Acorus gramineusand and Euodia ruticarpa Steam Distilled Essential Oils Exert Anti-Inflammatory Effects Through Decreasing Th1/Th2 and Pro-/Anti-Inflammatory Cytokine Secretion Ratios In Vitro


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Steam Distillation Essential Oils (SDEO) from Two Selected Herbs
2.2. Potential Phytochemicals Determination including Total Phenolic, Flavonoid and Saponin Contents in AG and ER SDEO
2.2.1. Total Phenolic Content
2.2.2. Total Flavonoid Content
2.2.3. Total Saponin Content
2.3. Determination of Chemical Components of ER SDEO using Gas Chromatography-Mass Spectrometry (GC-MS)
2.4. Experimental Animals
2.5. Isolation of Mouse Primary Peritoneal Macrophages and Splenocytes
2.5.1. Isolation of Primary Peritoneal Macrophages
2.5.2. Primary Splenocytes Isolation
2.6. Determination of Optimal AG and ER SDEO Concentrations
2.7. Mouse Splenocytes Cultures with AR and ER SDEO at Different Optimal Concentrations
2.8. Mouse Peritoneal Macrophages Cultures with AG and ER SDEO at Different Optimal Concentrations in the Absence or Presence of LPS
2.9. Th1/Th2 and pro-/anti-Inflammatory Cytokine Levels Secreted by Immune Cells Measured Using an Enzyme-Linked Immunosorbent Assay (ELISA)
2.10. Statistical Analysis
3. Results and Discussion
3.1. Total Flavonoid, Phenolic and Saponin Contents in AG and ER Selected SDEO
3.2. Optimal AG and ER SDEO Concentrations using Murine Splenocytes
3.3. AG and ER SDEO Effects on Th1/Th2 Cytokine Secretions by Mouse Primary Splenocytes
3.4. AG and ER SDEO Effects on pro-/anti-Inflammatory Cytokine Secretions by Mouse Peritoneal Macrophages in the Absence or Presence of LPS
3.5. The Correlation between Th1/Th2 Cytokine Secretion Levels in Mouse Primary Splenocyte Cultures and Total Flavonoid, Phenol or Saponin Contents in AG and ER SDEO
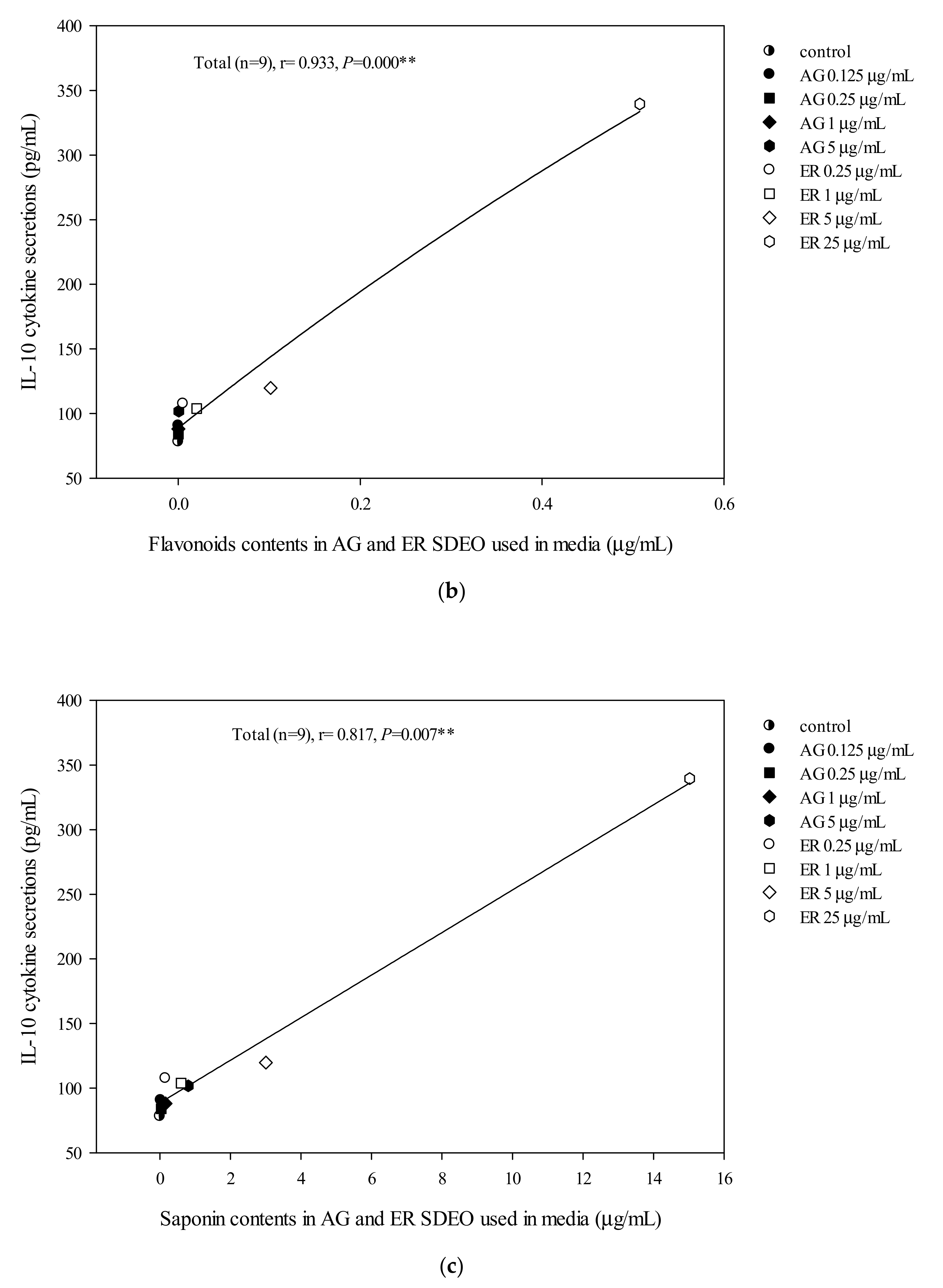
3.6. The Correlation between pro-/anti-Inflammatory Cytokine Secretion Levels in Mouse Primary Peritoneal Macrophage Cultures in the Absence or Presence of LPS and Total Flavonoid, Phenol or Saponin Contents in AG and ER SDEO
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vasto, S.; Candore, G.; Balistreri, C.R.; Caruso, M.; Colonna-Romano, G.; Grimaldi, M.P.; Listi, F.; Nuzzo, D.; Lio, D.; Caruso, C. Inflammatory networks in ageing, age-related diseases and longevity. Mech. Ageing Dev. 2007, 128, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Adeyemi, O.O.; Yemitan, O.K.; Afolabi, L. Inhibition of chemically induced inflammation and pain by orally and topically administered leaf extract of Manihot esculenta Crantz in rodents. J. Ethnopharmacol. 2008, 119, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.R.; Lin, J.Y. Quercetin intraperitoneal administration ameliorates lipopolysaccharide-induced systemic inflammation in mice. Life Sci. 2015, 137, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Senedese, J.M.; Rinaldi-Neto, F.; Furtado, R.A.; Nicollela, H.D.; de Souza, L.D.R.; Ribeiro, A.B.; Ferreira, L.S.; Magalhães, G.M.; Carlos, I.Z.; da Silva, J.J.M.; et al. Chemopreventive role of Copaifera reticulata Ducke oleoresin in colon carcinogenesis. Biomed. Pharmacother. 2019, 111, 331–337. [Google Scholar] [CrossRef]
- Sethi, G.; Shanmugam, M.K.; Ramachandran, L.; Kumar, A.P.; Tergaonkar, V. Multifaceted link between cancer and inflammation. Biosci. Rep. 2012, 32, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Moh, S.H.; Kim, S.B.; Yang, Y.; Kim, E.; Lee, Y.W.; Cho, C.K.; Kim, K.H.; Yoo, B.C.; Cho, J.Y.; et al. HangAmDan-B, an ethnomedicinal herbal mixture, suppresses inflammatory responses by inhibiting Syk/NF-kappaB and JNK/ATF-2 pathways. J. Med. Food 2013, 16, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Steward, W.P.; Brown, K. Cancer chemoprevention: A rapidly evolving field. Br. J. Cancer 2013, 109, 1–7. [Google Scholar] [CrossRef]
- Dickinson, S.E.; Olson, E.R.; Levenson, C.; Janda, J.; Rusche, J.J.; Alberts, D.S.; Bowden, G.T. A novel chemopreventive mechanism for a traditional medicine: East Indian sandalwood oil induces autophagy and cell death in proliferating keratinocytes. Arch. Biochem. Biophys. 2014, 558, 143–152. [Google Scholar] [CrossRef] [Green Version]
- Mokhtar, M.M.; Shaban, H.M.; Hegazy, M.E.A.F.; Ali, S.S. Evaluating the potential cancer chemopreventive efficacy of two different solvent extracts of Seriphidium herba-alba in vitro. Bull. Fac. Pharm. Cairo Univ. 2017, 55, 195–201. [Google Scholar] [CrossRef]
- Kidd, P. Th1/Th2 balance: The hypothesis, its limitations, and implications for health and disease. Altern. Med. Rev. 2003, 8, 223–246. [Google Scholar]
- Gagliani, N.; Huber, S. Basic aspects of T Helper cell differentiation. Methods Mol. Biol. 2017, 1514, 19–30. [Google Scholar] [PubMed]
- Kutukculer, N.; Azarsiz, E.; Aksu, G.; Karaca, N.E. CD4+CD25+Foxp3+ T regulatory cells, Th1 (CCR5, IL-2, IFN-gamma) and Th2 (CCR4, IL-4, IL-13) type chemokine receptors and intracellular cytokines in children with common variable immunodeficiency. Int. J. Immunopathol. Pharmacol. 2016, 29, 241–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuzaki, J.; Tsuji, T.; Imazeki, I.; Ikeda, H.; Nishimura, T. Immunosteroid as a regulator for Th1/Th2 balance: Its possible role in autoimmune diseases. Autoimmunity 2005, 38, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Peng, A.; Feng, Y.; Zhang, X.; Liu, F.; Chen, C.; Ye, X.; Qu, J.; Jin, C.; Wang, M.; et al. Detection of T lymphocyte subsets and related functional molecules in follicular fluid of patients with polycystic ovary syndrome. Sci. Rep. 2019, 9, 6040. [Google Scholar] [CrossRef] [PubMed]
- Raut, J.S.; Karuppayil, S.M. A status review on the medicinal properties of essential oils. Ind. Crop. Prod. 2014, 62, 250–264. [Google Scholar] [CrossRef]
- Edris, A.E. Pharmaceutical and therapeutic potentials of essential oils and their individual volatile constituents: A review. Phytother. Res. 2007, 21, 308–323. [Google Scholar] [CrossRef]
- Slamenova, D.; Horvathova, E. Cytotoxic, anti-carcinogenic and antioxidant properties of the most frequent plant volatiles. Neoplasma 2013, 60, 343–354. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.B.; Niu, W.M.; Yang, X.H.; Wang, Y.; Wang, W.G. Study on perfume stimulating olfaction with volatile oil of Acorus gramineus for treatment of the Alzheimer’s disease rat. J. Tradit Chin. Med. 2010, 30, 283–287. [Google Scholar] [CrossRef] [Green Version]
- Olas, B.; Brys, M. Is it safe to use Acorus calamus as a source of promising bioactive compounds in prevention and treatment of cardiovascular diseases? Chem. Biol. Interact. 2018, 281, 32–36. [Google Scholar] [CrossRef]
- Rajput, S.B.; Tonge, M.B.; Karuppayil, S.M. An overview on traditional uses and pharmacological profile of Acorus calamus Linn. (Sweet flag) and other Acorus species. Phytomedicine 2014, 21, 268–276. [Google Scholar] [CrossRef]
- Chellian, R.; Pandy, V.; Mohamed, Z. Pharmacology and toxicology of α-and β-asarone: A review of preclinical evidence. Phytomedicine 2017, 32, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Stegmüller, S.; Schrenk, D.; Cartus, A.T. Formation and fate of DNA adducts of alpha-and beta-asarone in rat hepatocytes. Food Chem. Toxicol. 2018, 116, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Xu, X.; Li, F.; Xie, G.; Zhang, T. Anti-inflammatory treatment with beta-asarone improves impairments in social interaction and cognition in MK-801 treated mice. Brain Res. Bull. 2019, 150, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; He, X.; Han, W.; Chen, X.; Liu, P.; Zhao, X.; Wang, X.; Zhang, L.; Wu, S.; Zheng, X. Genus Tetradium L.: A comprehensive review on traditional uses, phytochemistry, and pharmacological activities. J. Ethnopharmacol. 2019, 231, 337–354. [Google Scholar] [CrossRef] [PubMed]
- Pellati, F.; Benvenuti, S.; Yoshizaki, F.; Bertelli, D.; Rossi, M.C. Headspace solid-phase microextraction-gas chromatography--mass spectrometry analysis of the volatile compounds of Evodia species fruits. J. Chromatogr. A 2005, 1087, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Tundis, R.; Loizzo, M.R.; Menichini, F. An overview on chemical aspects and potential health benefits of limonoids and their derivatives. Crit. Rev. Food Sci. Nutr. 2014, 54, 225–250. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, T.; Zhu, L.; Ma, C.; Wang, S. Anti-ulcerogenic effect of Zuojin Pill against ethanol-induced acute gastric lesion in animal models. J. Ethnopharmacol. 2015, 173, 459–467. [Google Scholar] [CrossRef]
- Shen, Y.; Jin, L.; Xiao, P.; Lu, Y.; Bao, J. Total phenolics, flavonoids, antioxidant capacity in rice grain and their relations to grain color, size and weight. J. Cereal Sci. 2009, 49, 106–111. [Google Scholar] [CrossRef]
- Yu, H.; Zheng, L.; Yin, L.; Xu, L.; Qi, Y.; Han, X.; Xu, Y.; Liu, K.; Peng, J. Protective effects of the total saponins from Dioscorea nipponica Makino against carbon tetrachloride-induced liver injury in mice through suppression of apoptosis and inflammation. Int. Immunopharmacol. 2014, 19, 233–244. [Google Scholar] [CrossRef]
- Lin, J.Y.; Tang, C.Y. Strawberry, loquat, mulberry, and bitter melon juices exhibit prophylactic effects on LPS-induced inflammation using murine peritoneal macrophages. Food Chem. 2008, 107, 1587–1596. [Google Scholar] [CrossRef]
- Lin, W.C.; Lin, J.Y. Five bitter compounds display different anti-inflammatory effects through modulating cytokine secretion using mouse primary splenocytes in vitro. J. Agric. Food Chem. 2011, 59, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.F.; Chiang, B.L.; Lin, J.Y. Amaranthus spinosus water extract directly stimulates proliferation of B lymphocytes in vitro. Int. Immunopharmacol. 2005, 5, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Clery, R.A.; Hammond, C.J. Volatiles from leaves and rhizomes of fragrant Acorus spp. (Acoraceae). Chem. Biodivers. 2008, 5, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, J.; Sureda, A.; Tenore, G.C.; Daglia, M.; Sharifi-Rad, M.; Valussi, M.; Tundis, R.; Sharifi-Rad, M.; Loizzo, M.R.; Ademiluyi, A.O.; et al. Biological activities of essential oils: From plant chemoecology to traditional healing systems. Molecules 2017, 22, 70. [Google Scholar] [CrossRef]
- Dima, C.; Dima, S. Essential oils in foods: Extraction, stabilization, and toxicity. Curr. Opin. Food Sci. 2015, 5, 29–35. [Google Scholar] [CrossRef]
- Bhalla, Y.; Gupt, V.K.; Jaitak, V. Anticancer activity of essential oils: A review. J. Sci. Food Agric. 2013, 93, 3643–3653. [Google Scholar] [CrossRef]
- Dong, R.; Gong, Y.; Meng, W.; Yuan, M.; Zhu, H.; Ying, M.; He, Q.; Cao, J.; Yang, B. The involvement of M2 macrophage polarization inhibition in fenretinide-mediated chemopreventive effects on colon cancer. Cancer Lett. 2017, 388, 43–53. [Google Scholar] [CrossRef]
- Sun, K.; Song, X.; Jia, R.; Yin, Z.; Zou, Y.; Li, L.; Yin, L.; He, C.; Liang, X.; Yue, G.; et al. Evaluation of analgesic and anti-Inflammatory activities of water extract of Galla Chinensis in vivo models. Evid. Based Complement. Alternat. Med. 2018, 6784032. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; You, L.; Yin, X.; Liu, Y.; Leng, X.; Wang, W.; Sai, N.; Ni, J. Heterophyllin B ameliorates lipopolysaccharide-induced inflammation and oxidative stress in RAW 264.7 macrophages by suppressing the PI3K/Akt pathways. Molecules 2018, 23, 717. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Total Flavonoids (mg Quercetin Equivalent/g Sample) | Total Polyphenols (mg Gallic Acid Equivalent/g Sample) | Total Saponins (mg Oleanolic Acid Equivalent/g Sample) |
|---|---|---|---|
| AG | 0.1 ± 0.0 C,b | 17.3 ± 2.3 B,b | 161 ± 32 A,b |
| ER | 20.3 ± 1.6 B,a | 33.3 ± 0.7 B,a | 601 ± 13 A,a |
| NO. | RT | RI | Compounds | M.W. | Chemical Formula | CAS NO. |
|---|---|---|---|---|---|---|
| 1 | 18.619 | 1092.33 | Linalool | 154.14 | C10H18O | 000078-70-6 |
| 2 | 19.54 | 1118 | 4-Isopropyl-1-methyl-2-cyclohexen-1-ol | 154.14 | C10H18O | 029803-81-4 |
| 3 | 20.136 | 1135.1 | Terpenene-1-ol | 154.14 | C10H18O | 000586-82-3 |
| 4 | 21.321 | 1167.66 | 4-Isopropyl-1-methyl-2-cyclohexen-1-ol | 138.10 | C9H14O | 000500-02-7 |
| 5 | 21.539 | 1173.45 | p-Cymen-8-ol | 150.10 | C10H14O | 001197-01-9 |
| 6 | 21.975 | 1184.86 | β-Fenchyl alcohol | 154.14 | C10H18O | 000470-08-6 |
| 7 | 22.344 | 1194.35 | 4-Methyl-1,4-heptadiene | 110.11 | C8H14 | 013857-55-1 |
| 8 | 22.967 | 1212.25 | Trans-(+)-carveol | 152.12 | C10H16O | 001197-07-5 |
| 9 | 23.328 | 1223.12 | Carveol | 152.12 | C10H16O | 000000-00-0 |
| 10 | 23.471 | 1227.38 | Cyclobutanol | 150.1 | C10H14O | 091531-61-2 |
| 11 | 24.201 | 1248.73 | 3-Ethyl-2-pentanone | 114.10 | C7H14O | 006137-03-7 |
| 12 | 24.349 | 1252.98 | 2,5-Dimethyl-1,5-hexadiene-3,4-diol | 142.10 | C8H14O2 | 004723-10-8 |
| 13 | 24.648 | 1261.49 | 3,3,5-Trimethyl-heptane | 142.17 | C10H22 | 007154-80-5 |
| 14 | 25.22 | 1277.48 | 3,3,6-Trimethyl-4,5-heptadien-2-one | 152.12 | C10H16O | 081250-41-1 |
| 15 | 25.325 | 1280.38 | Methyl (2E)-2,5-dimethylhexa-2,4-dienoate | 154.10 | C9H14O2 | 000000-00-0 |
| 16 | 25.421 | 1283.01 | Cuminic alcohol | 150.10 | C10H14O | 000536-60-7 |
| 17 | 25.664 | 1289.65 | Dill ether | 152.12 | C10H16O | 000000-00-0 |
| 18 | 25.846 | 1294.57 | Perilla alcohol | 152.12 | C10H16O | 000536-59-4 |
| 19 | 25.986 | 1298.34 | 4-Methyl-2-(3-methyl-2-butenyl)-furan | 150.10 | C10H14O | 000000-00-0 |
| 20 | 26.787 | 1323.45 | Methyl anthranilate | 151.06 | C8H9NO2 | 000134-20-3 |
| 21 | 27.423 | 1343.13 | Eugenol | 164.08 | C10H12O2 | 000097-53-0 |
| 22 | 27.825 | 1355.33 | (±)-Eldanolide | 168.12 | C10H16O2 | 092843-42-0 |
| 23 | 29.064 | 1391.85 | (±)-Eldanolide | 168.12 | C10H16O2 | 092843-42-0 |
| 24 | 29.863 | 1417.23 | (E)-1-Cyclohexyl-3,3-dimethyl-1-butene | 166.17 | C12H22 | 109660-16-4 |
| 25 | 29.945 | 1419.95 | 1-(2-Hydroxy-4-methoxyphenyl)-ethanone | 166.06 | C9H10O3 | 000552-41-0 |
| 26 | 32.435 | 1499.08 | (E)-3,4-Epoxy-1-(1’,2’-epoxy-3’,3’-epoxymethano-2’,6’,6’-trimethyl-1’-cyclohexyl)-3-methyl-1-butene | 250.16 | C15H22O3 | 091186-32-2 |
| 27 | 32.997 | 1518.69 | β-myrcene | 136.13 | C10H16 | 000127-91-3 |
| 28 | 33.962 | 1551.85 | cis-5-Dodecenoic acid | 198.16 | C12H22O2 | 002430-94-6 |
| 29 | 34.239 | 1561.19 | 3,7,11-Trimethyl-, (Z,E)-1,3,6,10-Dodecatetraene | 204.19 | C15H24 | 026560-14-5 |
| 30 | 35.051 | 1588.15 | (+)-Spathulenol | 220.18 | C15H24O | 077171-55-2 |
| 31 | 35.153 | 1591.49 | 4-Hydroxy-β-ionone | 208.15 | C13H20O2 | 015401-34-0 |
| 32 | 35.474 | 1602.23 | Methyl 4-(4-methyl-3-pentenyl)-3-cyclohexen-1-yl ketone | 206.17 | C14H22O | 038758-04-2 |
| 33 | 35.864 | 1616.64 | 6-Isopropenyl-4,8a-dimethyl-1,2,3,5,6,7,8,8a-octahydro-2-naphthalenol | 220.18 | C15H24O | 000000-00-0 |
| 34 | 36.293 | 1632.31 | (-)-Spathulenol | 220.18 | C15H24O | 077171-55-2 |
| 35 | 36.636 | 1644.7 | Caryophylla-4(12),8(13)-dien-5β-ol | 220.18 | C15H24O | 000000-00-0 |
| 36 | 37.131 | 1662.39 | (+)-β-Costol | 220.18 | C15H24O | 065018-15-7 |
| 37 | 37.604 | 1679.07 | Patchulane | 206.20 | C15H26 | 019078-35-4 |
| 38 | 37.943 | 1690.9 | 2-Pentadecanone | 226.23 | C15H30O | 002345-28-0 |
| 39 | 38.502 | 1711.51 | Cedr-8-en-13-ol | 220.18 | C15H24O | 018319-35-2 |
| 40 | 38.819 | 1723.74 | (E)-2-Methyl-4-(2’,6’,6’-trimethyl-3’-methyliden-1’,2’-epoxy-1’-cyclohexyl)-1,3-butadiene | 218.17 | C15H22O | 077822-46-9 |
| 41 | 38.965 | 1729.33 | Valerenol | 220.18 | C15H24O | 084249-42-3 |
| 42 | 39.203 | 1738.41 | (+)-Valencene | 220.18 | C15H24O | 004630-07-3 |
| 43 | 39.462 | 1748.23 | 7-Isopropenyl-1,4a-dimethyl-4,4a,5,6,7,8-hexahydro-3H-naphthalen-2-one | 218.17 | C15H22O | 000473-08-5 |
| 44 | 40.416 | 1783.85 | 6-Phenyl(deuterate)-2,3,4,5-tetrahydro-3-pyridazinone | 179.11 | C10H5D5N2O | 055999-93-4 |
| 45 | 41.356 | 1820.32 | cis-Z-α-Bisabolene epoxide | 220.18 | C15H24O | 000000-00-0 |
| 46 | 42.828 | 1878.73 | 1-Isopropyl-4,8,12-trimethylcyclotetrtadeca-2,4,7,11-tetraene | 272.25 | C20H32 | 000000-00-0 |
| 47 | 42.917 | 1882.19 | 4,4,8-Trimethyltricyclo[6.3.1.0(1,5)]dodecane-2,9-diol | 238.19 | C15H26O2 | 000000-00-0 |
| 48 | 43.43 | 1902.27 | 7,11-Dimethyl-3-methylene-(Z)-1,6,10-dodecatriene | 204.19 | C15H24 | 028973-97-9 |
| 49 | 44.732 | 1957.07 | 2,6,11,15-Tetramethyl-hexadeca-2,6,8,10,14-pentaene | 272.25 | C20H32 | 038259-79-9 |
| 50 | 45.437 | 1986.09 | 2,4a,5,6,7,8,9,9a-Octahydro-3,5,5-trimethyl-9-methylene-1H-Benzocycloheptene | 204.19 | C15H24 | 080923-88-2 |
| 51 | 45.544 | 1990.45 | 2,6,11,15-Tetramethyl-hexadeca-2,6,8,10,14-pentaene | 272.25 | C20H32 | 038259-79-9 |
| Th1 | Th2 | Th2/Th1 Cytokines Ratio (pg/pg) | ||
|---|---|---|---|---|
| Cytokines (pg/mL) | ||||
| Samples | Treatments (μg/mL) | IL-2 | IL-5 | IL-5/IL-2 |
| AG | control | 23.9 ± 4.5 B | 0.0 ± 0.0 B | 0.00 ± 0.00 E |
| 0.125 | 21.3 ± 3.3 B | 15.3 ± 4.9 A | 0.73 ± 0.26 C,D | |
| 0.25 | 16.8 ± 2.4 C | 13.4 ± 3.4 A | 0.82 ± 0.26 B,C | |
| 1 | 9.7 ± 2.5 D | 12.0 ± 4.0 A | 1.28 ± 0.42 A | |
| 5 | 11.1 ± 2.6 D | 11.8 ± 4.6 A | 1.13 ± 0.59 A,B | |
| LPS | 35.3 ± 5.1 A | 13.4 ± 3.1 A | 0.39 ± 0.12 D | |
| ER | control | 23.9 ± 4.5 b | 0.0 ± 0.0 d | 0.00 ± 0.00 c |
| 0.25 | 9.8 ± 1.6 c | 50.9 ± 6.1 a | 5.50 ± 1.13 b | |
| 1 | 10.3 ± 2.3 c | 49.0 ± 2.5 a,b | 4.90 ± 0.97 b | |
| 5 | 9.4 ± 2.1 c | 50.8 ± 8.3 a | 5.43 ± 1.10 b | |
| 25 | 3.0 ± 1.1 d | 45.3 ± 2.5 b | 16.8 ± 6.72 a | |
| LPS | 35.3 ± 5.1 a | 13.4 ± 3.1 c | 0.39 ± 0.12 c | |
| Pro-Inflammatory | Anti-Inflammatory | Pro-/Anti-Inflammatory Cytokines Secretion Ratios (pg/pg) | |
|---|---|---|---|
| Cytokines (pg/mL) | |||
| Treatments (μg/mL) | TNF-α | IL-10 | TNF-α/IL-10 |
| AG control | 45 ± 20 B | 78 ± 19 B | 0.63 ± 0.34 A |
| 0.125 | 26 ± 5 B | 91 ± 31 B | 0.31 ± 0.07 B |
| 0.25 | 28 ± 7 B | 84 ± 27 B | 0.35 ± 0.16 A,B |
| 1 | 33 ± 5 B | 88 ± 27 B | 0.40 ± 0.04 A,B |
| 5 | 26 ± 4 B | 102 ± 24 B | 0.26 ± 0.04 B |
| LPS | 248 ± 152 A | 735 ± 148 A | 0.33 ± 0.19 A,B |
| ER control | 45 ± 20 b | 78 ± 19 c | 0.63 ± 0.34 a |
| 0.25 | 26 ± 18 b | 108 ± 14 c | 0.25 ± 0.20 b |
| 1 | 25 ± 14 b | 104 ± 9 c | 0.24 ± 0.13 b |
| 5 | 23 ± 11 b | 120 ± 7 c | 0.19 ± 0.08 b |
| 25 | 28 ± 14 b | 339 ± 34 b | 0.08 ± 0.04 b |
| LPS | 248 ± 152 a | 735 ± 148 a | 0.33 ± 0.19 b |
| Pro-Inflammatory | Anti-Inflammatory | Pro-/Anti-Inflammatory Cytokine Secretion Ratios (pg/pg) | |
|---|---|---|---|
| Cytokines (pg/mL) | |||
| Treatments (μg/mL) | TNF-α | IL-10 | TNF-α/IL-10 |
| VC | 144 ± 30 C | 190 ± 63 B | 0.81 ± 0.26 A |
| control (LPS alone) | 499 ± 81 B | 1005 ± 282 A | 0.52 ± 0.14 B |
| AG 0.125 | 592 ± 53 A | 1292 ± 405A | 0.50 ± 0.16 B |
| 0.25 | 580 ± 74 A,B | 1274 ± 371 A | 0.48 ± 0.10 B |
| 1 | 546 ± 68 A,B | 1234 ± 389 A | 0.47 ± 0.14 B |
| 5 | 557 ± 69 A,B | 1308 ± 405 A | 0.46 ± 0.15 B |
| VC | 144 ± 30 d | 190 ± 63 c | 0.81 ± 0.26 a |
| control (LPS alone) | 499 ± 81 a | 1005 ± 282 a | 0.52 ± 0.14 b |
| ER 0.25 | 239 ± 39 c | 726 ± 193 b | 0.34 ± 0.05 c |
| 1 | 222 ± 39 c | 712 ± 120 b | 0.31 ± 0.02 c |
| 5 | 235 ± 49 c | 817 ± 149 a,b | 0.29 ± 0.02 c |
| 25 | 396 ± 47 b | 922 ± 146 a,b | 0.43 ± 0.03 b,c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeh, T.-H.; Lin, J.-Y. Acorus gramineusand and Euodia ruticarpa Steam Distilled Essential Oils Exert Anti-Inflammatory Effects Through Decreasing Th1/Th2 and Pro-/Anti-Inflammatory Cytokine Secretion Ratios In Vitro. Biomolecules 2020, 10, 338. https://doi.org/10.3390/biom10020338
Yeh T-H, Lin J-Y. Acorus gramineusand and Euodia ruticarpa Steam Distilled Essential Oils Exert Anti-Inflammatory Effects Through Decreasing Th1/Th2 and Pro-/Anti-Inflammatory Cytokine Secretion Ratios In Vitro. Biomolecules. 2020; 10(2):338. https://doi.org/10.3390/biom10020338
Chicago/Turabian StyleYeh, Tzu-He, and Jin-Yuarn Lin. 2020. "Acorus gramineusand and Euodia ruticarpa Steam Distilled Essential Oils Exert Anti-Inflammatory Effects Through Decreasing Th1/Th2 and Pro-/Anti-Inflammatory Cytokine Secretion Ratios In Vitro" Biomolecules 10, no. 2: 338. https://doi.org/10.3390/biom10020338