht, T.A.; Dunman, P.M.; Roop, R.M., 2nd. Diverse genetic regulon of the virulence-associated transcriptional regulator MucR in Brucella abortus. Infect. Immun. 2013, 81, 1040–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arenas-Gamboa, A.M.; Rice-Ficht, A.C.; Kahl-McDonagh, M.M.; Ficht, T.A. Protective efficacy and safety of Brucella melitensis 16MΔmucR against intraperitoneal and aerosol challenge in BALB/c mice. Infect. Immun. 2011, 79, 3653–3658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baglivo, I.; Russo, L.; Esposito, S.; Malgieri, G.; Renda, M.; Salluzzo, A.; Di Blasio, B.; Isernia, C.; Fattorusso, R.; Pedone, P.V. The structural role of the zinc ion can be dispensable in prokaryotic zinc-finger domains. Proc. Natl. Acad. Sci. USA 2009, 106, 6933–6938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malgieri, G.; Russo, L.; Esposito, S.; Baglivo, I.; Zaccaro, L.; Pedone, E.M.; Di Blasio, B.; Isernia, C.; Pedone, P.V.; Fattorusso, R. The prokaryotic Cys2His2 zinc-finger adopts a novel fold as revealed by the NMR structure of Agrobacterium tumefaciens Ros DNA-binding domain. Proc. Natl. Acad. Sci. USA 2007, 104, 17341–17346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Abrosca, G.; Russo, L.; Palmieri, M.; Baglivo, I.; Netti, F.; de Paola, I.; Zaccaro, L.; Farina, B.; Iacovino, R.; Pedone, P.V.; et al. The (unusual) aspartic acid in the metal coordination sphere of the prokaryotic zinc finger domain. J. Inorg. Biochem. 2016, 161, 91–98. [Google Scholar] [CrossRef]
- Baglivo, I.; Palmieri, M.; Rivellino, A.; Netti, F.; Russo, L.; Esposito, S.; Iacovino, R.; Farina, B.; Isernia, C.; Fattorusso, R.; et al. Molecular strategies to replace the structural metal site in the prokaryotic zinc finger domain. Biochim. Biophys. Acta 2014, 1844, 497–504. [Google Scholar] [CrossRef]
- Cooley, M.B.; D’Souza, M.R.; Kado, C.I. The virC and virD operons of the Agrobacterium Ti plasmid are regulated by the ros chromosomal gene: Analysis of the cloned ros gene. J. Bacteriol. 1991, 173, 1608–2616. [Google Scholar] [CrossRef] [Green Version]
- Russo, L.; Palmieri, M.; Caso, J.V.; D’Abrosca, G.; Diana, D.; Malgieri, G.; Baglivo, I.; Isernia, C.; Pedone, P.V.; Fattorusso, R. Towards understanding the molecular recognition process in prokaryotic zinc-finger domain. Eur. J. Med. Chem. 2015, 91, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Baglivo, I.; Pirone, L.; Pedone, E.M.; Pitzer, J.E.; Muscariello, L.; Marino, M.M.; Malgieri, G.; Freschi, A.; Chambery, A.; Roop, R.M., II; et al. Ml proteins from Mesorhizobium loti and MucR from Brucella abortus: An AT-rich core DNA-target site and oligomerization ability. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baglivo, I.; Pirone, L.; Malgieri, G.; Fattorusso, R.; Roop, R.M., II; Pedone, E.M.; Pedone, P.V. MucR binds multiple target sites in the promoter of its own gene and is a heat-stable protein: Is MucR a H-NS-like protein? FEBS Open Bio 2018, 8, 711–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirone, L.; Pitzer, J.E.; D’Abrosca, G.; Fattorusso, R.; Malgieri, G.; Pedone, E.M.; Pedone, P.V.; Roop, R.M., 2nd; Baglivo, I. Identifying the region responsible for Brucella abortus MucR higher-order oligomer formation and examining its role in gene regulation. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, B.R.; Li, Y.; Cote, A.; Weirauch, M.T.; Ding, P.; Hughes, T.R.; Navarre, W.W.; Xia, B.; Liu, J. Structural basis for recognition of AT-rich DNA by unrelated xeogeneic silencing proteins. Proc. Natl. Acad. Sci. USA 2011, 108, 10690–10695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winardhi, R.S.; Yan, J.; Kenney, L.J. H-NS Regulates Gene Expression and Compacts the Nucleoid: Insights from Single-Molecule Experiments. Biophys. J. 2015, 109, 1321–1329. [Google Scholar] [CrossRef] [Green Version]
- Uzureau, S.; Godefroid, M.; Deschamps, C.; Lemaire, J.; De Bolle, X.; Letesson, J.J. Mutations of the quorum sensing-dependent regulator VjbR lead to drastic surface modifications in Brucella melitensis. J. Bacteriol. 2007, 186, 6035–6047. [Google Scholar] [CrossRef] [Green Version]
- Rambow-Larsen, A.A.; Rajashekara, G.; Petersen, E.; Splitter, G. Putative quorum-sensing regulator BlxR of Brucella melitensis regulates virulence factors including the type IV secretion system and flagella. J. Bacterial. 2008, 190, 3274–3282. [Google Scholar] [CrossRef] [Green Version]
- Mueller, K.; González, J.E. Complex regulation of symbiotic functions is coordinated by MucR and quorum sensing in Sinorhizobium meliloti. J. Bacterial. 2011, 193, 485–495. [Google Scholar] [CrossRef] [Green Version]
- Uzureau, S.; Lemaire, J.; Delaive, E.; Dieu, M.; Gaigneaux, A.; Raes, M.; De Bolle, X.; Letesson, J.J. Global analysis of quorum sensing targets in the intracellular pathogen Brucella melitensis 16 M. J. Proteome Res. 2010, 9, 3200–3217. [Google Scholar] [CrossRef]
- Weeks, J.N.; Galindo, C.L.; Drake, K.L.; Garner, H.R.; Fitch, T.A. Brucella melitensis VjbR and C12-HSL regulons: Contributions of N-dodecanoyl homoserine lactone signaling molecule and LuxR homologu VjbR to gene expression. BMC Microbiol. 2010, 10, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caswell, C.C.; Galnes, J.M.; Roop, R.M., II. The RNA chaperone Hfq independently coordinates expression of the VirB type IV secretion system and LuxR-type regulator BabR in Brucella abortus 2308. J. Bacteriol. 2011, 194, 3–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Jong, M.F.; Sun, Y.H.; den Hartigh, A.B.; van Dijl, J.M.; Tsolis, R.M. Identification of VceA and VceC, two members of the VjbR regulon that are translocated into macrophage by the Brucella type IV secretion system. Mol. Micrbiol. 2008, 70, 1378–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sieira, R.; Comerci, D.J.; Sánchez, D.O.; Ugalde, R.A. A homologue of an operon required for DNA transfer in Agrobacterium is required in Brucella abortus for virulence and intracellular multiplication. J. Bacteriol. 2000, 182, 4849–4855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Jong, M.F.; Tsolis, R.M. Brucellosis and type IV secretion. Future Microbiol. 2012, 7, 47–58. [Google Scholar] [CrossRef]
- Celli, J. The changing nature of the Brucella-containing vacuole. Cell. Microbiol. 2015, 17, 951–958. [Google Scholar] [CrossRef] [Green Version]
- Tendeng, C.; Bertin, P.N. H-NS in Gram-negative bacteria: A family of multifaceted proteins. Trends Microbiol. 2003, 11, 511–518. [Google Scholar] [CrossRef]
- Anvar, Z.; Cammisa, M.; Riso, V.; Baglivo, I.; Kukreja, H.; Sparago, A.; Girardot, M.; Lad, S.; De Feis, I.; Cerrato, F.; et al. ZFP57 recognizes multiple and closely spaced sequence motif variants to maintain repressive epigenetic marks in mouse embryonic stem cells. Nucleic Acids Res. 2016, 44, 1118–1132. [Google Scholar] [CrossRef] [Green Version]
- Budnick, J.A.; Sheehan, L.M.; Kang, L.; Michalak, P.; Caswell, C.C. Characterization of Three Small Proteins in Brucella abortus Linked to Fucose Utilization. J. Bacteriol. 2018, 200. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Scmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and 2(-Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- OIE World Organization for Animal Health. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; Chapter 3.1.4: Brucellosis (Brucella abortus, B. melitensis and B. suis) (infection with B. abortus, B. melitensis and B. suis); OIE World Organization for Animal Health: Paris, France, 2016. [Google Scholar]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, H.; Liu, W.; Peng, X.; Jing, Z.; Wu, Q. The effects of MucR on expression of type IV secretion system, quorum sensing system and stress responses in Brucella melitensis. Vet. Microbiol. 2013, 166, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Rihtar, E.; Žgur Bertok, D.; Podlesek, Z. The Uropathogenic Specific Protein Gene usp from Escherichia coli and Salmonella bongori is a Novel Member of the TyrR and H-NS Regulons. Microorganisms 2020, 8, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sieira, R. Regulation f virulence in Brucella: An ecletic repertoire of transcription factors defines the complex architecture of the virB promoter. Future Microbiol. 2013, 8, 1193–1208. [Google Scholar] [CrossRef]
- Sieira, R.; Commerci, D.J.; Pietrasanta, L.I.; Ugalde, R.A. Integration host factor is involved in trascriptional regulation of Brucella abortus virB operon. Mol. Microbiol. 2004, 54, 808–822. [Google Scholar] [CrossRef]
- Malgieri, G.; D’Abrosca, G.; Pirone, L.; Toto, A.; Palmieri, M.; Russo, L.; Sciacca, M.F.M.; Tatè, R.; Sivo, V.; Baglivo, I.; et al. Folding mechanisms steer the amyloid fibril formation propensity of highly homologous proteins. Chem. Sci. 2018, 9, 3290–3298. [Google Scholar] [CrossRef] [Green Version]
- Fang, F.C.; Rimsky, S. New insights into transcriptional regulation by H-NS. Curr. Opin. Microbiol. 2008, 11, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Paradiso, R.; Orsini, M.; Riccardi, M.G.; Cecere, B.; Cerrone, A.; Cammà, C.; Chiusano, M.L.; Galiero, G.; Borriello, G. Complete Genome Sequencing of eight Brucella abortus biovar 1 strains isolated from water buffalo. Genome Announc. 2018, 29. [Google Scholar] [CrossRef] [Green Version]
- Paradiso, R.; Orsini, M.; Riccardi, M.G.; Criscuolo, D.; Borrelli, R.; Valvini, O.; Cammà, C.; Chiusano, M.L.; Galiero, G.; Borriello, G. Complete Genome Sequencing of ten Brucella abortus biovar 3 strains isolated from water buffalo. Genome Announc. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Hui, Y.; Zai, X.; Xu, J.; Liang, L.; Wang, B.; Yue, J.; Li, S. Comparative genomic analysis of Brucella abortus vaccine strain 104M reveals a set of candidate genes associated with its virulence attenuation. Virulence 2015, 6, 745–754. [Google Scholar] [CrossRef] [Green Version]
- Fumeaux, C.; Radhakrishnan, S.K.; Ardissone, S.; Théraulaz, L.; Frandi, A.; Martins, D.; Nesper, J.; Abel, S.; Jenal, U.; Viollier, P.H. Cell cycle transition from S-phase to G1 in Caulobacter is mediated by ancestral virulence regulators. Nat. Commun. 2014, 5, 1–14. [Google Scholar] [CrossRef] [PubMed]
- De Bolle, X.; Crosson, S.; Matroule, J.Y.; Leteson, J.J. Brucella abortus cell cycle and infection are coordinated. Trends Microbiol. 2015, 23, 812–821. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.S.; Soo, J.; Rao, C.; Leung, A.S.; Ngai, D.H.M.; Ensminger, A.W.; Navarre, W.W. Silencing by H-NS Potentiated the Evolution. PLoS Pathog. 2014, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucchini, S.; Rowley, G.; Goldberg, M.D.; Hurd, D.; Harrison, M.; Hinton, J.C. H-NS mediates the silencing of laterally acquired genes in bacteria. PLoS Pathog. 2006, 2. [Google Scholar] [CrossRef] [PubMed]
- Navarre, W.W.; Porwollik, S.; Wang, Y.; McClelland, M.; Rosen, H.; Libby, S.J.; Fang, F.C. Selective Silencing of Foreign DNA with Low GC Content by the H-NS Protein in Salmonella. Science 2006, 313, 236–238. [Google Scholar] [CrossRef] [PubMed]
- Dorman, C.J. H-NS, the genome sentinel. Nat. Rev. Microbiol. 2007, 5, 157–161. [Google Scholar] [CrossRef]
- Sun, Y.H.; Rolan, G.; den Hartigh, A.B.; Sondervan, D.; Tsolis, R.M. Brucella abortus VirB12 is expressed during infection but is not an essential component of the type IV secretion system. Infect. Immun. 2005, 73, 6048–6054. [Google Scholar] [CrossRef] [Green Version]
- Borriello, G.; Peletto, S.; Lucibelli, M.G.; Acutis, P.L.; Ercolini, D.; Galiero, G. Link between geographical origin and occurrence of Brucella abortus biovars in cow and water buffalo herds. Appl. Environ. Microbiol. 2013, 79, 1039–1043. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borriello, G.; Russo, V.; Paradiso, R.; Riccardi, M.G.; Criscuolo, D.; Verde, G.; Marasco, R.; Pedone, P.V.; Galiero, G.; Baglivo, I. Different Impacts of MucR Binding to the babR and virB Promoters on Gene Expression in Brucella abortus 2308. Biomolecules 2020, 10, 788. https://doi.org/10.3390/biom10050788
Borriello G, Russo V, Paradiso R, Riccardi MG, Criscuolo D, Verde G, Marasco R, Pedone PV, Galiero G, Baglivo I. Different Impacts of MucR Binding to the babR and virB Promoters on Gene Expression in Brucella abortus 2308. Biomolecules. 2020; 10(5):788. https://doi.org/10.3390/biom10050788
Chicago/Turabian StyleBorriello, Giorgia, Veronica Russo, Rubina Paradiso, Marita Georgia Riccardi, Daniela Criscuolo, Gaetano Verde, Rosangela Marasco, Paolo Vincenzo Pedone, Giorgio Galiero, and Ilaria Baglivo. 2020. "Different Impacts of MucR Binding to the babR and virB Promoters on Gene Expression in Brucella abortus 2308" Biomolecules 10, no. 5: 788. https://doi.org/10.3390/biom10050788