Emerging Allergens in Goji Berry Superfruit: The Identification of New IgE Binding Proteins towards Allergic Patients’ Sera

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Clinical Cases Description
Skin Prick Tests
2.3. Sample Preparation
2.4. Electrophoretic Separation and Protein Assay
2.5. Proteomic Investigation
2.5.1. Protein Profiling of Goji Berries
2.5.2. In-Gel Protein Digestion
2.5.3. LC-HR-MS/MS Analysis and Bioinformatics
2.6. Immunoblotting Experiment
3. Results
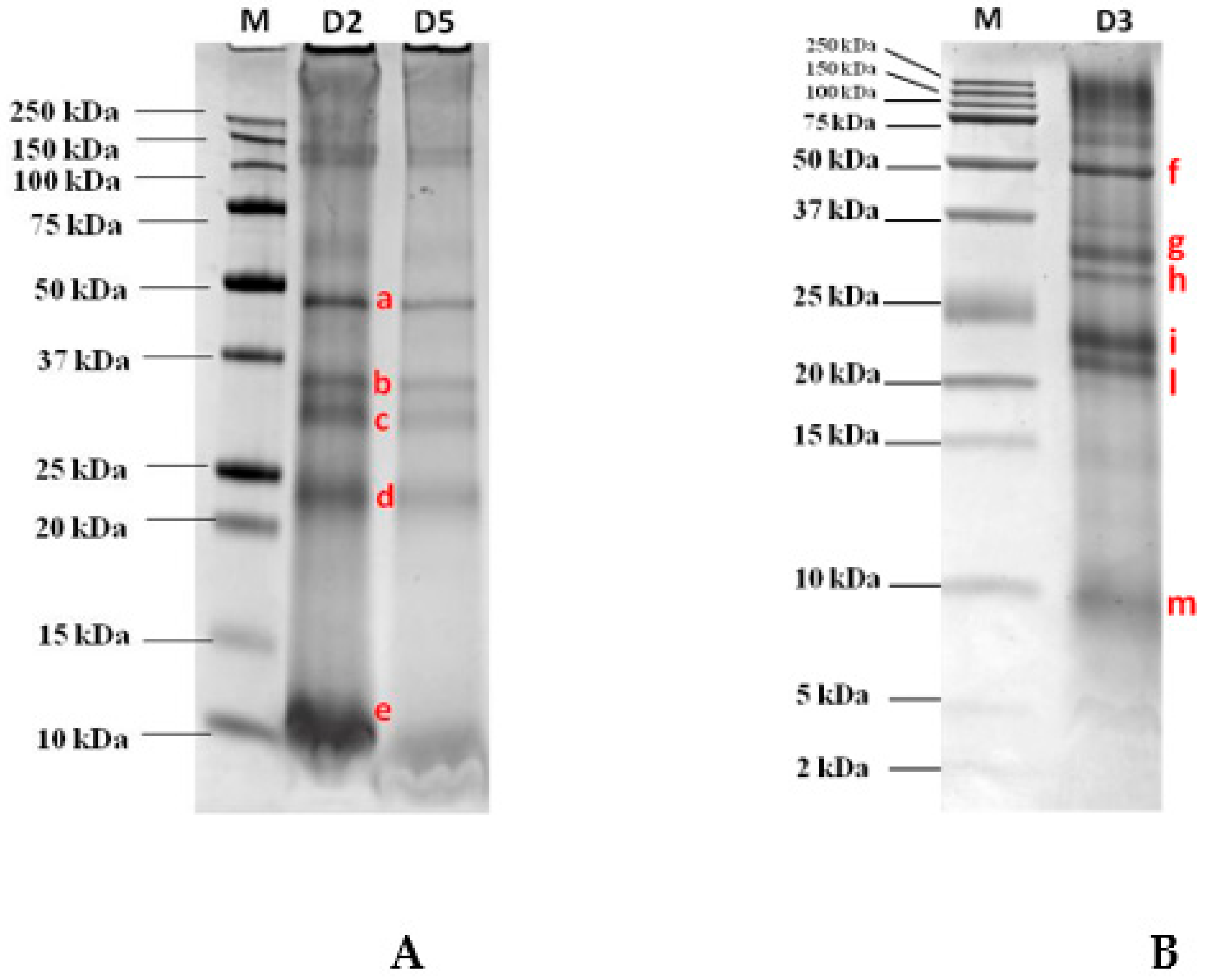
3.1. Gel Electrophoresis Separation and Immunoblot Analysis
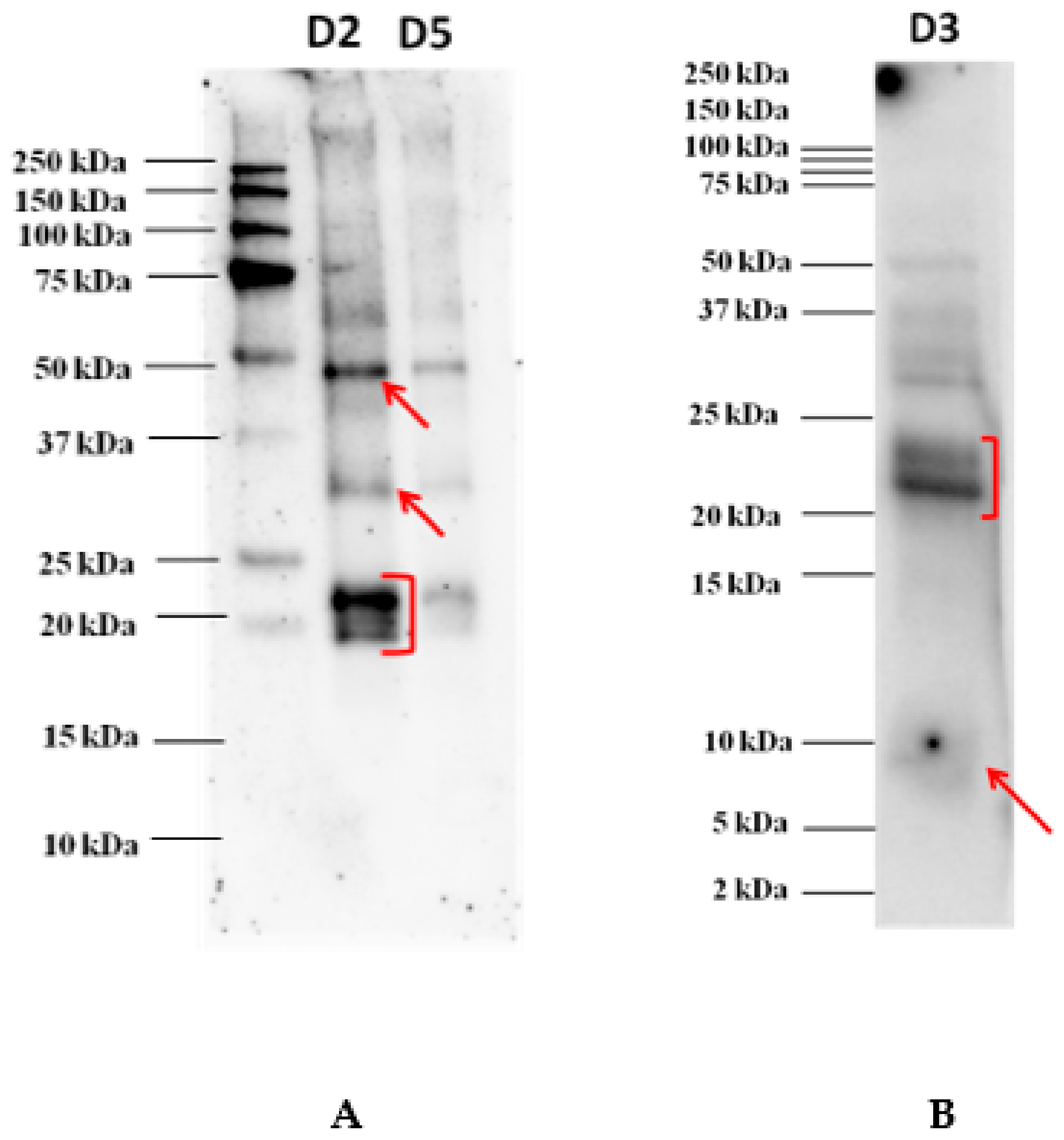
3.2. Immunoblot Analysis After Electrophoretic Separation
3.3. LC-HR-MS/MS Identification of the Excised Protein Bands
3.4. Bottom-Up Proteomics of Goji Berry Extract
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Amagase, H.; Farnsworth, N.R. A review of botanical characteristics, phytochemistry, clinical relevance in efficacy and safety of Lycium barbarum fruit (Goji). Food Res. Int. 2011, 44, 1702–1717. [Google Scholar] [CrossRef]
- Blaschek, W.; Hänsel, R.; Keller, K.; Reichling, J.; Rimpler, H.; Schneider, G. Hagers Handbuch der Pharmazeutischen Praxis: Folgeband 2: Drogen AK; Blaschek, W., Hänsel, R., Keller, K., Reichling, J., Rimpler, H., Schneider, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Chang, S.K.; Alasalvar, C.; Shahidi, F. Superfruits: Phytochemicals, antioxidant efficacies, and health effects–A comprehensive review. Crit. Rev. Food Sci. 2019, 59, 1580–1604. [Google Scholar] [CrossRef]
- Niro, S.; Fratianni, A.; Panfili, G.; Falasca, L.; Cinquanta, L.; Alam, M.R. Nutritional evaluation of fresh and dried goji berries cultivated in Italy. Ital. J. Food Sci. 2017, 29, 398–408. [Google Scholar] [CrossRef]
- Kulczyński, B.; Gramza-Michałowska, A. Goji berry (Lycium barbarum): Composition and health effects—A review. Pol. J. Food Nutr. Sci. 2016, 66, 67–76. [Google Scholar] [CrossRef]
- Larramendi, C.H.; García-Abujeta, J.L.; Vicario, S.; García-Endrino, A.; López-Matas, M.A.; García-Sedeño, M.D.; Carnés, J. Goji berries (Lycium barbarum): Risk of allergic reactions in individuals with food allergy. J. Investig. Allergol. Clin. Immunol. 2012, 22, 345–350. [Google Scholar] [PubMed]
- Monzón Ballarín, S.; López-Matas, M.A.; Sáenz Abad, D.; Pérez-Cinto, N.; Carnés, J. Anaphylaxis associated with the ingestion of Goji berries (Lycium barbarum). J. Investig. Allergol. Clin. Immunol. 2011, 21, 567–570. [Google Scholar] [PubMed]
- Carnés, J.; De Larramendi, C.H.; Ferrer, A.; Huertas, A.J.; López-Matas, M.A.; Pagán, J.A.; Navarro, L.A.; García-Abujeta, J.L.; Vicario, S.; Peña, M. Recently introduced foods as new allergenic sources: Sensitisation to Goji berries (Lycium barbarum). Food Chem. 2013, 137, 130–135. [Google Scholar] [CrossRef]
- Carnés, J.; de Larramendi, C.H.; López-Matas, M.A.; Ferrer, A.; Huertas, J. Allergenic sensitisation mediated by Wolfberry. In Lycium barbarum and Human Health, 1st ed.; Chang, R.C.-C., So, K.-F., Eds.; Springer: Dordrecht, The Netherlands; Berlin/Heidelberg, Germany; New York, NY, USA, 2015; pp. 179–198. [Google Scholar]
- Teixeira, S.; Luísb, I.M.; Oliveira, M.M.; Abreu, I.A.; Batista, R. Goji berries superfood—Contributions for the characterisation of proteome and IgE-binding proteins. Food Agric. Immunol. 2019, 30, 262–280. [Google Scholar] [CrossRef] [Green Version]
- Brusca, I.; Barrale, M.; Onida, R.; La Chiusa, S.M.; Gjomarkaj, M.; Uasuf, C.G. The extract, the molecular allergen or both for the in vitro diagnosis of peach and peanut sensitization? Clin. Chim. Acta 2019, 493, 25–30. [Google Scholar] [CrossRef]
- De Angelis, E.; Pilolli, R.; Bavaro, S.L.; Monaci, L. Insight into the gastro-duodenal digestion resistance of soybean proteins and potential implications for residual immunogenicity. Food Funct. 2017, 8, 1599–1610. [Google Scholar] [CrossRef]
- De Angelis, E.; Bavaro, S.L.; Forte, G.; Pilolli, G.; Monaci, L. Heat and pressure treatments on almond protein stability and change in immunoreactivity after simulated human digestion. Nutrients 2018, 10, 1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bavaro, S.L.; De Angelis, E.; Barni, S.; Pilolli, R.; Mori, F.; Novembre, E.M.; Monaci, L. Modulation of milk allergenicity by baking milk in foods: A proteomic investigation. Nutrients 2019, 11, 1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheurer, S.; Pastorello, E.A.; Wangorsch, A.; Kästner, M.; Haustein, D.; Vieths, S. Recombinant allergens Pru av 1 and Pru av 4 and a newly identified lipid transfer protein in the in vitro diagnosis of cherry allergy. J. Allergy Clin. Immunol. 2001, 107, 724–731. [Google Scholar] [CrossRef]
- Pastorello, E.A.; D’Ambrosio, F.P.; Pravettoni, V.; Farioli, L.; Giuffrida, G.; Monza, M.; Ansaloni, R.; Fortunate, D.; Scibola, E.; Rivolta, F.; et al. Evidence for a lipid transfer protein as the major allergen of apricot. J. Allergy Clin. Immunol. 2000, 105, 371–377. [Google Scholar] [CrossRef]
- Pastorello, E.A.; Farioli, L.; Pravettoni, V.; Ortolani, C.; Ispano, M.; Monza, M.; Baroglio, C.; Scibola, E.; Ansaloni, R.; Incorvaia, C.; et al. The major allergen of peach (Prunus persica) is a lipid transfer protein. J. Allergy Clin. Immunol. 1999, 103, 520–526. [Google Scholar] [CrossRef]
- Pastorello, E.A.; Pravettoni, V.; Farioli, L.; Ispano, M.; Fortunato, D.; Monza, M.; Giuffrida, M.G.; Rivolta, F.; Scibola, E.; Ansaloni, R.; et al. Clinical role of a lipid transfer protein that acts as a new apple- specific allergen. J. Allergy Clin. Immunol. 1999, 104, 1099–1106. [Google Scholar] [CrossRef]
- Geiselhart, S.; Hoffmann-Sommergruber, K.; Bublin, M. Tree nut allergens. Mol. Immunol. 2018, 100, 71–81. [Google Scholar] [CrossRef]
- Monaci, L.; Pilolli, R.; De Angelis, E.; Crespo, J.; Novak, N.; Cabanillas, B. Food allergens: Classification, molecular properties, characterization, and detection in food sources. Adv. Food Nutr. Res. 2020, in press. [Google Scholar] [CrossRef]
- Kesari, P.; Sharma, A.; Katiki, M.; Kumar, P.; R Gurjar, B.; Tomar, S.; K Sharma, A.; Kumar, P. Structural, functional and evolutionary aspects of seed globulins. Protein Peptide Lett. 2017, 24, 267–277. [Google Scholar] [CrossRef]
- Asero, R.; Piantanida, M.; Pinter, E.; Pravettoni, V. The clinical relevance of lipid transfer protein. Clin. Experim. Allergy 2017, 48, 6–12. [Google Scholar] [CrossRef] [Green Version]
- Matricardi, P.M.; Kleine-Tebbe, J.; Hoffmann, H.J.; Valenta, R.; Hilger, C.; Hofmaier, S.; Barber, D. EAACI molecular allergology user’s guide. Pediatr. Allergy Immunol. 2016, 27 (Suppl. 23), 1–250. [Google Scholar] [CrossRef] [PubMed]
- Uasuf, C.G.; Villalta, D.; Conte, M.E.; Di Sano, C.; Barrale, M.; Cantisano, V.; Pace, E.; Gjomarkaj, M.; Gangemi, S.; Brusca, I. Different co-sensitizations could determine different risk assessment in peach allergy? Evaluation of an anaphylactic biomarker in Pru p 3 positive patients. Clin. Mol. Allergy. 2015, 13, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scala, E.; Till, S.J.; Asero, R.; Abeni, D.; Guerra, E.C.; Pirrotta, L.; Paganelli, R.; Pomponi, D.; Giani, M.; De Pita, O.; et al. Lipid transfer protein sensitization: Reactivity profiles and clinical risk assessment in an Italian cohort. Allergy 2015, 70, 933–943. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| AnyKD Gel | Tris-Tricine Gel | Putative Identification (Accession) | Type of Protein | Coverage (%) | Peptides (Unique) | MW | Protein Score |
|---|---|---|---|---|---|---|---|
| a | F | A0A2R2JFS1 | Vicilin, OS = Solanum melongena | 14.08 | 6 (4) | 48.6 | 3.17 |
| A0A1S4CJA8 | Vicilin-like, OS = Nicotiana tabacum | 6.53 | 7 (4) | 94.6 | 2.7 | ||
| b | G | A0A0A7DVY6 | Fibrillin, OS = Lycium barbarum | 51.09 | 13 (13) | 35.3 | 16.74 |
| c | H | A0A3Q7IWI5 | Uncharacterized protein, OS = Solanum lycopersicum | 12.87 | 6 (6) | 53.3 | 3.75 |
| A0A0A7DVY6 (Fibrillin) | Fibrillin, OS = Lycium barbarum | 29.28 | 8 (8) | 35.3 | 1.8 | ||
| d | I | A0A1J6JXR5 (glutelin) | Glutelin-like, OS = Nicotiana attenuata | 14.73 | 9 (4) | 53.6 | 10.93 |
| L | A0A2G2ZXN5 (legumin) | Legumin-like, OS = Capsicum annuum | 9.72 | 6 (1) | 52.9 | 3.82 | |
| e | M | B3A0N2 | LTP, OS = Lycium barbarum (fragment) | 39.21 | 1 (1) | 4.9 | 6.52 |
| N. | Accession | Description | Coverage (%) | Peptides (Unique) | PSMs | Score Sequest HT |
|---|---|---|---|---|---|---|
| 1 | A0A0A7DVY6 | Fibrillin OS = Lycium barbarum | 69.16 | 18 (13) | 302 | 524.75 |
| 2 | A0A3Q7IWI5 | Uncharacterized protein OS = Solanum lycopersicum | 23.21 | 11 (4) | 187 | 271.84 |
| 3 | A0A1J6JXR5 | Glutelin type-b 5 OS = Nicotiana attenuata | 22.53 | 12 (4) | 196 | 247.78 |
| 4 | A0A1U7X9B2 | 11S globulin seed storage protein 2-like OS = Nicotiana sylvestris | 19.02 | 10 (2) | 116 | 87.44 |
| 5 | M1B1M5 | Uncharacterized protein OS = Solanum tuberosum | 15.81 | 9 (1) | 115 | 81.78 |
| 6 | A0A3Q7FEY7 | Uncharacterized protein OS = Solanum lycopersicum | 4.52 | 8 (1) | 81 | 67.30 |
| 7 | A0A1J6L9S7 | 11s globulin subunit beta OS = Nicotiana attenuata | 17.10 | 13 (1) | 124 | 66.05 |
| 8 | A0A1U7VG63 | Legumin B-like OS = Nicotiana sylvestris | 21.17 | 12 (2) | 119 | 55.54 |
| 9 | A0A2G9GMS2 | Uncharacterized protein OS = Handroanthus impetiginosus | 12.66 | 4 (1) | 41 | 53.98 |
| 10 | A0A144YUS5 | Ribulose bisphosphate carboxylase large chain OS = Iochroma lehmannii | 23.69 | 10 (2) | 75 | 53.58 |
| 11 | A0A022S2J1 | Uncharacterized protein OS = Erythranthe guttata | 5.18 | 3 (2) | 48 | 53.58 |
| 12 | G0WZI6 | Ribulose bisphosphate carboxylase large chain (Fragment) OS = Solanum chenopodioides | 16.56 | 7 (1) | 42 | 51.71 |
| 13 | A0A2G2X5V4 | 11S globulin seed storage protein 2 OS = Capsicum baccatum | 14.95 | 9 (2) | 89 | 50.63 |
| 14 | A0A1U7W1Q9 | Vicilin-like antimicrobial peptides 2-3 OS = Nicotiana sylvestris | 10.80 | 9 (6) | 92 | 49.82 |
| 15 | M1A8H0 | Uncharacterized protein OS = Solanum tuberosum | 66.67 | 5 (4) | 56 | 46.28 |
| 16 | A0A2R2JFS1 | SM80.1 Vicilin OS = Solanum melongena | 14.79 | 7 (3) | 77 | 44.34 |
| 17 | Q948T8 | Histone H4 (Fragment) OS=Citrus jambhiri | 45.10 | 5 (2) | 40 | 43.92 |
| 18 | A0A1U8GRB4 | catechol oxidase B, chloroplastic OS = Capsicum annuum | 19.35 | 10 (2) | 74 | 41.66 |
| 19 | A0A2C9V2W2 | Uncharacterized protein OS = Manihot esculenta | 22.99 | 10 (0) | 91 | 38.72 |
| 20 | A0A103XB50 | Histone H4 OS = Cynara cardunculus var. scolymus | 45.63 | 5 (1) | 46 | 36.74 |
| 21 | M1ANI2 | Uncharacterized protein OS = Solanum tuberosum | 49.59 | 6 (3) | 37 | 35.68 |
| 22 | A0A1U8F773 | Uncharacterized protein OS = Capsicum annuum | 8.89 | 4 (1) | 47 | 34.82 |
| 23 | A0A059AJT0 | Uncharacterized protein OS = Eucalyptus grandis | 18.40 | 7 (1) | 75 | 34.60 |
| 24 | A0A1S4BK33 | 11S globulin subunit beta-like OS = Nicotiana tabacum | 11.58 | 7 (2) | 70 | 33.12 |
| 25 | A7UGG9 | Non-specific lipid-transfer protein OS = Solanum tuberosum | 17.54 | 2 (1) | 21 | 30.61 |
| 26 | Q40151 | Hsc70 OS = Solanum lycopersicum | 18.28 | 6 (1) | 63 | 27.21 |
| 27 | B3A0N2 | Non-specific lipid-transfer protein (Fragments) OS = Lycium barbarum | 39.22 | 1 (1) | 21 | 26.93 |
| 28 | F1DBB8 | Chloroplast polyphenol oxidase OS = Solanum melongena | 20.31 | 7 (1) | 68 | 26.24 |
| 29 | C0SQK3 | Elongation factor1-alpha (Fragment) OS = Rosa hybrid cultivar | 14.63 | 3 (0) | 35 | 26.01 |
| 30 | K4D1U9 | Non-specific lipid-transfer protein OS= Solanum lycopersicum | 25.44 | 2 (1) | 27 | 25.73 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uasuf, C.G.; De Angelis, E.; Guagnano, R.; Pilolli, R.; D’Anna, C.; Villalta, D.; Brusca, I.; Monaci, L. Emerging Allergens in Goji Berry Superfruit: The Identification of New IgE Binding Proteins towards Allergic Patients’ Sera. Biomolecules 2020, 10, 689. https://doi.org/10.3390/biom10050689
Uasuf CG, De Angelis E, Guagnano R, Pilolli R, D’Anna C, Villalta D, Brusca I, Monaci L. Emerging Allergens in Goji Berry Superfruit: The Identification of New IgE Binding Proteins towards Allergic Patients’ Sera. Biomolecules. 2020; 10(5):689. https://doi.org/10.3390/biom10050689
Chicago/Turabian StyleUasuf, Carina Gabriela, Elisabetta De Angelis, Rocco Guagnano, Rosa Pilolli, Claudia D’Anna, Danilo Villalta, Ignazio Brusca, and Linda Monaci. 2020. "Emerging Allergens in Goji Berry Superfruit: The Identification of New IgE Binding Proteins towards Allergic Patients’ Sera" Biomolecules 10, no. 5: 689. https://doi.org/10.3390/biom10050689