DNA Repair Biosensor-Identified DNA Damage Activities of Endophyte Extracts from Garcinia cowa

 ,
, 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Culture Conditions
2.2. Construction of the Fluorescent Biosensor Clones
2.3. Proliferation Determination
2.4. Focus Formation by Immunofluorescence
2.5. Plant Extraction
2.6. Isolation of Endophytic Fungi from Garcinia cowa
2.7. Fermentation Condition and Metabolite Extraction
2.8. Determination of Viability with DNA Repair-Deficient Cell Line Panels
2.9. Statistical Analysis
3. Results
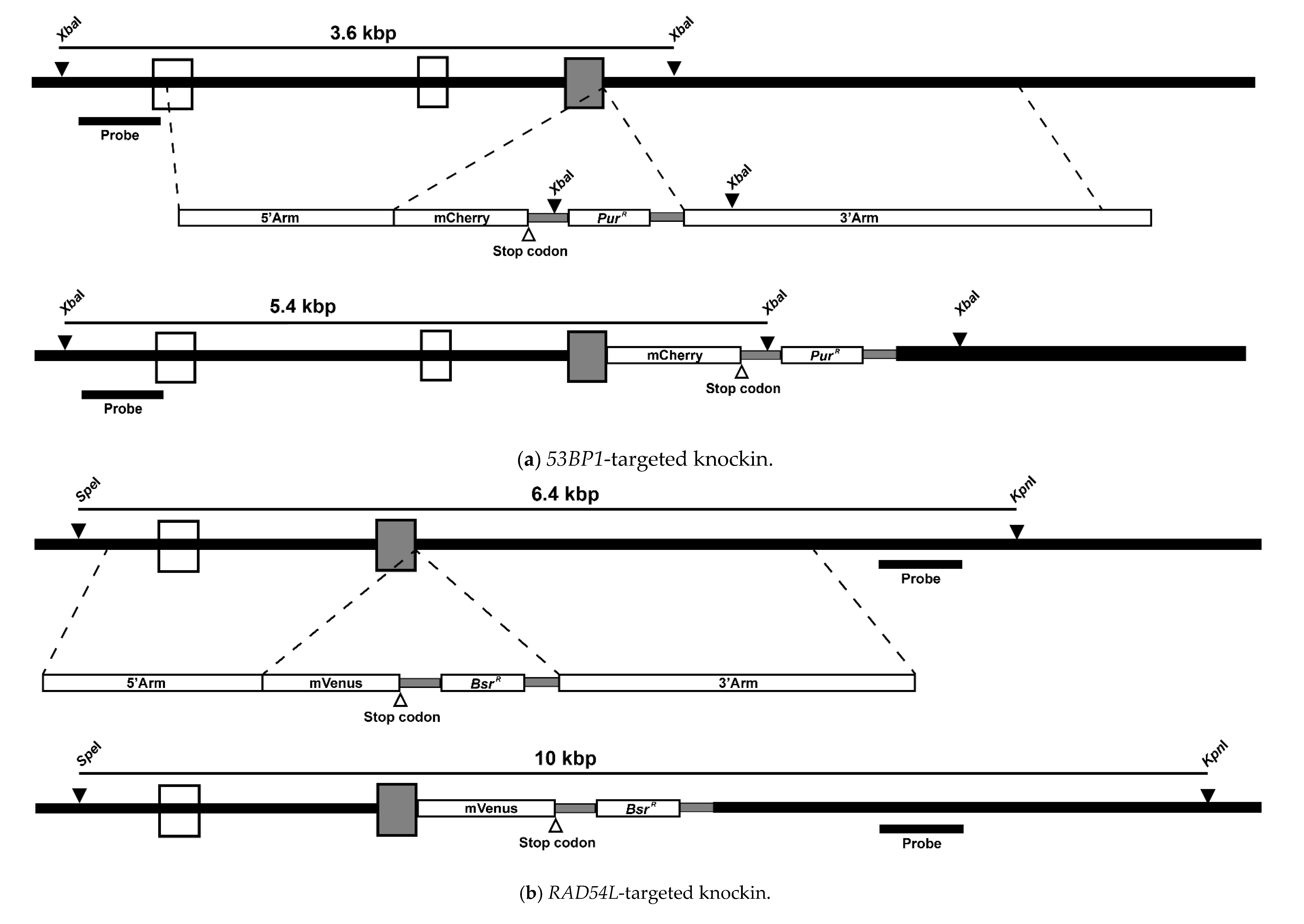
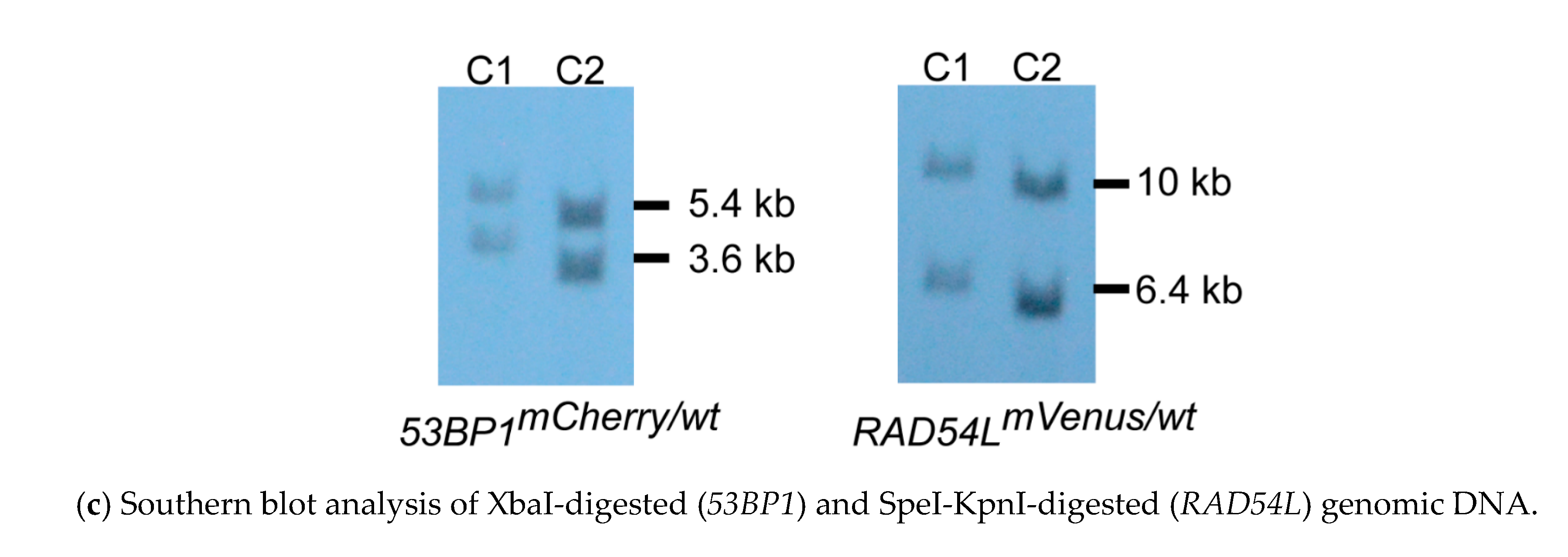
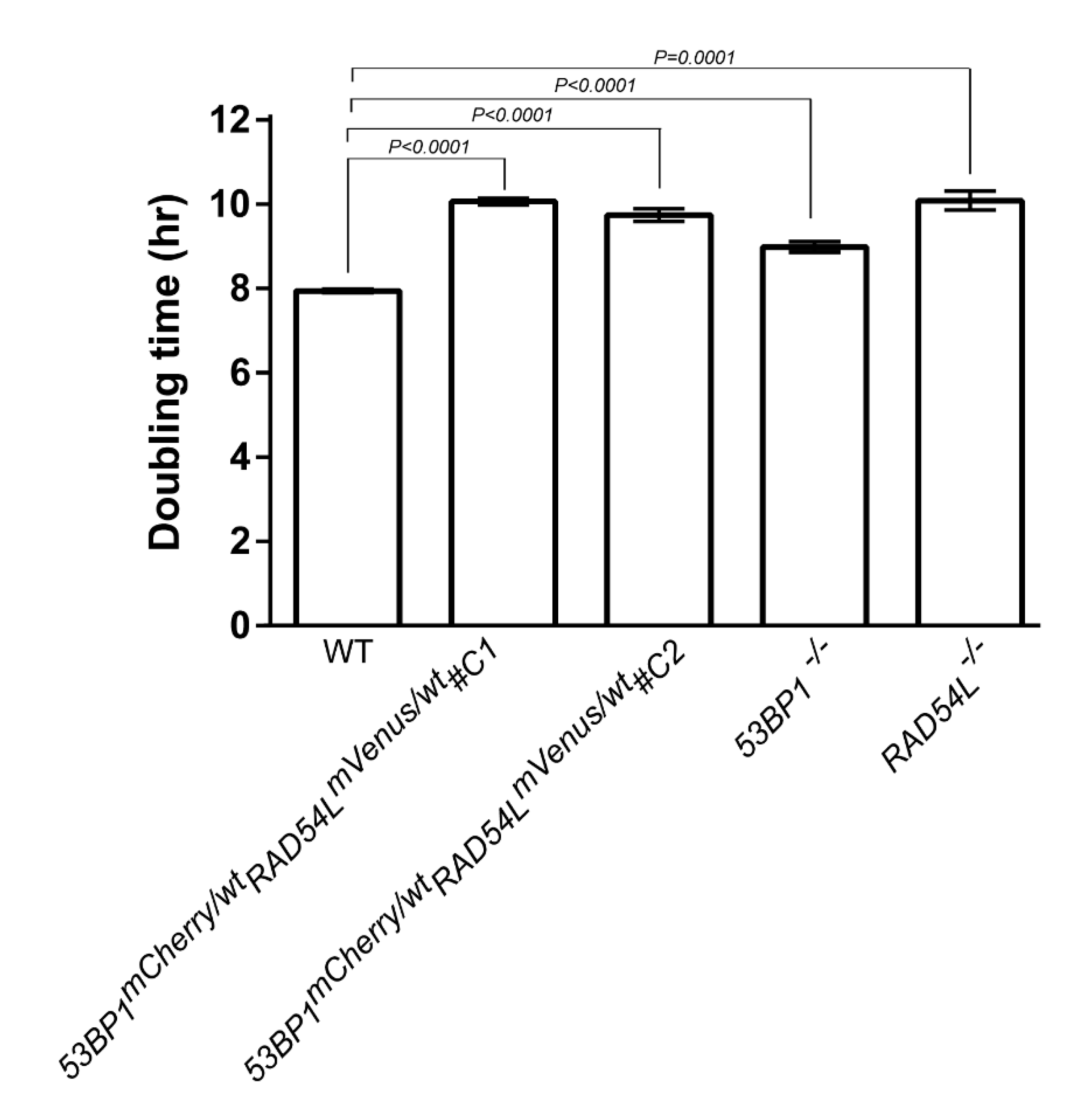
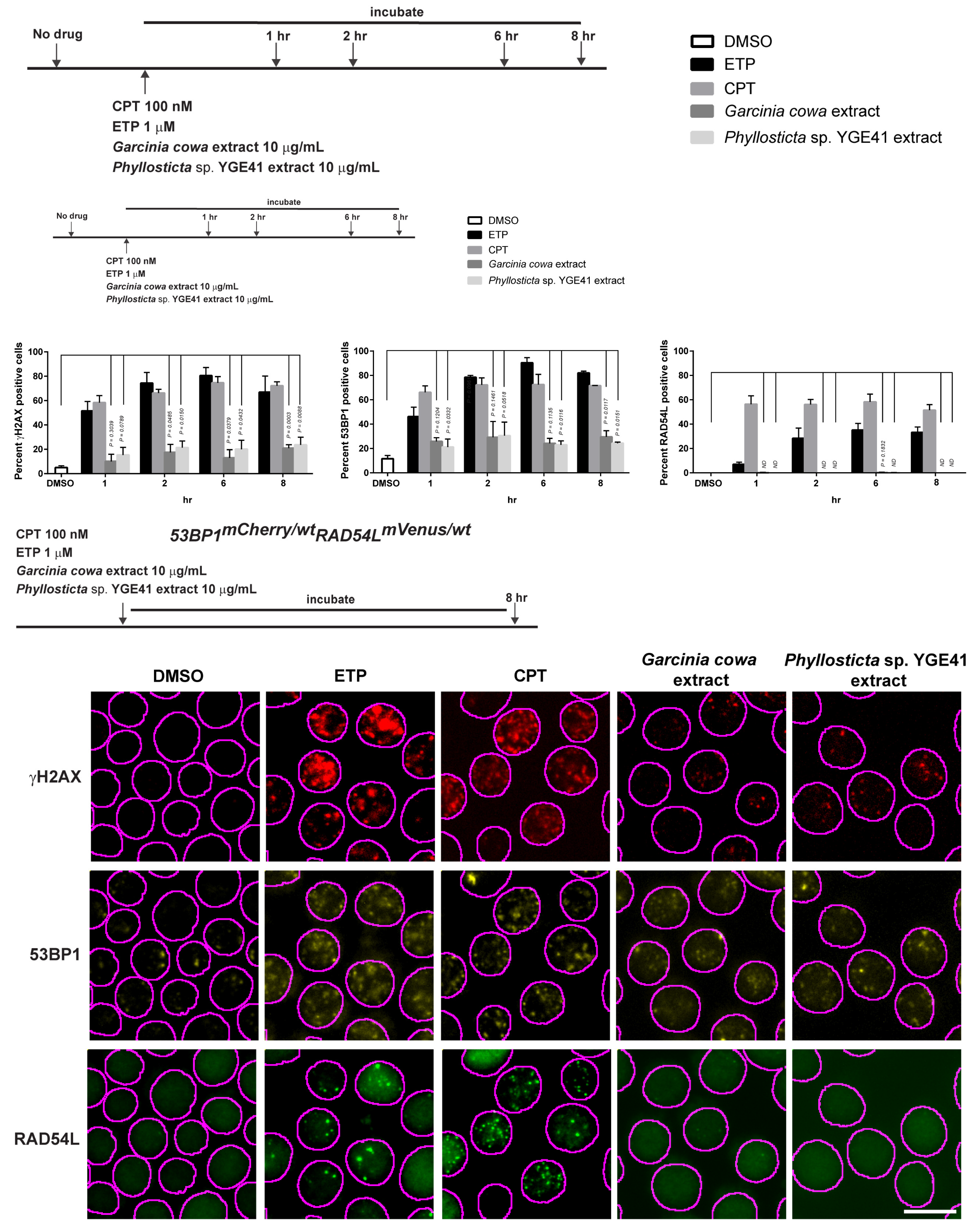
3.1. Fluorescent Biosensor Construction and Proliferative Properties of Biosensor Clones
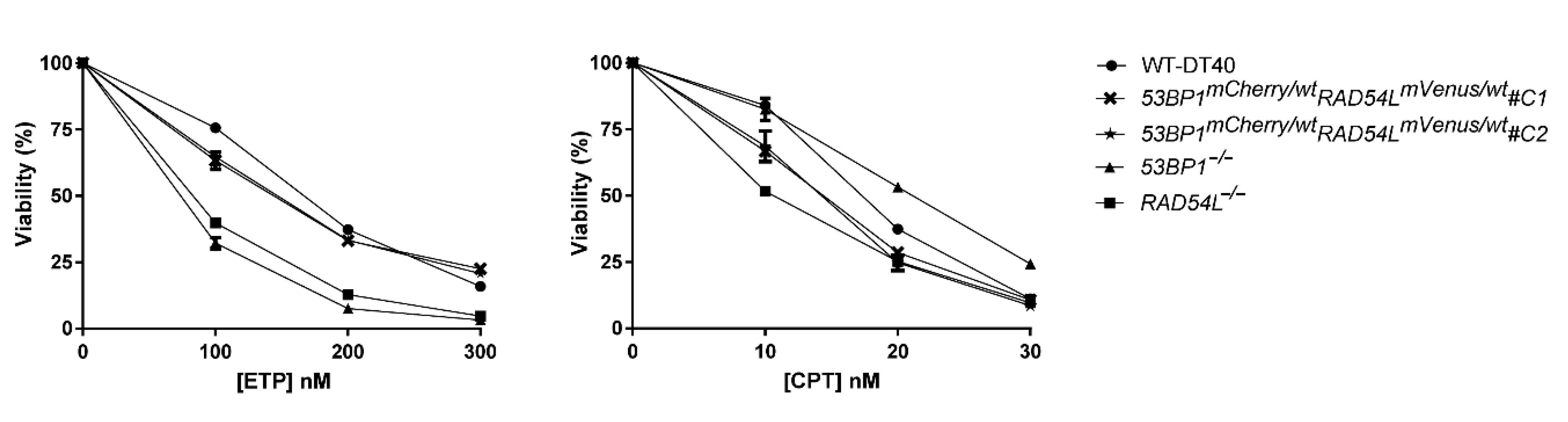
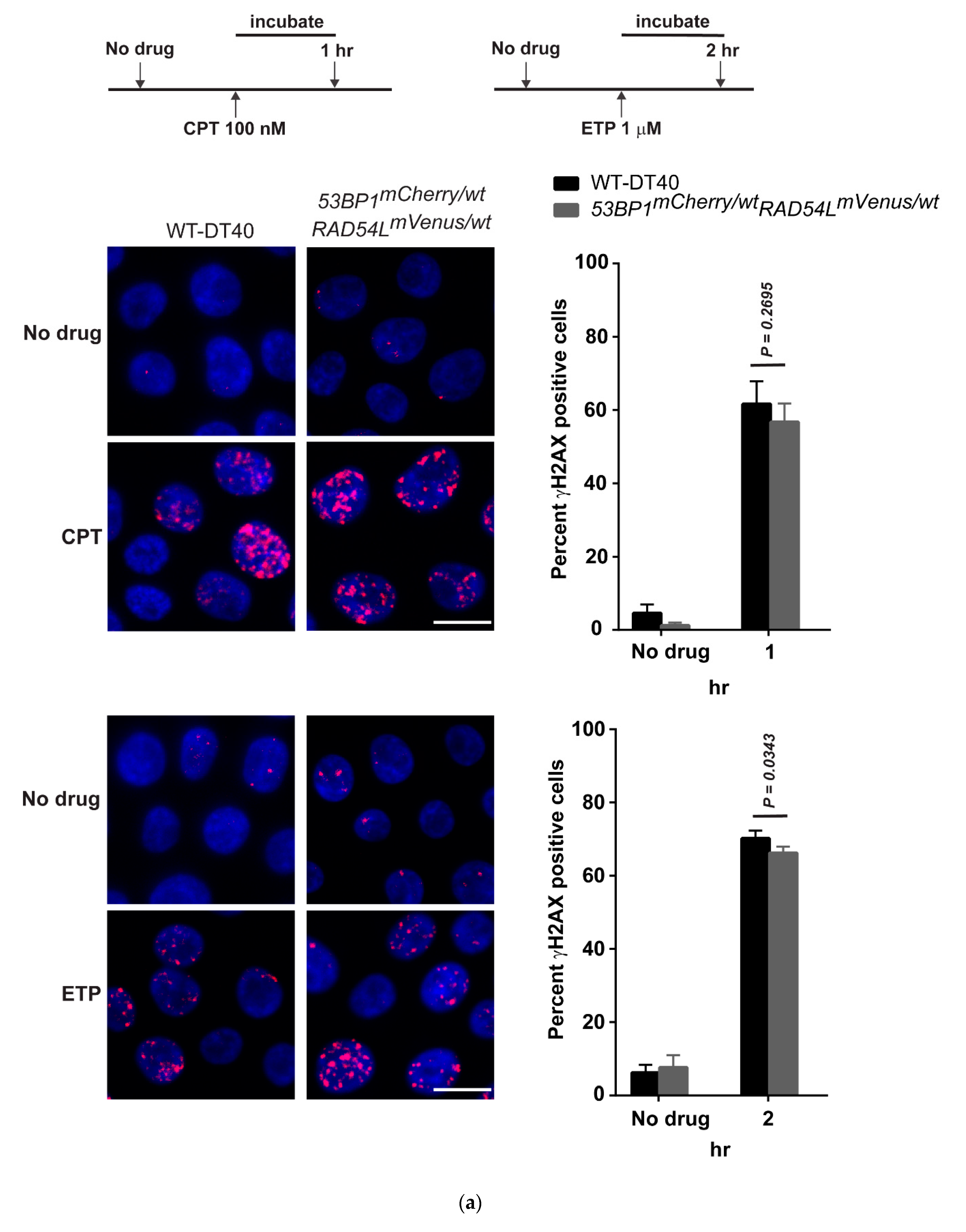
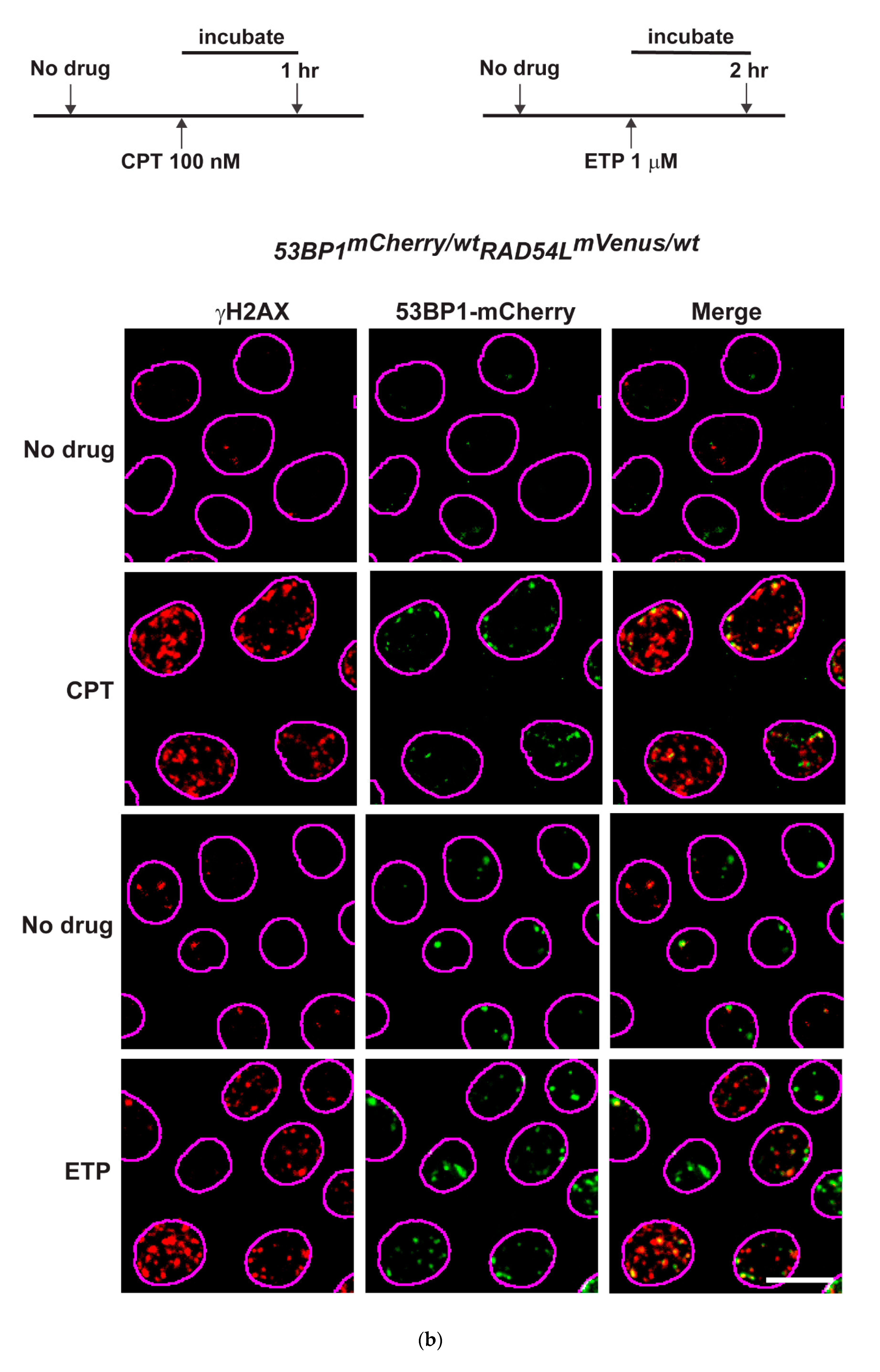
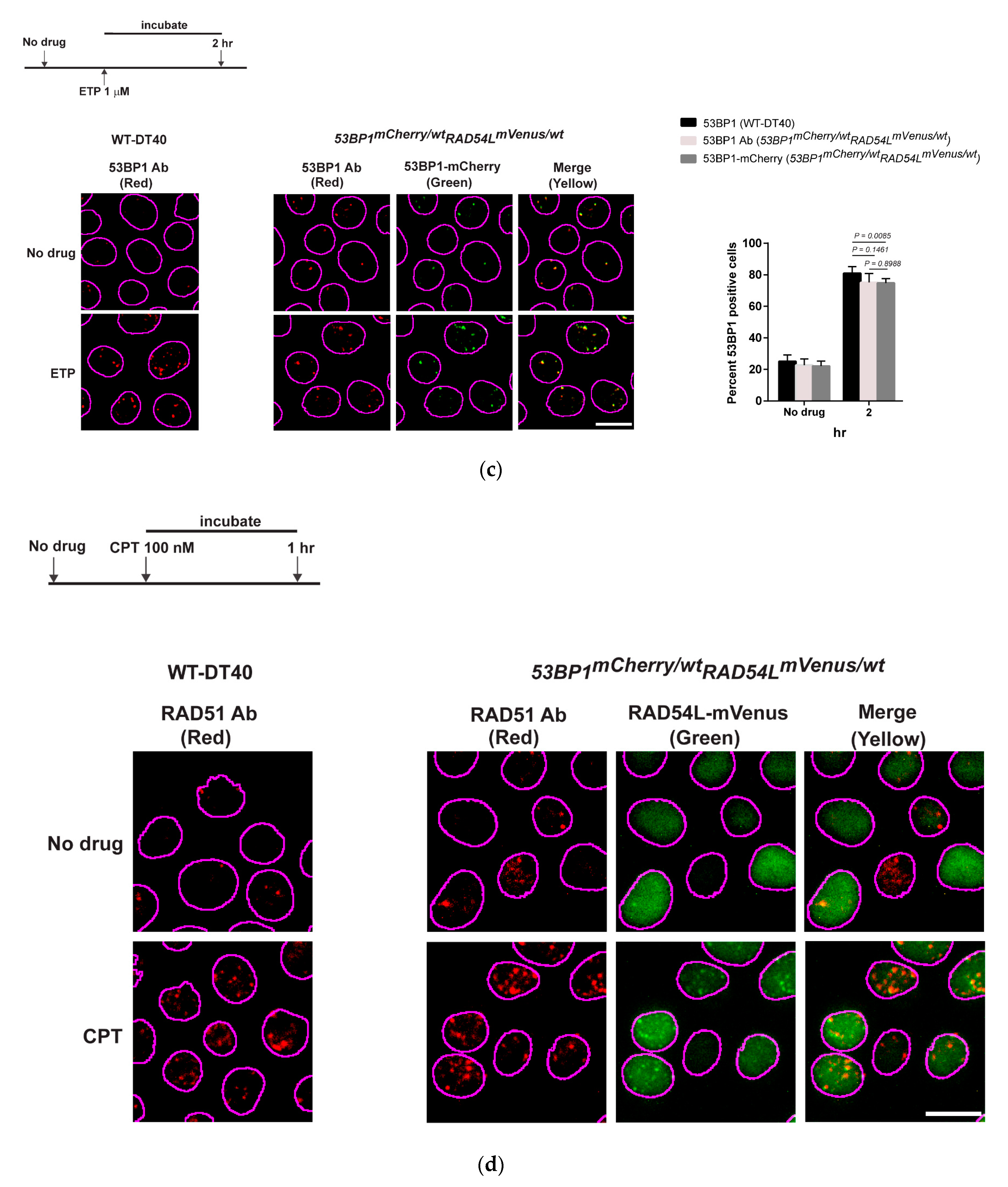
3.2. DNA Repair Phenotypic Validation of Biosensor Clones
3.3. Expression of Fluorescent mCherry 53BP1 and mVenus-RAD54L Proteins
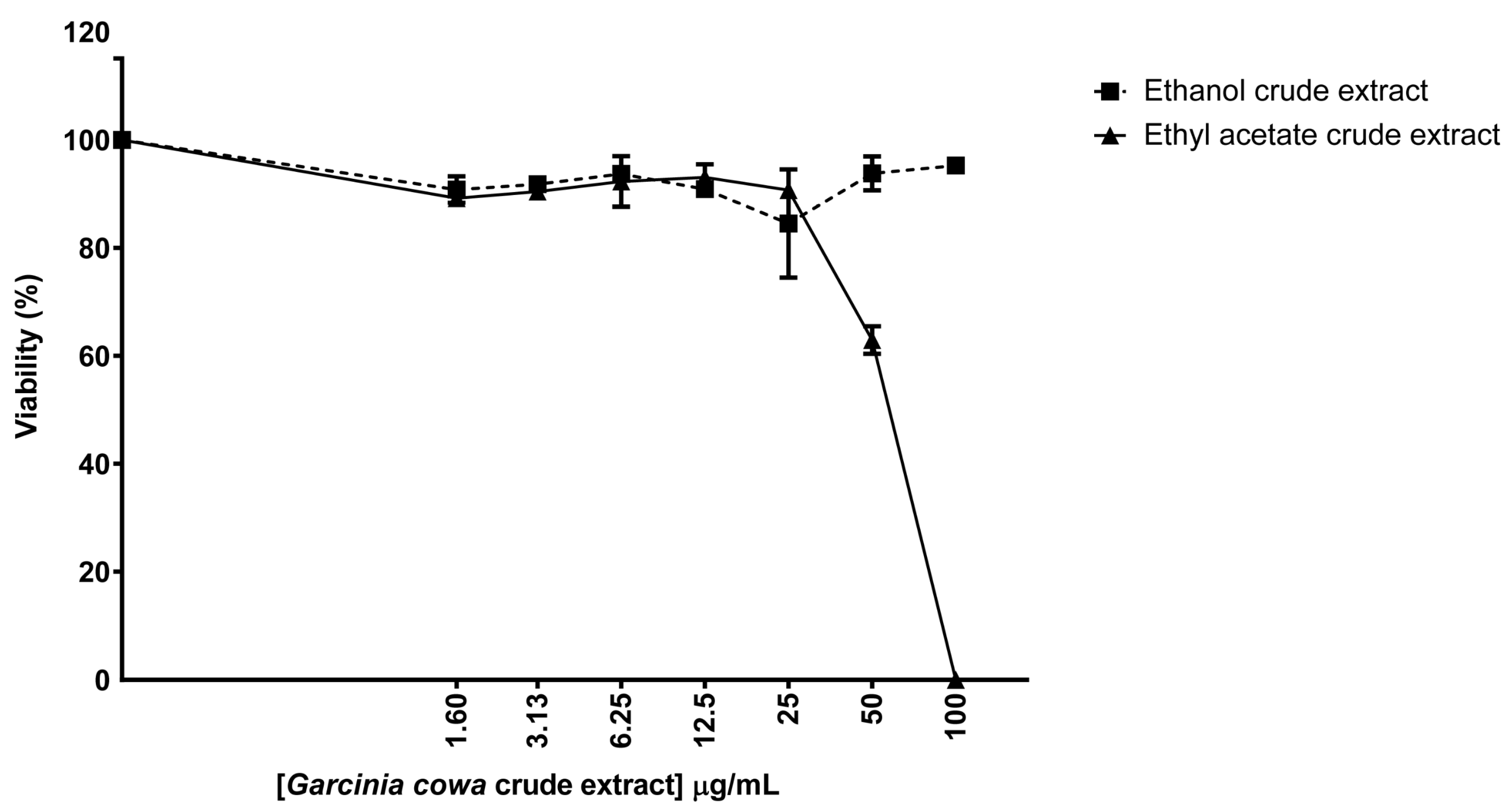
3.4. Biological Properties of Crude Extracts from Garcinia cowa
3.5. Identification of Fungal Endophyte from Garcinia cowa
3.6. Biological Properties of Crude Extracts from Phyllosticta sp. YGE41
3.7. Garcinia cowa Extract-Induced DNA DSBs Damage through 53BP1
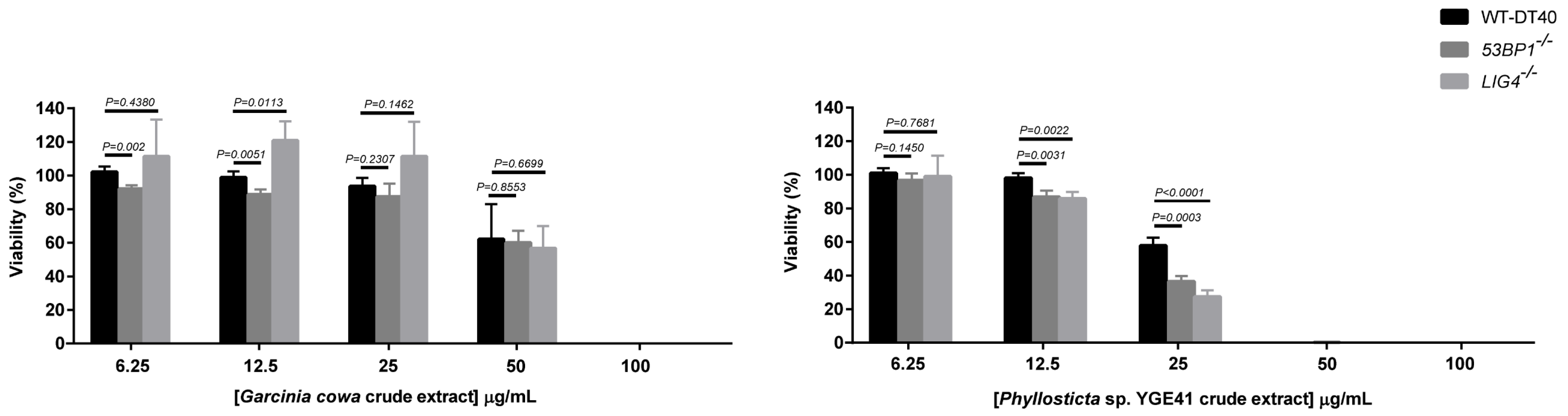
3.8. Viability of Crude Extract with NHEJ-Deficient Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schirrmacher, V. From chemotherapy to biological therapy: A review of novel concepts to reduce the side effects of systemic cancer treatment (Review). Int. J. Oncol. 2019, 54, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Remon, J.; Dienstmann, R. Precision oncology: Separating the wheat from the chaff. ESMO Open 2018, 3, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helleday, T.; Petermann, E.; Lundin, C.; Hodgson, B.; Sharma, R.A. DNA repair pathways as targets for cancer therapy. Nat. Rev. Cancer 2008, 8, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Nickoloff, J.A.; Jones, D.; Lee, S.H.; Williamson, E.A.; Hromas, R. Drugging the Cancers Addicted to DNA Repair. J. Natl. Cancer Inst. 2017, 109, 1–13. [Google Scholar] [CrossRef]
- Lee, J.M.; Ledermann, J.A.; Kohn, E.C. PARP Inhibitors for BRCA1/2 mutation-associated and BRCA-like malignancies. Ann. Oncol. 2014, 25, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Hosoya, N.; Miyagawa, K. Targeting DNA damage response in cancer therapy. Cancer Sci. 2014, 105, 370–388. [Google Scholar] [CrossRef] [Green Version]
- O’Kane, G.M.; Connor, A.A.; Gallinger, S. Characterization, Detection, and Treatment Approaches for Homologous Recombination Deficiency in Cancer. Trends Mol. Med. 2017, 23, 1121–1137. [Google Scholar] [CrossRef]
- Murai, J.; Pommier, Y. PARP Trapping Beyond Homologous Recombination and Platinum Sensitivity in Cancers. Annu. Rev. Cancer Biol. 2019, 3, 131–150. [Google Scholar] [CrossRef]
- Prasad, V.; De Jesus, K.; Mailankody, S. The high price of anticancer drugs: Origins, implications, barriers, solutions. Nat. Rev. Clin. Oncol. 2017, 14, 381–390. [Google Scholar] [CrossRef]
- Schleicher, S.M.; Bach, P.B.; Matsoukas, K.; Korenstein, D. Medication overuse in oncology: Current trends and future implications for patients and society. Lancet Oncol. 2018, 19, 200–208. [Google Scholar] [CrossRef]
- Kinghorn, A.D.; EJ, D.E.B.; Lucas, D.M.; Rakotondraibe, H.L.; Orjala, J.; Soejarto, D.D.; Oberlies, N.H.; Pearce, C.J.; Wani, M.C.; Stockwell, B.R.; et al. Discovery of Anticancer Agents of Diverse Natural Origin. Anticancer Res. 2016, 36, 5623–5637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strobel, G. The Emergence of Endophytic Microbes and Their Biological Promise. J. Fungi 2018, 4, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, D.J. Are Microbial Endophytes the ‘Actual’ Producers of Bioactive Antitumor Agents? Trends Cancer 2018, 4, 662–670. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Chen, L.; Xin, H.L.; Zheng, C.J.; Rahman, K.; Han, T.; Qin, L.P. A Friendly Relationship between Endophytic Fungi and Medicinal Plants: A Systematic Review. Front. Microbiol. 2016, 7, 906–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strobel, G.; Daisy, B.; Castillo, U.; Harper, J. Natural products from endophytic microorganisms. J. Nat. Prod. 2004, 67, 257–268. [Google Scholar] [CrossRef]
- Strobel, G.; Daisy, B. Bioprospecting for microbial endophytes and their natural products. Microbiol. Mol. Biol. Rev. 2003, 67, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Uzma, F.; Mohan, C.D.; Hashem, A.; Konappa, N.M.; Rangappa, S.; Kamath, P.V.; Singh, B.P.; Mudili, V.; Gupta, V.K.; Siddaiah, C.N.; et al. Endophytic Fungi-Alternative Sources of Cytotoxic Compounds: A Review. Front. Pharmacol. 2018, 9, 309–346. [Google Scholar] [CrossRef]
- Chakravarty, K.; Gaur, S. Fungal Endophytes as Novel Sources of Anticancer Compounds. In Anticancer Plants: Natural Products and Biotechnological Implements: Volume 2; Akhtar, M.S., Swamy, M.K., Eds.; Springer: Singapore, 2018; pp. 1–18. [Google Scholar] [CrossRef]
- Wall, M.E.; Wani, M.C. Camptothecin and taxol: From discovery to clinic. J. Ethnopharmacol. 1996, 51, 239–253; discussion 253-234. [Google Scholar] [CrossRef]
- Miller, K.; Neilan, B.; Sze, D.M. Development of Taxol and other endophyte produced anti-cancer agents. Recent Pat. Anticancer Drug Discov. 2008, 3, 14–19. [Google Scholar]
- Vasundhara, M.; Kumar, A.; Reddy, M.S. Molecular Approaches to Screen Bioactive Compounds from Endophytic Fungi. Front. Microbiol. 2016, 7, 1774–1786. [Google Scholar] [CrossRef] [Green Version]
- Kusari, S.; Zuhlke, S.; Spiteller, M. An endophytic fungus from Camptotheca acuminata that produces camptothecin and analogues. J. Nat. Prod. 2009, 72, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Shan, T.; Mou, Y.; Zhou, L. Plant-derived bioactive compounds produced by endophytic fungi. Mini Rev. Med. Chem. 2011, 11, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhang, X.; Bussmann, R.W.; Hart, R.H.; Li, P.; Bai, Y.; Long, C. Garcinia in Southern China: Ethnobotany, Management, and Niche Modeling. Econ. Bot. 2016, 70, 416–430. [Google Scholar] [CrossRef]
- Gogoi, B.; Das, R.P.; Sarma, R.N.; Barua, U. Morpholocical characters and floral biology of Garcinia species of Assam. Ecol. Environ. Conserv. 2016, 22, 1929–1933. [Google Scholar]
- Lim, T.K. Garcinia cowa. In Edible Medicinal And Non-Medicinal Plants: Volume 2, Fruits; Springer: Dordrecht, The Netherlands, 2012; pp. 29–34. [Google Scholar] [CrossRef]
- Yapwattanaphun, C.; Subhadrabandhu, S.; Sugiura, A.; Yonemori, K.; Utsunomiya, N. Utilization of some Garcinia species in Thailand. Acta Horticult. 2002, 575, 563–570. [Google Scholar] [CrossRef]
- Ezhilarasi, P.N.; Indrani, D.; Jena, B.S.; Anandharamakrishnan, C. Freeze drying technique for microencapsulation of Garcinia fruit extract and its effect on bread quality. J. Food Eng. 2013, 117, 513–520. [Google Scholar] [CrossRef]
- Ritthiwigrom, T.; Laphookhieo, S.; Pyne, S.G. Chemical constituents and biological activities of Garcinia cowa Roxb. Maejo Int. J. Sci. Technol. 2013, 7, 212–231. [Google Scholar]
- Likhitwitayawuid, K.; Phadungcharoen, T.; Krungkrai, J. Antimalarial xanthones from Garcinia cowa. Planta Med. 1998, 64, 70–72. [Google Scholar] [CrossRef]
- Lipipun, V.; Kurokawa, M.; Suttisri, R.; Taweechotipatr, P.; Pramyothin, P.; Hattori, M.; Shiraki, K. Efficacy of Thai medicinal plant extracts against herpes simplex virus type 1 infection in vitro and in vivo. Antivir. Res. 2003, 60, 175–180. [Google Scholar] [CrossRef]
- Joseph, G.S.; Jayaprakasha, G.K.; Selvi, A.T.; Jena, B.S.; Sakariah, K.K. Antiaflatoxigenic and antioxidant activities of Garcinia extracts. Int. J. Food Microbiol. 2005, 101, 153–160. [Google Scholar] [CrossRef]
- Shen, J.; Tian, Z.; Yang, J.S. The constituents from the stems of Garcinia cowa Roxb. and their cytotoxic activities. Pharmazie 2007, 62, 549–551. [Google Scholar] [CrossRef] [PubMed]
- Negi, P.S.; Jayaprakasha, G.K.; Jena, B.S. Antibacterial activity of the extracts from the fruit rinds of Garcinia cowa and Garcinia pedunculata against food borne pathogens and spoilage bacteria. LWT Food Sci. Technol. 2008, 41, 1857–1861. [Google Scholar] [CrossRef]
- Jabit, M.L.; Wahyuni, F.S.; Khalid, R.; Israf, D.A.; Shaari, K.; Lajis, N.H.; Stanslas, J. Cytotoxic and nitric oxide inhibitory activities of methanol extracts of garcinia species. Pharm. Biol. 2009, 47, 1019–1026. [Google Scholar] [CrossRef]
- Nanasombat, S.; Teckchuen, N. Antimicrobial, antioxidant and anticancer activities of Thai local vegetables. J. Med. Plants Res. 2009, 3, 443–449. [Google Scholar]
- Konstantinopoulos, P.A.; Matulonis, U.A. Targeting DNA Damage Response and Repair as a Therapeutic Strategy for Ovarian Cancer. Hematol. Oncol. Clin. North Am. 2018, 32, 997–1010. [Google Scholar] [CrossRef] [PubMed]
- Sakunpak, A.; Panichayupakaranant, P. Antibacterial activity of Thai edible plants against gastrointestinal pathogenic bacteria and isolation of a new broad spectrum antibacterial polyisoprenylated benzophenone, chamuangone. Food Chem. 2012, 130, 826–831. [Google Scholar] [CrossRef]
- Kan, W.L.T.; Yin, C.; Xu, H.X.; Xu, G.; To, K.K.W.; Cho, C.H.; Rudd, J.A.; Lin, G. Antitumor effects of novel compound, guttiferone K, on colon cancer by p21Waf1/Cip1-mediated G0/G1 cell cycle arrest and apoptosis. Int. J. Cancer 2013, 132, 707–716. [Google Scholar] [CrossRef]
- Tayana, N.; Suteerapataranon, S.; Deachathai, S. Phytochemistry and bioactive compounds from Garcinia cowa roxb. Asia Pac. J. Sci. Technol. 2017, 22, 1–8. [Google Scholar] [CrossRef]
- Chowchaikong, N.; Nilwarangkoon, S.; Laphookhieo, S.; Tanunyutthawongse, C.; Watanapokasin, R. p38 inhibitor inhibits the apoptosis of cowanin-treated human colorectal adenocarcinoma cells. Int. J. Oncol. 2018, 52, 2031–2040. [Google Scholar] [CrossRef] [Green Version]
- Sriyatep, T.; Siridechakorn, I.; Maneerat, W.; Pansanit, A.; Ritthiwigrom, T.; Andersen, R.J.; Laphookhieo, S. Bioactive prenylated xanthones from the young fruits and flowers of Garcinia cowa. J. Nat. Prod. 2015, 78, 265–271. [Google Scholar] [CrossRef]
- Sae-Lim, P.; Yuenyongsawad, S.; Panichayupakaranant, P. Chamuangone-enriched Garcinia cowa leaf extract with rice bran oil: Extraction and cytotoxic activity against cancer cells. Pharmacogn. Mag. 2019, 15, 183–188. [Google Scholar] [CrossRef]
- Xu, G.; Kan, W.L.T.; Zhou, Y.; Song, J.Z.; Han, Q.B.; Qiao, C.F.; Cho, C.H.; Rudd, J.A.; Lin, G.; Xu, H.X. Cytotoxic acylphloroglucinol derivatives from the twigs of garcinia cowa. J. Natural. Prod. 2010, 73, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Husni, E.; Nahari, F.; Wirasti, Y.; Wahyuni, F.S. Dachriyanus. Cytotoxicity study of ethanol extract of the stem bark of asam kandis (Garcinia cowa Roxb.) on T47D breast cancer cell line. Asian Pac. J. Trop. Biomed 2015, 5, 249–252. [Google Scholar] [CrossRef] [Green Version]
- Mao, Z.; Bozzella, M.; Seluanov, A.; Gorbunova, V. DNA repair by nonhomologous end joining and homologous recombination during cell cycle in human cells. Cell Cycle 2008, 7, 2902–2906. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Bozzella, M.; Seluanov, A.; Gorbunova, V. Comparison of nonhomologous end joining and homologous recombination in human cells. DNA Repair 2008, 7, 1765–1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chien, J.C.; Tabet, E.; Pinkham, K.; da Hora, C.C.; Chang, J.C.; Lin, S.; Badr, C.E.; Lai, C.P. A multiplexed bioluminescent reporter for sensitive and non-invasive tracking of DNA double strand break repair dynamics in vitro and in vivo. Nucleic Acids Res. 2020, 48, 1–49. [Google Scholar] [CrossRef] [PubMed]
- Dhar, P.K.; Sonoda, E.; Fujimori, A.; Yamashita, Y.M.; Takeda, S. DNA Repair Studies: Experimental Evidence in Support of Chicken DT40 Cell Line as a Unique Model. J. Environ. Pathol. Toxicol. Oncol. 2001, 20, 11. [Google Scholar] [CrossRef]
- Yamazoe, M.; Sonoda, E.; Hochegger, H.; Takeda, S. Reverse genetic studies of the DNA damage response in the chicken B lymphocyte line DT40. DNA Repair 2004, 3, 1175–1185. [Google Scholar] [CrossRef]
- Sonoda, E.; Morrison, C.; Yamashita, Y.M.; Takata, M.; Takeda, S. Reverse genetic studies of homologous DNA recombination using the chicken B-lymphocyte line, DT40. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2001, 356, 111–117. [Google Scholar] [CrossRef]
- Al Abo, M.; Dejsuphong, D.; Hirota, K.; Yonetani, Y.; Yamazoe, M.; Kurumizaka, H.; Takeda, S. Compensatory functions and interdependency of the DNA-binding domain of BRCA2 with the BRCA1-PALB2-BRCA2 complex. Cancer Res. 2014, 74, 797–807. [Google Scholar] [CrossRef] [Green Version]
- Murai, J.; Marchand, C.; Shahane, S.A.; Sun, H.; Huang, R.; Zhang, Y.; Chergui, A.; Ji, J.; Doroshow, J.H.; Jadhav, A.; et al. Identification of novel PARP inhibitors using a cell-based TDP1 inhibitory assay in a quantitative high-throughput screening platform. DNA Repair 2014, 21, 177–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishihara, K.; Huang, R.; Zhao, J.; Shahane, S.A.; Witt, K.L.; Smith-Roe, S.L.; Tice, R.R.; Takeda, S.; Xia, M. Identification of genotoxic compounds using isogenic DNA repair deficient DT40 cell lines on a quantitative high throughput screening platform. Mutagenesis 2016, 31, 69–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, K.; Kogame, T.; Choi, K.; Wang, X.; Lee, J.; Taniguchi, Y.; Takeda, S. A novel approach using DNA-repair-deficient chicken DT40 cell lines for screening and characterizing the genotoxicity of environmental contaminants. Environ. Health Perspect. 2009, 117, 1737–1744. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.N.; Hirota, K.; Kono, K.; Takeda, S.; Sakamuru, S.; Xia, M.; Huang, R.; Austin, C.P.; Witt, K.L.; Tice, R.R. Characterization of environmental chemicals with potential for DNA damage using isogenic DNA repair-deficient chicken DT40 cell lines. Environ. Mol. Mutagen. 2011, 52, 547–561. [Google Scholar] [CrossRef]
- Bezzubova, O.; Silbergleit, A.; Yamaguchi-Iwai, Y.; Takeda, S.; Buerstedde, J.M. Reduced X-ray resistance and homologous recombination frequencies in a RAD54-/-mutant of the chicken DT40 cell line. Cell 1997, 89, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Iwabuchi, K.; Hashimoto, M.; Matsui, T.; Kurihara, T.; Shimizu, H.; Adachi, N.; Ishiai, M.; Yamamoto, K.; Tauchi, H.; Takata, M.; et al. 53BP1 contributes to survival of cells irradiated with X-ray during G1 without Ku70 or Artemis. Genes Cells 2006, 11, 935–948. [Google Scholar] [CrossRef]
- Adachi, N.; Ishino, T.; Ishii, Y.; Takeda, S.; Koyama, H. DNA ligase IV-deficient cells are more resistant to ionizing radiation in the absence of Ku70: Implications for DNA double-strand break repair. Proc. Natl. Acad. Sci. USA 2001, 98, 12109–12113. [Google Scholar] [CrossRef] [Green Version]
- Ridpath, J.R.; Takeda, S.; Swenberg, J.A.; Nakamura, J. Convenient, multi-well plate-based DNA damage response analysis using DT40 mutants is applicable to a high-throughput genotoxicity assay with characterization of modes of action. Environ. Mol. Mutagenesis 2011, 52, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Arakawa, H.; Lodygin, D.; Buerstedde, J.M. Mutant loxP vectors for selectable marker recycle and conditional knock-outs. BMC Biotechnol. 2001, 1, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Arakawa, H.; Buerstedde, J.-M. DT40 gene disruptions: A how-to for the design and the construction of targeting vectors. In Reviews and Protocols in DT40 Research: Subcellular Biochemistry; Buerstedde, J.-M., Takeda, S., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 1–9. [Google Scholar] [CrossRef]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529–536. [Google Scholar] [CrossRef]
- Wiyakrutta, S.; Sriubolmas, N.; Panphut, W.; Thongon, N.; Danwisetkanjana, K.; Ruangrungsi, N.; Meevootisom, V. Endophytic fungi with anti-microbial, anti-cancer and anti-malarial activities isolated from Thai medicinal plants. World J. Microbiol. Biotechnol. 2004, 20, 265–272. [Google Scholar] [CrossRef]
- Potshangbam, M.; Devi, S.I.; Sahoo, D.; Strobel, G.A. Functional Characterization of Endophytic Fungal Community Associated with Oryza sativa L. and Zea mays L. Front. Microbiol. 2017, 8, 325–340. [Google Scholar] [CrossRef] [PubMed]
- Jumpathong, J.; Nuengchamnong, N.; Masin, K.; Nakaew, N.; Suphrom, N. Thin layer chromatography-bioautography assay for antibacterial compounds from streptomyces sp. TBRC 8912, a newly isolated actinomycin D producer. Chiang Mai J. Sci. 2019, 46, 839–849. [Google Scholar]
- Heyer, W.-D.; Li, X.; Rolfsmeier, M.; Zhang, X.-P. Rad54: The Swiss Army knife of homologous recombination? Nucleic Acids Res. 2006, 34, 4115–4125. [Google Scholar] [CrossRef]
- Maede, Y.; Shimizu, H.; Fukushima, T.; Kogame, T.; Nakamura, T.; Miki, T.; Takeda, S.; Pommier, Y.; Murai, J. Differential and common DNA repair pathways for topoisomerase I- and II-targeted drugs in a genetic DT40 repair cell screen panel. Mol. Cancer Ther. 2014, 13, 214–220. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, M.; de Lange, T. 53BP1: Pro choice in DNA repair. Trends Cell Biol. 2014, 24, 108–117. [Google Scholar] [CrossRef] [Green Version]
- Rogakou, E.P.; Pilch, D.R.; Orr, A.H.; Ivanova, V.S.; Bonner, W.M. DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139. J. Biol. Chem. 1998, 273, 5858–5868. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.H.Y.; Pannunzio, N.R.; Adachi, N.; Lieber, M.R. Non-homologous DNA end joining and alternative pathways to double-strand break repair. Nat. Rev. Mol. Cell Biol. 2017, 18, 495–506. [Google Scholar] [CrossRef]
- Winding, P.; Berchtold, M.W. The chicken B cell line DT40: A novel tool for gene disruption experiments. J. Immunol. Methods 2001, 249, 1–16. [Google Scholar] [CrossRef]
- Oestergaard, V.H.; Pentzold, C.; Pedersen, R.T.; Iosif, S.; Alpi, A.; Bekker-Jensen, S.; Mailand, N.; Lisby, M. RNF8 and RNF168 but not HERC2 are required for DNA damage-induced ubiquitylation in chicken DT40 cells. DNA Repair 2012, 11, 892–905. [Google Scholar] [CrossRef]
- Pedersen, R.T.; Kruse, T.; Nilsson, J.; Oestergaard, V.H.; Lisby, M. TopBP1 is required at mitosis to reduce transmission of DNA damage to G1 daughter cells. J. Cell Biol. 2015, 210, 565–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, S.; van Cappellen, W.A.; Guenole, A.; Eppink, B.; Linsen, S.E.; Meijering, E.; Houtsmuller, A.; Kanaar, R.; Essers, J. ATP-dependent and independent functions of Rad54 in genome maintenance. J. Cell Biol. 2011, 192, 735–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eppink, B.; Tafel, A.A.; Hanada, K.; van Drunen, E.; Hickson, I.D.; Essers, J.; Kanaar, R. The response of mammalian cells to UV-light reveals Rad54-dependent and independent pathways of homologous recombination. DNA Repair 2011, 10, 1095–1105. [Google Scholar] [CrossRef] [PubMed]
- Polo, S.E.; Jackson, S.P. Dynamics of DNA damage response proteins at DNA breaks: A focus on protein modifications. Genes Dev. 2011, 25, 409–433. [Google Scholar] [CrossRef] [Green Version]
- Tan, T.L.; Kanaar, R.; Wyman, C. Rad54, a Jack of all trades in homologous recombination. DNA Repair 2003, 2, 787–794. [Google Scholar] [CrossRef]
- Li, S.J.; Zhang, X.; Wang, X.H.; Zhao, C.Q. Novel natural compounds from endophytic fungi with anticancer activity. Eur. J. Med. Chem. 2018, 156, 316–343. [Google Scholar] [CrossRef]
- Xia, Z.; Zhang, H.; Xu, D.; Lao, Y.; Fu, W.; Tan, H.; Cao, P.; Yang, L.; Xu, H. Xanthones from the leaves of garcinia cowa induce cell cycle arrest, apoptosis, and autophagy in cancer cells. Molecules 2015, 20, 11387–11399. [Google Scholar] [CrossRef] [Green Version]
- Tanvir, R.; Javeed, A.; Bajwa, A.G. Endophyte bioprospecting in South Asian medicinal plants: An attractive resource for biopharmaceuticals. Appl. Microbiol. Biotechnol. 2017, 101, 1831–1844. [Google Scholar] [CrossRef]
- Martin, A.C.; Pawlus, A.D.; Jewett, E.M.; Wyse, D.L.; Angerhofer, C.K.; Hegeman, A.D. Evaluating solvent extraction systems using metabolomics approaches. RSC Adv. 2014, 4, 26325–26334. [Google Scholar] [CrossRef]
- Arai, M.A.; Akamine, R.; Tsuchiya, A.; Yoneyama, T.; Koyano, T.; Kowithayakorn, T.; Ishibashi, M. The Notch inhibitor cowanin accelerates nicastrin degradation. Sci. Rep. 2018, 8, 5376. [Google Scholar] [CrossRef]
- Shiekh, K.A.; Benjakul, S.; Sae-Leaw, T. Effect of Chamuang (Garcinia cowaRoxb.) leaf extract on inhibition of melanosis and quality changes of Pacific white shrimp during refrigerated storage. Food Chem. 2019, 270, 554–561. [Google Scholar] [CrossRef] [PubMed]
- Molitor, D.; Beyer, M. Epidemiology, identification and disease management of grape black rot and potentially useful metabolites of black rot pathogens for industrial applications—A review. Ann. Appl. Biol. 2014, 165, 305–317. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | 5′ to 3′ Direction | Description |
|---|---|---|
| pBlue-5′Arm53BP | CACCGCGGTGGCGGCCGCTCTAGAAAATACTTGCTGTGCCTTG | 5′-Arm 53BP1 |
| mTur-BglII-5′Arm53BP | CCTTGCTCACCATACGAGGGACATAGTCATG | 5′-Arm 53BP1 |
| 5′Arm-mCher53BP | CTATGTCCCTCGTATGGTGAGCAAGGGCGAG | FP-53BP1 |
| 3′Arm-mCher53BP | GCAGGACAGCAGCAGATCTTTACTTGTACAGCTCGTCC | FP-53BP1 |
| mTur-BglII-3′Arm53BP | CAAGTAAAGATCTGCTGCTGTCCTGCTGTCC | 3′-Arm 53BP1 |
| pBlue-3′Arm53BP | ACTAAAGGGAACAAAAGCTGGGTACGTTGTTTTGGGGATAGGTAGAAG | 3′-Arm 53BP1 |
| pBlu-5′ArmRad54 | CGGTGGCGGCCGCTCTAGAAATCCACTCCTGTGGTCAC | 5′-Arm Rad54 |
| mVenus-BamHI-5′ArmRad54 | TGCTCACCATGGGAATCCCTCGCTGCTC | 5′-Arm Rad54 |
| 5′Arm-mVenusRad54 | AGGGATTCCCATGGTGAGCAAGGGCGAG | FP-Rad54 |
| 3′Arm-mVenusRad54 | AGTTAACCTGGGATCCTTATTTGTACAATTCGTCC | FP-Rad54 |
| mVenus-BamHI-3′ArmRad54 | ATAAGGATCCCAGGTTAACTGTCCCCTG | 3′-Arm Rad54 |
| 3Arm-Rad54PrimerR1 | AGGGAACAAAAGCTGGGTACCATTTCTCTCCTCTTAGGG | 3′-Arm Rad54L |
| Rad54-Probe | TGCAGTCCTTTCAGTGGCTA | Southern blot probe |
| Rad54-Probe | CAGCCTTTAACTGGGTCAGC | Southern blot probe |
| 53BP-Probe | ACATTGGCTGGTTGGATCTC | Southern blot probe |
| 53BP-Probe | GTGTTCGACAATGCTGATCC | Southern blot probe |
| ITS1 5′ | TCCGTAGGTGAACCTGCGG | Internal transcribed spacer |
| ITS4 5′ | TCCTCCGCTTATTGATATGC | Internal transcribed spacer |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lerksuthirat, T.; Wikiniyadhanee, R.; Chitphuk, S.; Stitchantrakul, W.; Sampattavanich, S.; Jirawatnotai, S.; Jumpathong, J.; Dejsuphong, D. DNA Repair Biosensor-Identified DNA Damage Activities of Endophyte Extracts from Garcinia cowa. Biomolecules 2020, 10, 1680. https://doi.org/10.3390/biom10121680
Lerksuthirat T, Wikiniyadhanee R, Chitphuk S, Stitchantrakul W, Sampattavanich S, Jirawatnotai S, Jumpathong J, Dejsuphong D. DNA Repair Biosensor-Identified DNA Damage Activities of Endophyte Extracts from Garcinia cowa. Biomolecules. 2020; 10(12):1680. https://doi.org/10.3390/biom10121680
Chicago/Turabian StyleLerksuthirat, Tassanee, Rakkreat Wikiniyadhanee, Sermsiri Chitphuk, Wasana Stitchantrakul, Somponnat Sampattavanich, Siwanon Jirawatnotai, Juangjun Jumpathong, and Donniphat Dejsuphong. 2020. "DNA Repair Biosensor-Identified DNA Damage Activities of Endophyte Extracts from Garcinia cowa" Biomolecules 10, no. 12: 1680. https://doi.org/10.3390/biom10121680