Current Advances in Allosteric Modulation of Muscarinic Receptors

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Advantages of Allosteric Modulators as Therapeutics
2.1. Selectivity by Targeting Less Conserved Domains on the Receptor
2.2. Conservation of Space and Time Pattern of Signaling
2.3. Absolute Selectivity
2.4. Selective Blocking of Activated Receptors
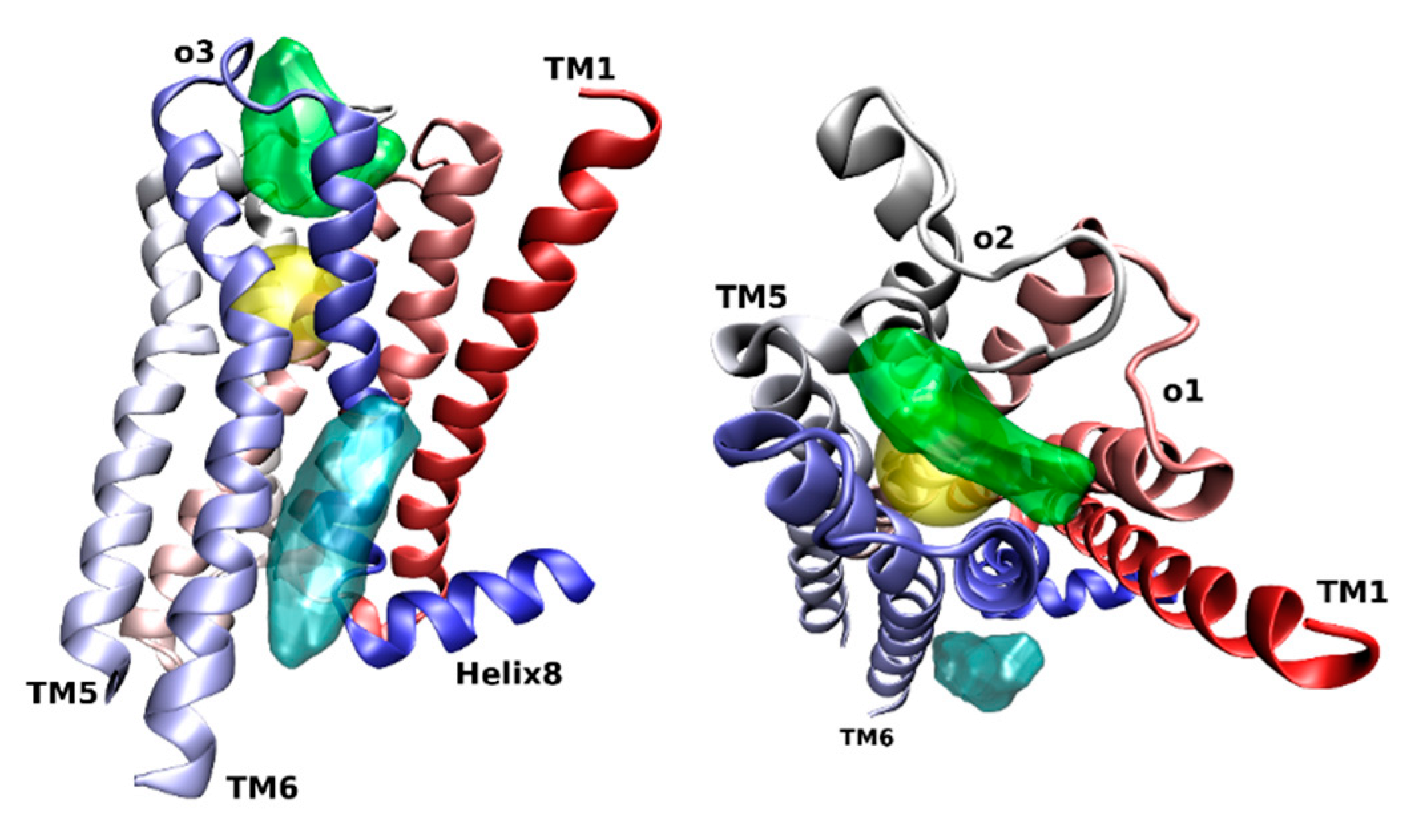
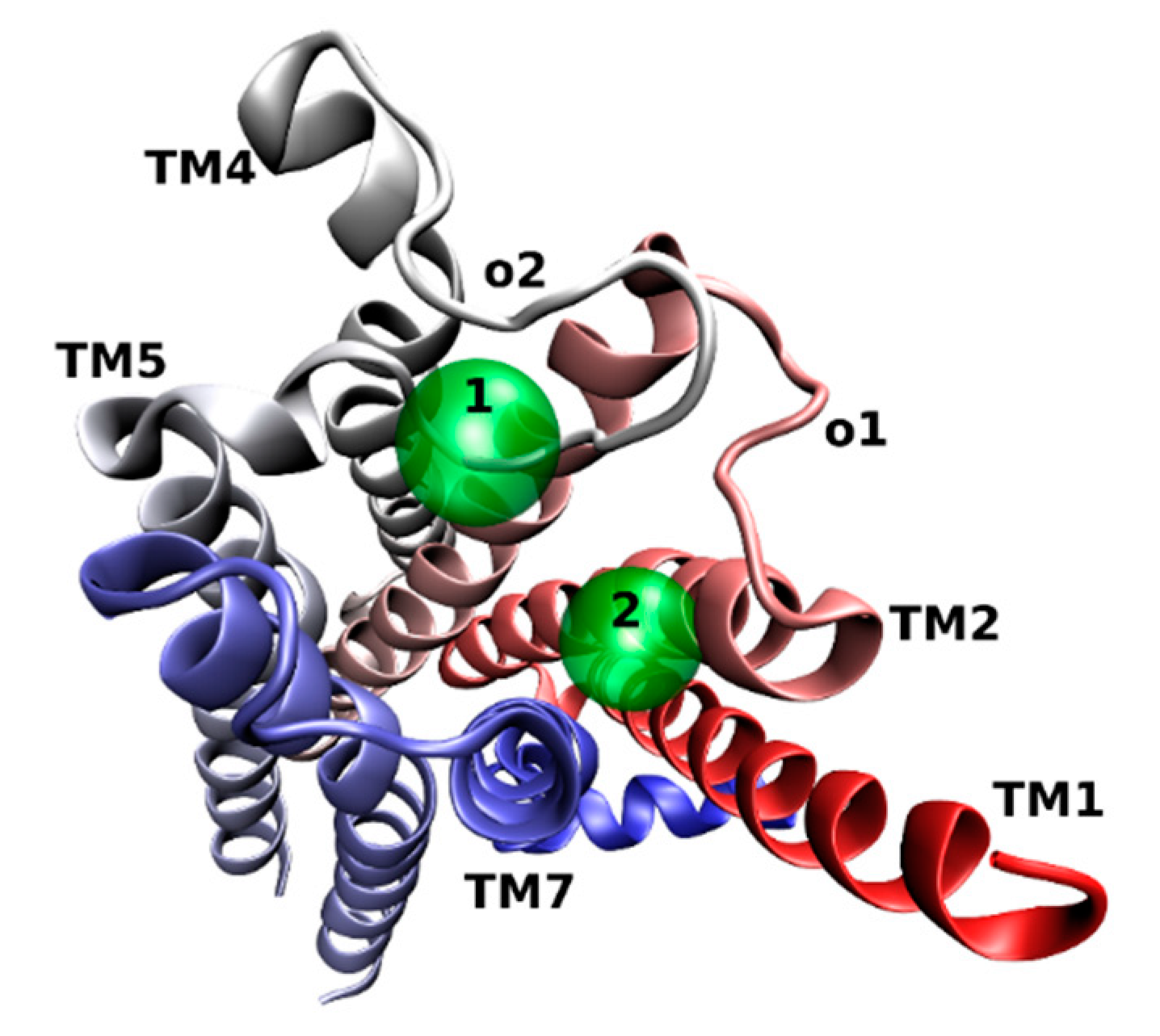
3. Location of the Allosteric Binding Sites on Muscarinic Receptors
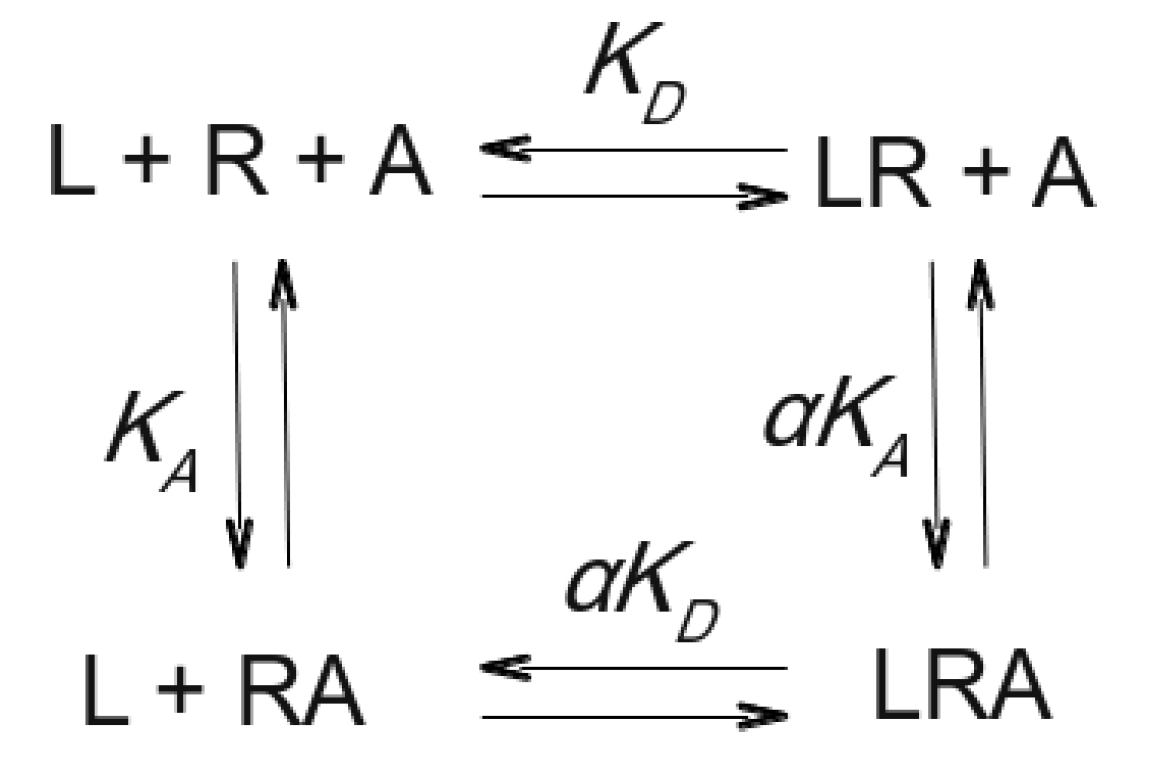
4. Molecular Mechanisms of Action of Allosteric Modulators
5. Role of the Common Allosteric Binding Site in the Binding of Orthosteric Ligands
6. Bitopic Ligands
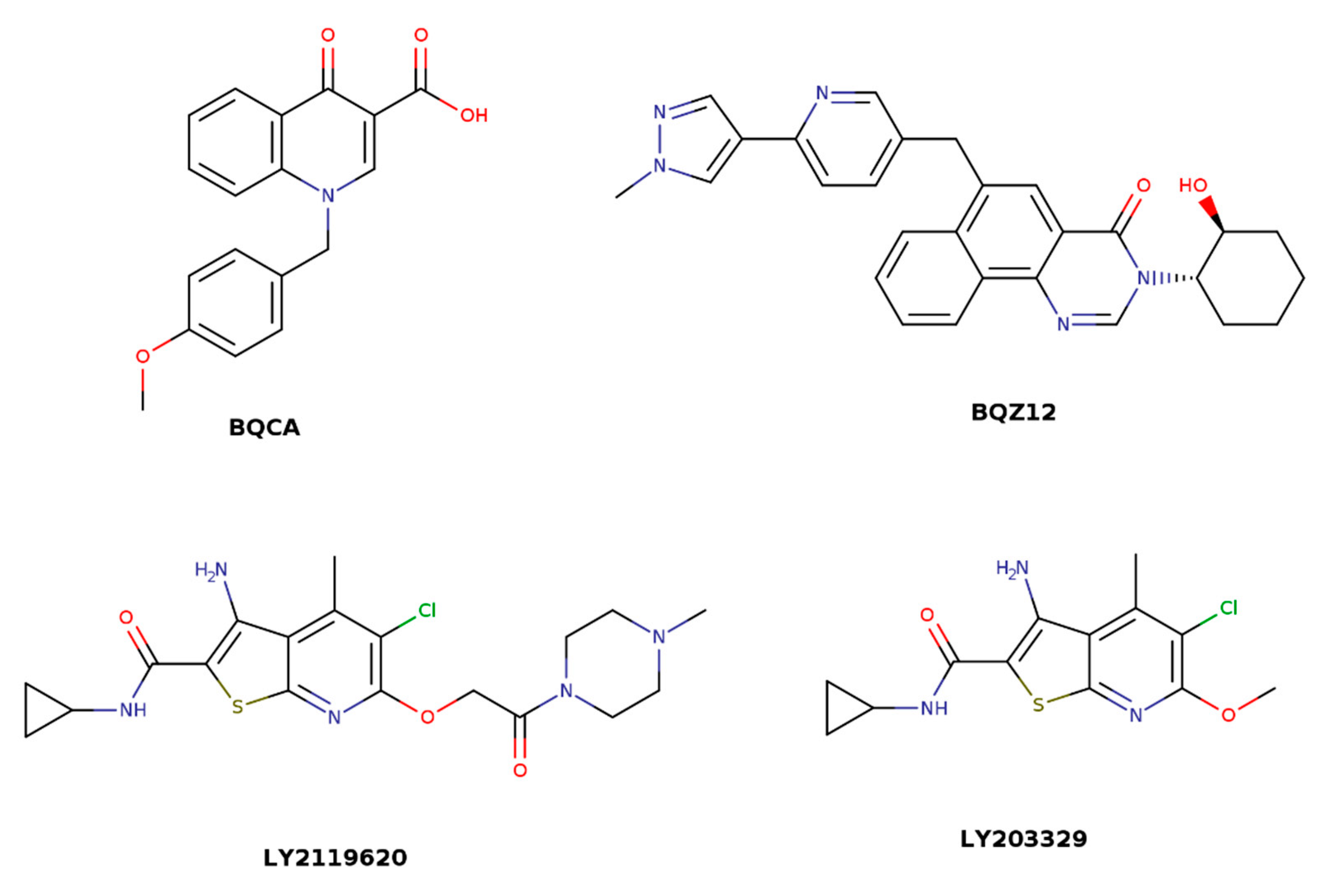
7. Novel Allosteric Modulators
8. Perspectives
Author Contributions
Funding
Conflicts of Interest
Nomenclature
| Compounds | Chemical Names |
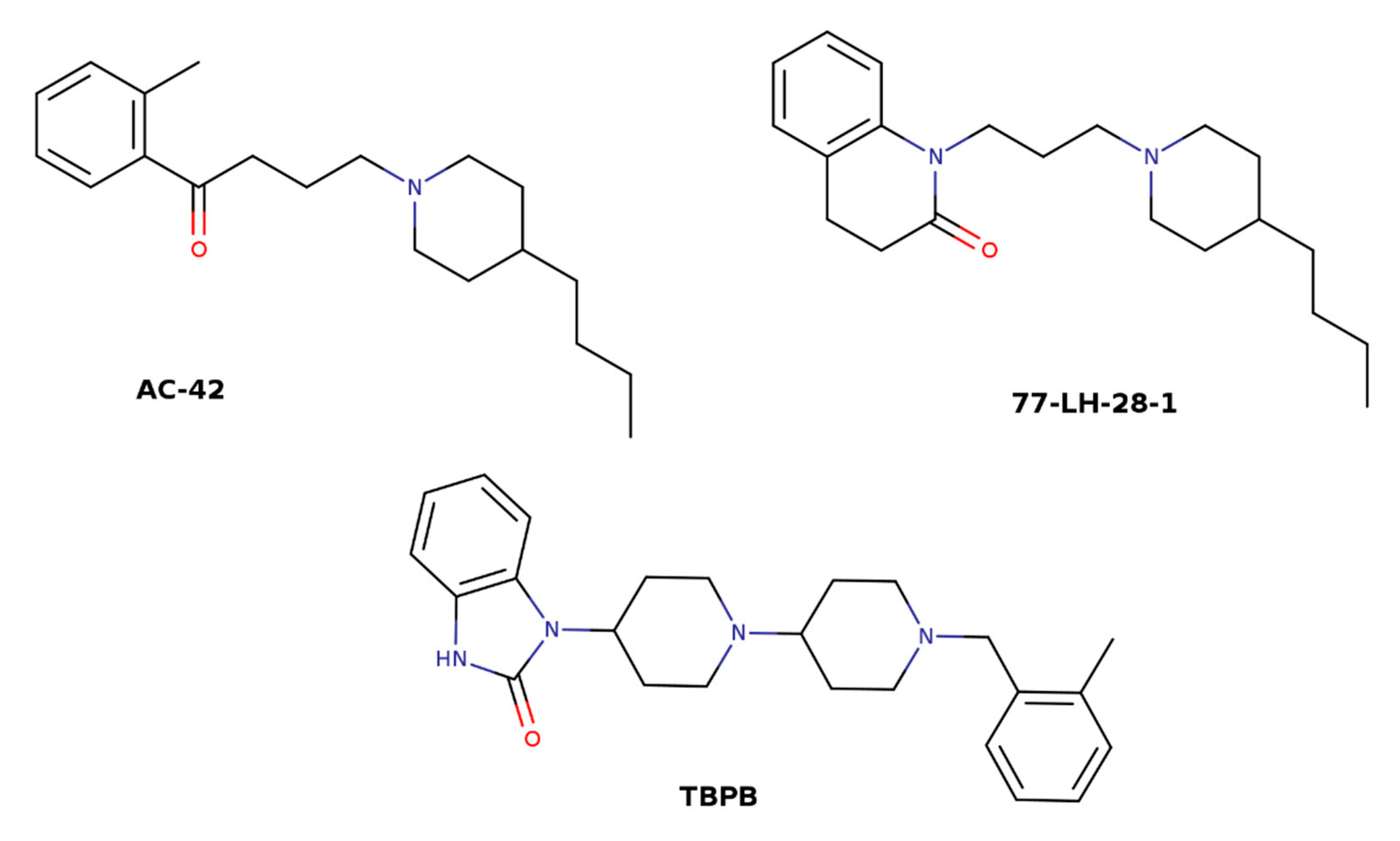
| 77-LH-28-1 | 1-[3-(4-Butylpiperidino)propyl]-3,4-dihydroquinoline-2(1H)-one |
| AC-42 | 4-(4-Butylpiperidin-1-yl)-1-o-tolylbutan-1-one |
| Alcuronium | 2-[(1S,9Z,11S,13S,17S,25Z,27S,33S,35S,36S)-38-(2-hydroxyethylidene)-14,30-bis(prop-2-enyl)-8,24-diaza-14,30-diazoniaundecacyclo [2 5.5.2.211,14.11,26.110,17.02,7.013,17.018,23.030,33.08,35.024,36]octatriaconta-2,4,6,9,18,20,22,25-octaen-28-ylidene]ethanol |
| BQCA | 1-(4-Methoxybenzyl)-4-oxo-1,4-dihydroquinoline-3-carboxylic acid |
| BQZ12 | 3-((1S,2S)-2-hydroxycyclohexyl)-6-((6-(1-methyl-1H-pyrazol-4-yl)pyridin-3-yl)methyl)benzo [h]quinazolin-4(3H)-one |
| Gallamine | 2-[2,3-bis [2-(triethylazaniumyl)ethoxy]phenoxy]ethyl-triethylazanium |
| LY2119620 | 3-Amino-5-chloro-N-cyclopropyl-4-methyl-6-[2-(4-methylpiperazin-1-yl)-2-oxoethoxy]thieno [2,3-b]pyridine-2-carboxamide |
| LY2033298 | 3-Amino-5-Chloro-N-Cyclopropyl-6-Methoxy-4-Methylthieno [2,3-B]Pyridine-2-Carboxamide |
| McN-A-343 | 4-(m-chlorophenyl-carbamoyloxy)-2-butynyltri-methylammonium |
| Methoctramine | N,N′-bis [6-[[(2-methoxyphenyl)-methyl]hexyl]-1,8-octane]diamine |
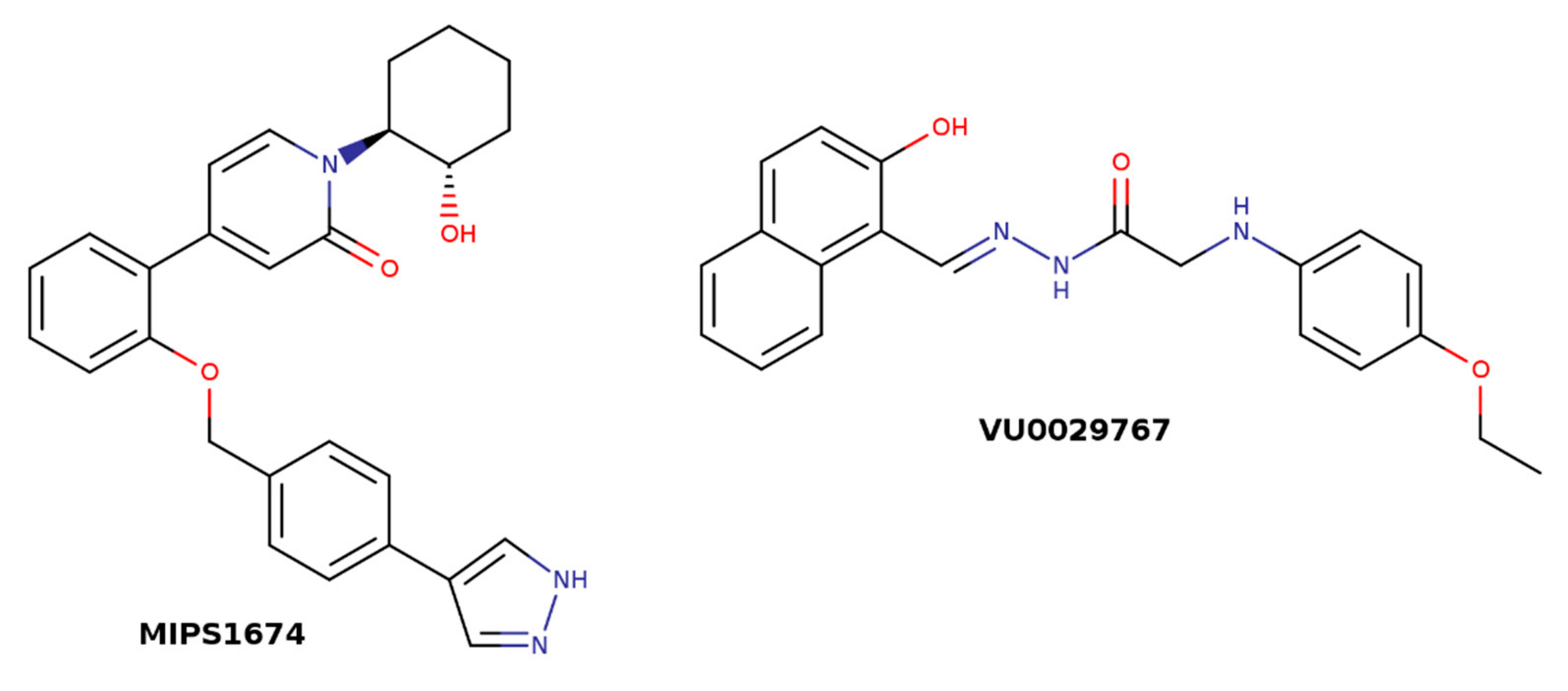
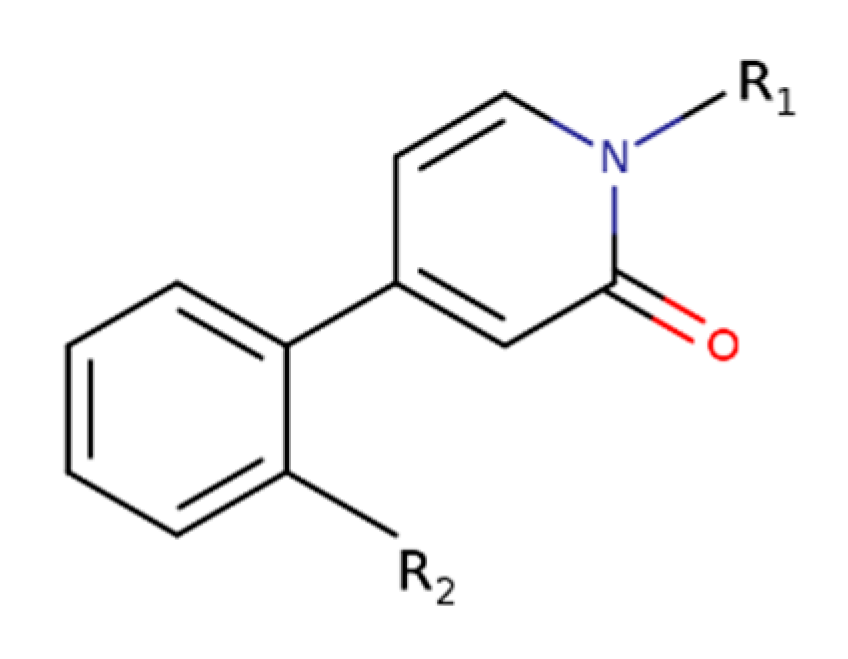
| MIPS1674 | 1-[(1S,2S)-2-hydroxycyclohexyl]-4-[2-[[4-(1H-pyrazol-4-yl)phenyl]methoxy]phenyl]pyridin-2-one |
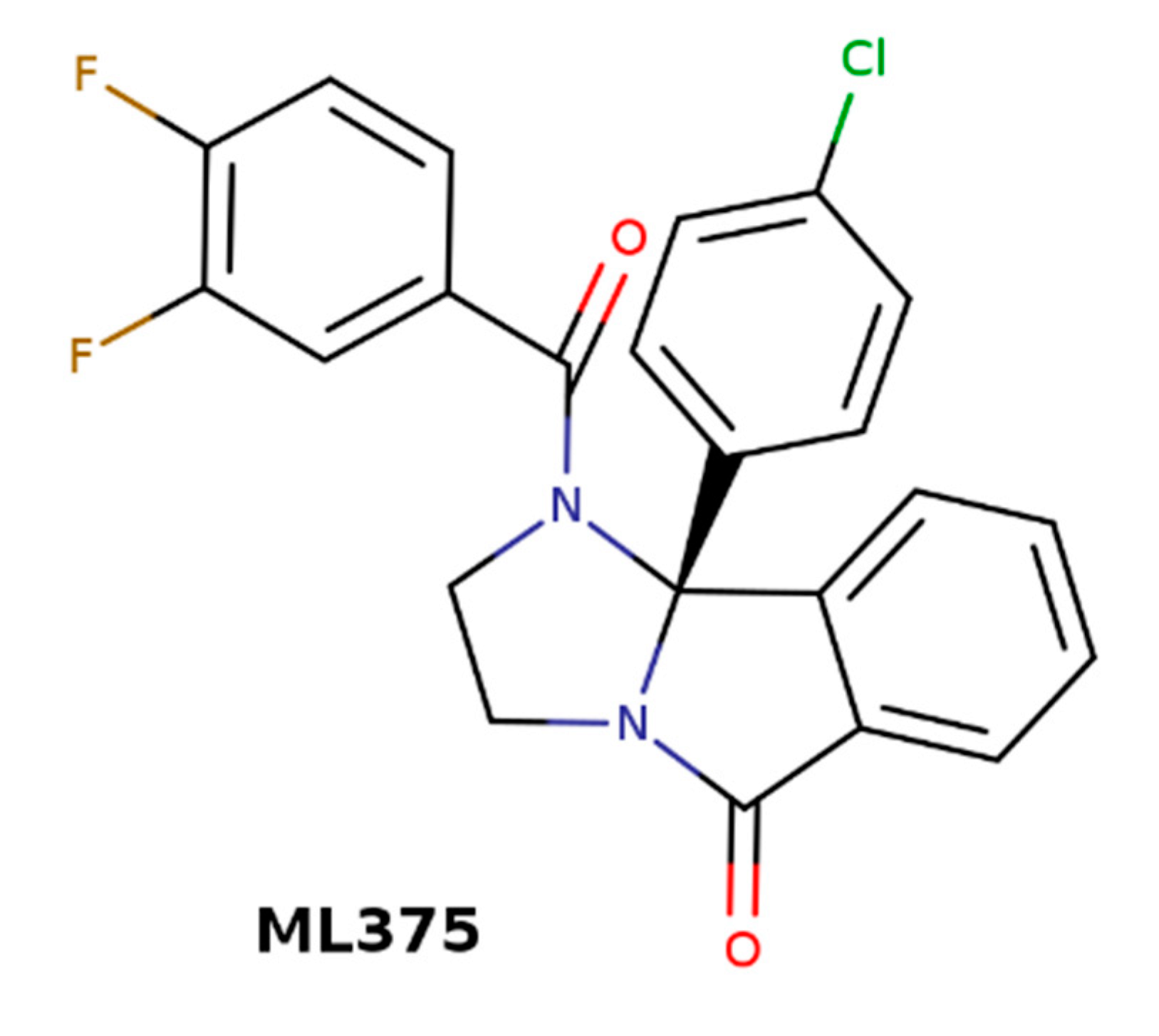
| ML375 | (S)-9b-(4-chlorophenyl)-1-(3,4-difluorobenzoyl)-2,3- dihydro-1H-imidazo [2,1-a]isoindol-5(9bH)-one |
| Naphmethonium | [3-(1,3-dioxobenzo [de]isoquinolin-2-yl)-2,2-dimethylpropyl]-[6-[3-(1,3-dioxoisoindol-2-yl)propyl-dimethylazaniumyl]hexyl]-dimethylazanium |
| NDMC | N-desmethylclozapine; 3-chloro-6-piperazin-1-yl-11H-benzo [b] [1,4]benzodiazepine |
| NMS | N-methylscopolamine; [(1S,2S,4R,5R)-9,9-dimethyl-3-oxa-9-azoniatricyclo [3.3.1.02,4]nonan-7-yl] (2S)-3-hydroxy-2-phenylpropanoate |
| Strychnine | (4aR,5aS,8aR,13aS,15aS,15bR)-4a,5,5a,7,8,13a,15,15a,15b,16-decahydro-2H-4,6-methanoindolo [3,2,1-ij]oxepino [2,3,4-de]pyrrolo [2,3-h]quinolin-14-one |
| TBPB | 1-(1′-(2-methylbenzyl)-1,4′-bipiperidin-4-yl)-1H-benzo [d]imidazol-2(3H)-one |
| Tiotropium | [(1S,2S,4R,5R)-9,9-dimethyl-3-oxa-9-azoniatricyclo [3.3.1.02,4]nonan-7-yl]2-hydroxy-2,2-dithiophen-2-ylacetate |
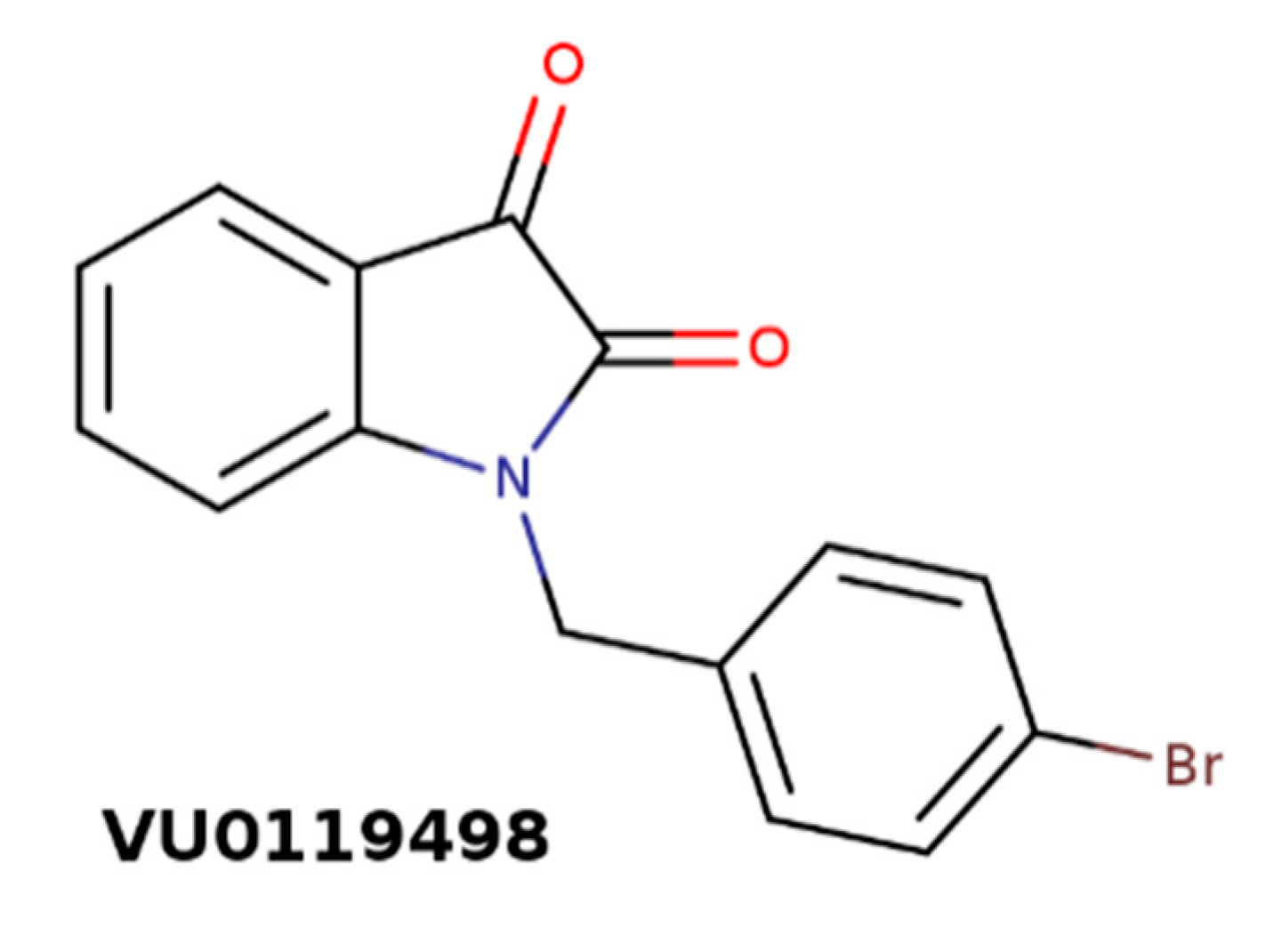
| VU0119498 | 1-(4-bromobenzyl)indole-2,3-dione |
| VU0029767 | (E)-2-(4-ethoxyphenylamino)-N′-((2-hydroxynaphthalen-1-yl)methylene)acetohydrazide |
| W84 | 6-[dimethyl-[3-(4-methyl-1,3-dioxoisoindol-2-yl)propyl]azaniumyl]hexyl-dimethyl-[3-(4-methyl-1,3-dioxoisoindol-2-yl)propyl]azanium |
References
- Bonner, T.I.; Buckley, N.J.; Young, A.C.; Brann, M.R. Identification of a family of muscarinic acetylcholine receptor genes. Science 1987, 237, 527–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haga, K.; Kruse, A.C.; Asada, H.; Yurugi-Kobayashi, T.; Shiroishi, M.; Zhang, C.; Weis, W.I.; Okada, T.; Kobilka, B.K.; Haga, T.; et al. Structure of the human M2 muscarinic acetylcholine receptor bound to an antagonist. Nature 2012, 482, 547–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruse, A.C.; Hu, J.; Pan, A.C.; Arlow, D.H.; Rosenbaum, D.M.; Rosemond, E.; Green, H.F.; Liu, T.; Chae, P.S.; Dror, R.O.; et al. Structure and dynamics of the M3 muscarinic acetylcholine receptor. Nature 2012, 482, 552–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruse, A.C.; Ring, A.M.; Manglik, A.; Hu, J.; Hu, K.; Eitel, K.; Hübner, H.; Pardon, E.; Valant, C.; Sexton, P.M.; et al. Activation and allosteric modulation of a muscarinic acetylcholine receptor. Nature 2013, 504, 101–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thal, D.M.; Sun, B.; Feng, D.; Nawaratne, V.; Leach, K.; Felder, C.C.; Bures, M.G.; Evans, D.A.; Weis, W.I.; Bachhawat, P.; et al. Crystal structures of the M1 and M4 muscarinic acetylcholine receptors. Nature 2016, 531, 335–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuckovic, Z.; Gentry, P.R.; Berizzi, A.E.; Hirata, K.; Varghese, S.; Thompson, G.; van der Westhuizen, E.T.; Burger, W.A.C.; Rahmani, R.; Valant, C.; et al. Crystal structure of the M5 muscarinic acetylcholine receptor. Proc. Natl. Acad. Sci. USA 2019, 116, 26001–26007. [Google Scholar] [CrossRef] [Green Version]
- Eglen, R.M. Overview of muscarinic receptor subtypes. In Handb Exp Pharmacol; Springer: Heidelberg, Germany, 2012; pp. 3–28. ISBN 0171-2004. [Google Scholar]
- Conn, P.J.; Jones, C.K.; Lindsley, C.W. Subtype-selective allosteric modulators of muscarinic receptors for the treatment of CNS disorders. Trends Pharmacol Sci 2009, 30, 148–155. [Google Scholar] [CrossRef] [Green Version]
- Bock, A.; Schrage, R.; Mohr, K. Allosteric modulators targeting CNS muscarinic receptors. Neuropharmacology 2018, 136, 427–437. [Google Scholar] [CrossRef] [Green Version]
- Gautam, D.; Han, S.-J.; Duttaroy, A.; Mears, D.; Hamdan, F.F.; Li, J.H.; Cui, Y.; Jeon, J.; Wess, J. Role of the M3 muscarinic acetylcholine receptor in beta-cell function and glucose homeostasis. Diabetes Obes. Metab. 2007, 9, 158–169. [Google Scholar] [CrossRef]
- Gosens, R.; Zaagsma, J.; Meurs, H.; Halayko, A.J. Muscarinic receptor signaling in the pathophysiology of asthma and COPD. Respir. Res. 2006, 7, 73. [Google Scholar] [CrossRef] [Green Version]
- Hegde, S.S. Muscarinic receptors in the bladder: From basic research to therapeutics. Br. J. Pharmacol. 2006, 147, S80–S87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monod, J.; Wyman, J.; Changeux, J.P. On the nature of allosteric transitions: A plausible model. J. Mol. Biol. 1965, 12, 88–118. [Google Scholar] [CrossRef]
- Koshland, D.E.; Némethy, G.; Filmer, D. Comparison of experimental binding data and theoretical models in proteins containing subunits. Biochemistry 1966, 5, 365–385. [Google Scholar] [CrossRef]
- Jakubík, J.; El-Fakahany, E.E. Allosteric Modulation of Muscarinic Receptors. In Muscarinic Receptor: From Structure to Animal Models; Myslivecek, J., Jakubik, J., Eds.; Neuromethods; Humana Press; Springer Science + Business Media LLC: New York, NY, USA, 2016; Volume 107, pp. 95–130. ISBN 978-1-4939-287-6. [Google Scholar]
- Kenakin, T.; Strachan, R.T. PAM-Antagonists: A Better Way to Block Pathological Receptor Signaling? Trends Pharmacol. Sci. 2018, 39, 748–765. [Google Scholar] [CrossRef] [PubMed]
- Jakubík, J.; Bacáková, L.; Lisá, V.; El-Fakahany, E.E.; Tucek, S. Activation of muscarinic acetylcholine receptors via their allosteric binding sites. Proc. Natl. Acad. Sci. USA 1996, 93, 8705–8709. [Google Scholar] [CrossRef] [Green Version]
- Lüllmann, H.; Ohnesorge, F.K.; Schauwecker, G.C.; Wassermann, O. Inhibition of the actions of carbachol and DFP on guinea pig isolated atria by alkane-bis-ammonium compounds. Eur. J. Pharmacol. 1969, 6, 241–247. [Google Scholar] [CrossRef]
- Clark, A.L.; Mitchelson, F. The inhibitory effect of gallamine on muscarinic receptors. Br. J. Pharmacol. 1976, 58, 323–331. [Google Scholar] [CrossRef] [Green Version]
- Jakubík, J.; El-Fakahany, E.E. Allosteric Modulation of Muscarinic Acetylcholine Receptors. Pharmaceuticals 2010, 3, 2838–2860. [Google Scholar] [CrossRef]
- Mash, D.C.; Flynn, D.D.; Potter, L.T. Loss of M2 muscarine receptors in the cerebral cortex in Alzheimer’s disease and experimental cholinergic denervation. Science 1985, 228, 1115–1117. [Google Scholar] [CrossRef]
- Perry, E.K.; Smith, C.J.; Court, J.A.; Perry, R.H. Cholinergic nicotinic and muscarinic receptors in dementia of Alzheimer, Parkinson and Lewy body types. J. Neural Transm. Park. Dis. Dement. Sect. 1990, 2, 149–158. [Google Scholar] [CrossRef]
- Buels, K.S.; Fryer, A.D. Muscarinic receptor antagonists: Effects on pulmonary function. Handb. Exp. Pharmacol. 2012, 208, 317–341. [Google Scholar]
- Krejčí, A.; Tuček, S. Changes of cooperativity between N-methylscopolamine and allosteric modulators alcuronium and gallamine induced by mutations of external loops of muscarinic M(3) receptors. Mol. Pharmacol. 2001, 60, 761–767. [Google Scholar]
- Huang, X.-P.; Prilla, S.; Mohr, K.; Ellis, J. Critical amino acid residues of the common allosteric site on the M2 muscarinic acetylcholine receptor: more similarities than differences between the structurally divergent agents gallamine and bis(ammonio)alkane-type hexamethylene-bis-[dimethyl-(3-phthalimidopropyl)ammonium]dibromide. Mol. Pharmacol. 2005, 68, 769–778. [Google Scholar] [PubMed] [Green Version]
- Jakubík, J.; Krejcí, A.; Dolezal, V. Asparagine, valine, and threonine in the third extracellular loop of muscarinic receptor have essential roles in the positive cooperativity of strychnine-like allosteric modulators. J. Pharmacol. Exp. Ther. 2005, 313, 688–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leppik, R.A.; Miller, R.C.; Eck, M.; Paquet, J.L. Role of acidic amino acids in the allosteric modulation by gallamine of antagonist binding at the m2 muscarinic acetylcholine receptor. Mol. Pharmacol. 1994, 45, 983–990. [Google Scholar]
- Dror, R.O.; Green, H.F.; Valant, C.; Borhani, D.W.; Valcourt, J.R.; Pan, A.C.; Arlow, D.H.; Canals, M.; Lane, J.R.; Rahmani, R.; et al. Structural basis for modulation of a G-protein-coupled receptor by allosteric drugs. Nature 2013, 503, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Ridha, A.; López, L.; Keov, P.; Thal, D.M.; Mistry, S.N.; Sexton, P.M.; Lane, J.R.; Canals, M.; Christopoulos, A. Molecular determinants of allosteric modulation at the M1 muscarinic acetylcholine receptor. J. Biol. Chem. 2014, 289, 6067–6079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawaratne, V.; Leach, K.; Felder, C.C.; Sexton, P.M.; Christopoulos, A. Structural determinants of allosteric agonism and modulation at the M4 muscarinic acetylcholine receptor: Identification of ligand-specific and global activation mechanisms. J. Biol. Chem. 2010, 285, 19012–19021. [Google Scholar] [CrossRef] [Green Version]
- Hollingsworth, S.A.; Kelly, B.; Valant, C.; Michaelis, J.A.; Mastromihalis, O.; Thompson, G.; Venkatakrishnan, A.J.; Hertig, S.; Scammells, P.J.; Sexton, P.M.; et al. Cryptic pocket formation underlies allosteric modulator selectivity at muscarinic GPCRs. Nat. Commun. 2019, 10, 3289. [Google Scholar] [CrossRef]
- Ballesteros, J.A.; Weinstein, H. Integrated methods for the construction of three dimensional models and computational probing of structure function relations in G protein-coupled receptors. In Methods in Neurosciences; Sealfon, S.C., Conn, P.M., Eds.; Academic Press: San Diego, CA, USA, 1995; Volume 25, pp. 366–428. ISBN 9780121852955. [Google Scholar]
- Isberg, V.; De Graaf, C.; Bortolato, A.; Cherezov, V.; Katritch, V.; Marshall, F.H.; Mordalski, S.; Pin, J.-P.P.; Stevens, R.C.; Vriend, G.; et al. Generic GPCR residue numbers—Aligning topology maps while minding the gaps. Trends Pharmacol. Sci. 2015, 36, 22–31. [Google Scholar] [CrossRef] [Green Version]
- Abdul-Ridha, A.; Lane, J.R.; Mistry, S.N.; López, L.; Sexton, P.M.; Scammells, P.J.; Christopoulos, A.; Canals, M. Mechanistic insights into allosteric structure-function relationships at the M1 muscarinic acetylcholine receptor. J. Biol. Chem. 2014, 289, 33701–33711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradley, S.J.; Bourgognon, J.M.; Sanger, H.E.; Verity, N.; Mogg, A.J.; White, D.J.; Butcher, A.J.; Moreno, J.A.; Molloy, C.; Macedo-Hatch, T.; et al. M1 muscarinic allosteric modulators slow prion neurodegeneration and restore memory loss. J. Clin. Investig. 2016, 127, 487–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazareno, S.; Popham, A.; Birdsall, N.J.M. Analogs of WIN 62,577 define a second allosteric site on muscarinic receptors. Mol. Pharmacol. 2002, 62, 1492–1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michal, P.; Rudajev, V.; El-Fakahany, E.E.; Doležal, V. Membrane cholesterol content influences binding properties of muscarinic M2 receptors and differentially impacts activation of second messenger pathways. Eur. J. Pharmacol. 2009, 606, 50–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michal, P.; El-Fakahany, E.E.; Doležal, V. Changes in Membrane Cholesterol Differentially Influence Preferential and Non-preferential Signaling of the M1 and M3 Muscarinic Acetylcholine Receptors. Neurochem. Res. 2014, 40, 2068–2077. [Google Scholar] [CrossRef] [Green Version]
- Janickova, H.; Rudajev, V.; Dolejsi, E.; Koivisto, H.; Jakubik, J.; Tanila, H.; El-Fakahany, E.E.; Dolezal, V. Lipid-Based Diets Improve Muscarinic Neurotransmission in the Hippocampus of Transgenic APPswe/PS1dE9 Mice. Curr. Alzheimer Res. 2015, 12, 923–931. [Google Scholar] [CrossRef]
- Randáková, A.; Dolejší, E.; Rudajev, V.; Zimčík, P.; Doležal, V.; El-Fakahany, E.E.; Jakubík, J. Role of membrane cholesterol in differential sensitivity of muscarinic receptor subtypes to persistently bound xanomeline. Neuropharmacology 2018, 133, 129–144. [Google Scholar] [CrossRef]
- Gimpl, G. Interaction of G protein coupled receptors and cholesterol. Chem. Phys. Lipids 2016, 199, 61–73. [Google Scholar] [CrossRef]
- Lee, A.G. Interfacial Binding Sites for Cholesterol on G Protein-Coupled Receptors. Biophys. J. 2019, 116, 1586–1597. [Google Scholar] [CrossRef]
- Zlotos, D.P.; Buller, S.; Stiefl, N.; Baumann, K.; Mohr, K. Probing the pharmacophore for allosteric ligands of muscarinic M2 receptors: SAR and QSAR studies in a series of bisquaternary salts of caracurine V and related ring systems. J. Med. Chem. 2004, 47, 3561–3571. [Google Scholar] [CrossRef]
- Bermudez, M.; Wolber, G. Structure versus function—The impact of computational methods on the discovery of specific GPCR-ligands. Bioorganic Med. Chem. 2015, 23, 3907–3912. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros, J.A.; Weinstein, H. Integrated methods for the construction of three-dimensional models and computational probing of structure-function relations in G protein-coupled receptors. Methods Neurosci. 1995, 25, 366–428. [Google Scholar]
- Burger, W.A.C.; Sexton, P.M.; Christopoulos, A.; Thal, D.M. Toward an understanding of the structural basis of allostery in muscarinic acetylcholine receptors. J. Gen. Physiol. 2018, 150, 1360–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laugwitz, K.L.; Allgeier, A.; Offermanns, S.; Spicher, K.; Van Sande, J.; Dumont, J.E.; Schultz, G. The human thyrotropin receptor: A heptahelical receptor capable of stimulating members of all four G protein families. Proc. Natl. Acad. Sci. USA 1996, 93, 116–120. [Google Scholar] [CrossRef] [Green Version]
- Hermans, E. Biochemical and pharmacological control of the multiplicity of coupling at G-protein-coupled receptors. Pharmacol. Ther. 2003, 99, 25–44. [Google Scholar] [CrossRef]
- Jakubik, J.; El-Fakahany, E.E.; Doležal, V. Differences in kinetics of xanomeline binding and selectivity of activation of G proteins at M(1) and M(2) muscarinic acetylcholine receptors. Mol. Pharmacol. 2006, 70, 656–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenakin, T.; Christopoulos, A. Signalling bias in new drug discovery: detection, quantification and therapeutic impact. Nat. Rev. Drug Discov. 2013, 12, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Marlo, J.E.; Niswender, C.M.; Days, E.L.; Bridges, T.M.; Xiang, Y.; Rodriguez, A.L.; Shirey, J.K.; Brady, A.E.; Nalywajko, T.; Luo, Q.; et al. Discovery and characterization of novel allosteric potentiators of M1 muscarinic receptors reveals multiple modes of activity. Mol. Pharmacol. 2009, 75, 577–588. [Google Scholar] [CrossRef] [Green Version]
- Van der Westhuizen, E.T.; Spathis, A.; Khajehali, E.; Jörg, M.; Mistry, S.N.; Capuano, B.; Tobin, A.B.; Sexton, P.M.; Scammells, P.J.; Valant, C.; et al. Assessment of the Molecular Mechanisms of Action of Novel 4-Phenylpyridine-2-One and 6-Phenylpyrimidin-4-One Allosteric Modulators at the M1 Muscarinic Acetylcholine Receptors. Mol. Pharmacol. 2018, 94, 770–783. [Google Scholar] [CrossRef] [Green Version]
- Wacker, D.; Stevens, R.C.; Roth, B.L. How Ligands Illuminate GPCR Molecular Pharmacology. Cell 2017, 170, 414–427. [Google Scholar] [CrossRef] [Green Version]
- Bermudez, M.; Bock, A. Does Divergent Binding Pocket Closure Drive Ligand Bias for Class A GPCRs? Trends Pharmacol. Sci. 2019, 40, 236–239. [Google Scholar] [CrossRef] [PubMed]
- Jakubik, J.; El-Fakahany, E.E.; Tucek, S. Evidence for a tandem two-site model of ligand binding to muscarinic acetylcholine receptors. J. Biol. Chem. 2000, 275, 18836–18844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Järv, J.; Hedlund, B.; Bartfai, T. Isomerization of the muscarinic receptor. antagonist complex. J. Biol. Chem. 1979, 254, 5595–5598. [Google Scholar] [PubMed]
- Järv, J.; Hedlund, B.; Bartfai, T. Kinetic studies on muscarinic antagonist-agonist competition. J. Biol. Chem. 1980, 255, 2649–2651. [Google Scholar]
- Redka, D.S.; Pisterzi, L.F.; Wells, J.W. Binding of orthosteric ligands to the allosteric site of the M(2) muscarinic cholinergic receptor. Mol. Pharmacol. 2008, 74, 834–843. [Google Scholar] [CrossRef] [Green Version]
- Jakubík, J.; Randáková, A.; Zimčík, P.; El-Fakahany, E.E.; Doležal, V. Binding of N-methylscopolamine to the extracellular domain of muscarinic acetylcholine receptors. Sci. Rep. 2017, 7, 40381. [Google Scholar] [CrossRef] [Green Version]
- Kistemaker, L.E.M.; Elzinga, C.R.S.; Tautermann, C.S.; Pieper, M.P.; Seeliger, D.; Alikhil, S.; Schmidt, M.; Meurs, H.; Gosens, R. Second M3 muscarinic receptor binding site contributes to bronchoprotection by tiotropium. Br. J. Pharmacol. 2019, 176, 2864–2876. [Google Scholar] [CrossRef]
- Jakubík, J.; Zimčík, P.; Randáková, A.; Fuksová, K.; El-Fakahany, E.E.; Doležal, V. Molecular mechanisms of methoctramine binding and selectivity at muscarinic acetylcholine receptors. Mol. Pharmacol. 2014, 86, 180–192. [Google Scholar] [CrossRef] [Green Version]
- Bock, A.; Bermudez, M.; Krebs, F.; Matera, C.; Chirinda, B.; Sydow, D.; Dallanoce, C.; Holzgrabe, U.; De Amici, M.; Lohse, M.J.; et al. Ligand binding ensembles determine graded agonist efficacies at a G protein-coupled receptor. J. Biol. Chem. 2016, 291, 16375–16389. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, J.; Van Der Mey, D.; Bermudez, M.; Klöckner, J.; Schrage, R.; Kostenis, E.; Tränkle, C.; Wolber, G.; Mohr, K.; Holzgrabe, U. Dualsteric muscarinic antagonists-orthosteric binding pose controls allosteric subtype selectivity. J. Med. Chem. 2014, 57, 6739–6750. [Google Scholar] [CrossRef]
- Avlani, V.A.; Langmead, C.J.; Guida, E.; Wood, M.D.; Tehan, B.G.; Herdon, H.J.; Watson, J.M.; Sexton, P.M.; Christopoulos, A. Orthosteric and allosteric modes of interaction of novel selective agonists of the M1 muscarinic acetylcholine receptor. Mol. Pharmacol. 2010, 78, 94–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keov, P.; Valant, C.; Devine, S.M.; Lane, J.R.; Scammells, P.J.; Sexton, P.M.; Christopoulos, A. Reverse engineering of the selective agonist TBPB unveils both orthosteric and allosteric modes of action at the M₁ muscarinic acetylcholine receptor. Mol. Pharmacol 2013, 84, 425–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregory, K.J.; Hall, N.E.; Tobin, A.B.; Sexton, P.M.; Christopoulos, A. Identification of orthosteric and allosteric site mutations in M2 muscarinic acetylcholine receptors that contribute to ligand-selective signaling bias. J. Biol. Chem. 2010, 285, 7459–7474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valant, C.; Gregory, K.J.; Hall, N.E.; Scammells, P.J.; Lew, M.J.; Sexton, P.M.; Christopoulos, A. A novel mechanism of G protein-coupled receptor functional selectivity. Muscarinic partial agonist McN-A-343 as a bitopic orthosteric/allosteric ligand. J. Biol Chem 2008, 283, 29312–29321. [Google Scholar] [CrossRef] [Green Version]
- Agnetta, L.; Kauk, M.; Canizal, M.C.A.; Messerer, R.; Holzgrabe, U.; Hoffmann, C.; Decker, M. A Photoswitchable Dualsteric Ligand Controlling Receptor Efficacy. Angew. Chem. Int. Ed. 2017, 56, 7282–7287. [Google Scholar] [CrossRef]
- Matera, C.; Flammini, L.; Quadri, M.; Vivo, V.; Ballabeni, V.; Holzgrabe, U.; Mohr, K.; De Amici, M.; Barocelli, E.; Bertoni, S.; et al. Bis(ammonio)alkane-type agonists of muscarinic acetylcholine receptors: synthesis, in vitro functional characterization, and in vivo evaluation of their analgesic activity. Eur. J. Med. Chem. 2014, 75, 222–232. [Google Scholar] [CrossRef]
- Cristofaro, I.; Spinello, Z.; Matera, C.; Fiore, M.; Conti, L.; De Amici, M.; Dallanoce, C.; Tata, A.M. Activation of M2 muscarinic acetylcholine receptors by a hybrid agonist enhances cytotoxic effects in GB7 glioblastoma cancer stem cells. Neurochem. Int. 2018, 118, 52–60. [Google Scholar] [CrossRef]
- Jakubík, J.; Bacáková, L.; El-Fakahany, E.E.; Tucek, S. Positive cooperativity of acetylcholine and other agonists with allosteric ligands on muscarinic acetylcholine receptors. Mol. Pharmacol. 1997, 52, 172–179. [Google Scholar] [CrossRef] [Green Version]
- Birdsall, N.J.; Farries, T.; Gharagozloo, P.; Kobayashi, S.; Kuonen, D.; Lazareno, S.; Popham, A.; Sugimoto, M. Selective allosteric enhancement of the binding and actions of acetylcholine at muscarinic receptor subtypes. Life Sci. 1997, 60, 1047–1052. [Google Scholar] [CrossRef]
- Bridges, T.M.; Marlo, J.E.; Niswender, C.M.; Jones, C.K.; Jadhav, S.B.; Gentry, P.R.; Plumley, H.C.; Weaver, C.D.; Conn, P.J.; Lindsley, C.W. Discovery of the first highly M5-preferring muscarinic acetylcholine receptor ligand, an M5 positive allosteric modulator derived from a series of 5-trifluoromethoxy N-benzyl isatins. J. Med. Chem. 2009, 52, 3445–3448. [Google Scholar] [CrossRef] [Green Version]
- Kuduk, S.D.; Chang, R.K.; Di Marco, C.N.; Pitts, D.R.; Greshock, T.J.; Ma, L.; Wittmann, M.; Seager, M.A.; Koeplinger, K.A.; Thompson, C.D.; et al. Discovery of a selective allosteric M1 receptor modulator with suitable development properties based on a quinolizidinone carboxylic acid scaffold. J. Med. Chem. 2011, 54, 4773–4780. [Google Scholar] [CrossRef]
- Davie, B.J.; Valant, C.; White, J.M.; Sexton, P.M.; Capuano, B.; Christopoulos, A.; Scammells, P.J. Synthesis and pharmacological evaluation of analogues of benzyl quinolone carboxylic acid (BQCA) designed to bind irreversibly to an allosteric site of the M ₁ muscarinic acetylcholine receptor. J. Med. Chem. 2014, 57, 5405–5418. [Google Scholar] [CrossRef] [PubMed]
- Pancani, T.; Foster, D.J.; Moehle, M.S.; Bichell, T.J.; Bradley, E.; Bridges, T.M.; Klar, R.; Poslusney, M.; Rook, J.M.; Daniels, J.S.; et al. Allosteric activation of M4 muscarinic receptors improve behavioral and physiological alterations in early symptomatic YAC128 mice. Proc. Natl. Acad. Sci. USA 2015, 112, 14078–14083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dallagnol, J.C.C.; Khajehali, E.; Van Der Westhuizen, E.T.; Jörg, M.; Valant, C.; Gonçalves, A.G.; Capuano, B.; Christopoulos, A.; Scammells, P.J. Synthesis and Pharmacological Evaluation of Heterocyclic Carboxamides: Positive Allosteric Modulators of the M1Muscarinic Acetylcholine Receptor with Weak Agonist Activity and Diverse Modulatory Profiles. J. Med. Chem. 2018, 61, 2875–2894. [Google Scholar] [CrossRef] [PubMed]
- Mistry, S.N.; Jörg, M.; Lim, H.; Vinh, N.B.; Sexton, P.M.; Capuano, B.; Christopoulos, A.; Lane, J.R.; Scammells, P.J. 4-Phenylpyridin-2-one Derivatives: A Novel Class of Positive Allosteric Modulator of the M1 Muscarinic Acetylcholine Receptor. J. Med. Chem. 2016, 59, 388–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, Y.; Goldfeld, D.A.; Von Moo, E.; Sexton, P.M.; Christopoulos, A.; McCammon, J.A.; Valant, C. Accelerated structure-based design of chemically diverse allosteric modulators of a muscarinic G protein-coupled receptor. Proc. Natl. Acad. Sci. USA 2016, 113, E5675–E5684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korczynska, M.; Clark, M.J.; Valant, C.; Xu, J.; Von Moo, E.; Albold, S.; Weiss, D.R.; Torosyan, H.; Huang, W.; Kruse, A.C.; et al. Structure-based discovery of selective positive allosteric modulators of antagonists for the M2 muscarinic acetylcholine receptor. Proc. Natl. Acad. Sci. USA 2018, 115, E2419–E2428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldberg, J.A.; Ding, J.B.; Surmeier, D.J. Muscarinic modulation of striatal function and circuitry. Handb. Exp. Pharmacol. 2012, 208, 223–241. [Google Scholar]
- Choy, K.H.C.; Shackleford, D.M.; Malone, D.T.; Mistry, S.N.; Patil, R.T.; Scammells, P.J.; Langmead, C.J.; Pantelis, C.; Sexton, P.M.; Lane, J.R.; et al. Positive Allosteric Modulation of the Muscarinic M1 Receptor Improves Efficacy of Antipsychotics in Mouse Glutamatergic Deficit Models of Behavior. J. Pharmacol. Exp. Ther. 2016, 359, 354–365. [Google Scholar] [CrossRef] [Green Version]
- Nickols, H.H.; Conn, J.P. Development of allosteric modulators of GPCRs for treatment of CNS disorders. Neurobiol. Dis. 2014, 61, 55–71. [Google Scholar] [CrossRef] [Green Version]
- Hopper, S.; Pavey, G.M.; Gogos, A.; Dean, B. Widespread Changes in Positive Allosteric Modulation of the Muscarinic M1 Receptor in Some Participants With Schizophrenia. Int. J. Neuropsychopharmacol. 2019, 22, 640–650. [Google Scholar] [CrossRef] [PubMed]
- Tzavara, E.T.; Bymaster, F.P.; Davis, R.J.; Wade, M.R.; Perry, K.W.; Wess, J.; McKinzie, D.L.; Felder, C.; Nomikos, G.G. M4 muscarinic receptors regulate the dynamics of cholinergic and dopaminergic neurotransmission: relevance to the pathophysiology and treatment of related CNS pathologies. FASEB J. 2004, 18, 1410–1412. [Google Scholar] [CrossRef] [PubMed]
- Brady, A.E.; Jones, C.K.; Bridges, T.M.; Kennedy, J.P.; Thompson, A.D.; Heiman, J.U.; Breininger, M.L.; Gentry, P.R.; Yin, H.; Jadhav, S.B.; et al. Centrally active allosteric potentiators of the M4 muscarinic acetylcholine receptor reverse amphetamine-induced hyperlocomotor activity in rats. J. Pharmacol. Exp. Ther. 2008, 327, 941–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeomans, J.S. Muscarinic receptors in brain stem and mesopontine cholinergic arousal functions. Handb. Exp. Pharmacol. 2012, 208, 243–259. [Google Scholar]
- Gentry, P.R.; Kokubo, M.; Bridges, T.M.; Kett, N.R.; Harp, J.M.; Cho, H.P.; Smith, E.; Chase, P.; Hodder, P.S.; Niswender, C.M.; et al. Discovery of the first M5-selective and CNS penetrant negative allosteric modulator (NAM) of a muscarinic acetylcholine receptor: (S)-9b-(4-chlorophenyl)-1-(3,4-difluorobenzoyl)-2,3-dihydro-1H-imidazo [2,1-a]isoindol-5(9bH)-one (ML375). J. Med. Chem. 2013, 56, 9351–9355. [Google Scholar] [CrossRef] [Green Version]
- Gunter, B.W.; Gould, R.W.; Bubser, M.; McGowan, K.M.; Lindsley, C.W.; Jones, C.K. Selective inhibition of M5 muscarinic acetylcholine receptors attenuates cocaine self-administration in rats. Addict. Biol. 2018, 23, 1106–1116. [Google Scholar] [CrossRef]
- Gilon, P.; Henquin, J.C. Mechanisms and physiological significance of the cholinergic control of pancreatic beta-cell function. Endocr. Rev. 2001, 22, 565–604. [Google Scholar]
- Duttaroy, A.; Zimliki, C.L.; Gautam, D.; Cui, Y.; Mears, D.; Wess, J. Muscarinic stimulation of pancreatic insulin and glucagon release is abolished in M3 muscarinic acetylcholine receptor-deficient mice. Diabetes 2004, 53, 1714–1720. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Rossi, M.; Cohen, A.; Pham, J.; Zheng, H.; Dattaroy, D.; Mukaibo, T.; Melvin, J.E.; Langel, J.L.; Hattar, S.; et al. Allosteric modulation of β-cell M3 muscarinic acetylcholine receptors greatly improves glucose homeostasis in lean and obese mice. Proc. Natl. Acad. Sci. USA 2019, 116, 18684–18690. [Google Scholar] [CrossRef] [Green Version]










© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakubik, J.; El-Fakahany, E.E. Current Advances in Allosteric Modulation of Muscarinic Receptors. Biomolecules 2020, 10, 325. https://doi.org/10.3390/biom10020325
Jakubik J, El-Fakahany EE. Current Advances in Allosteric Modulation of Muscarinic Receptors. Biomolecules. 2020; 10(2):325. https://doi.org/10.3390/biom10020325
Chicago/Turabian StyleJakubik, Jan, and Esam E. El-Fakahany. 2020. "Current Advances in Allosteric Modulation of Muscarinic Receptors" Biomolecules 10, no. 2: 325. https://doi.org/10.3390/biom10020325