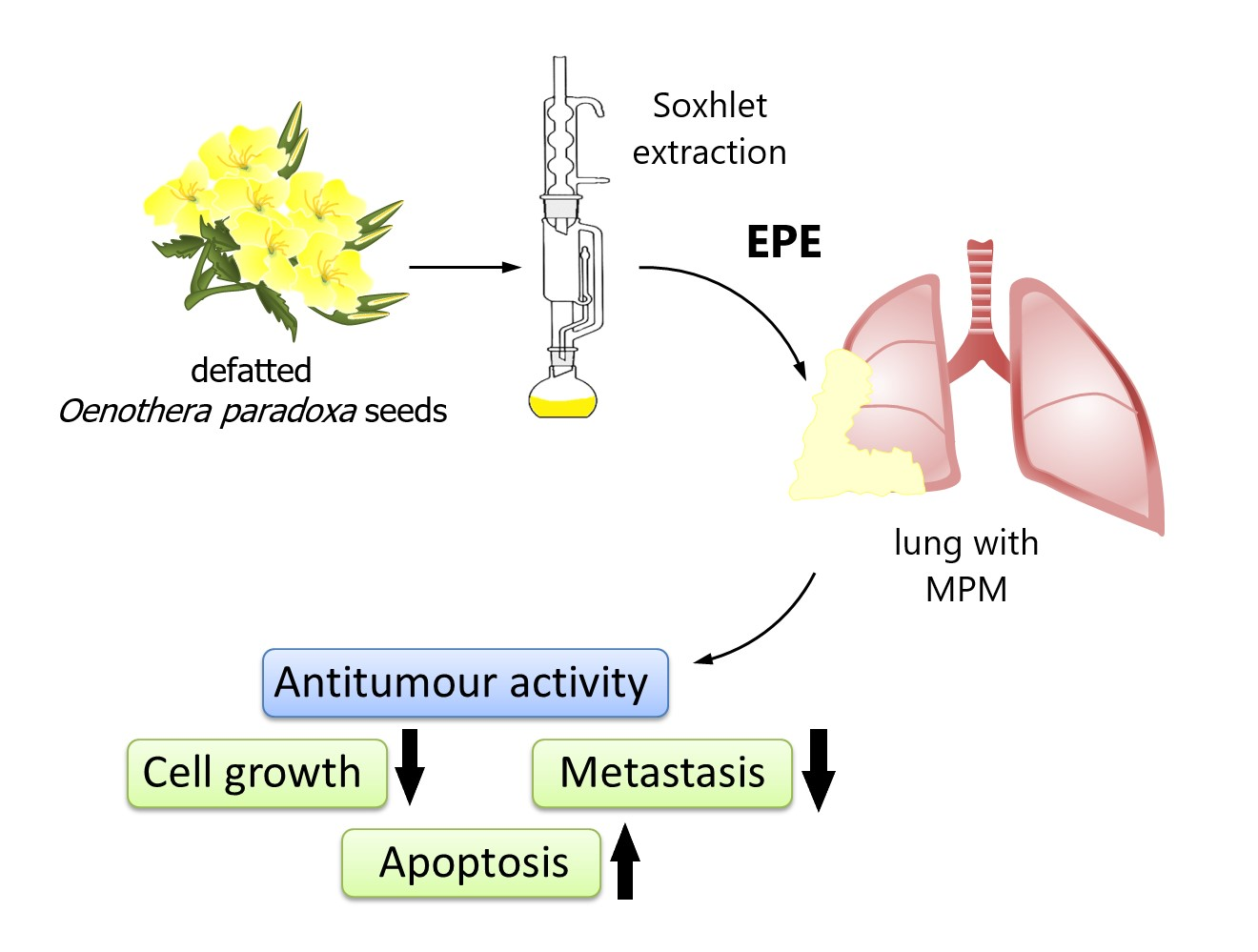
Polyphenol Extract from Evening Primrose (Oenothera paradoxa) Inhibits Invasion Properties of Human Malignant Pleural Mesothelioma Cells

 ,
,  , ,
, ,  , , , and
, , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Material and Extract Preparation
2.3. Characterization of Polyphenol Extract
2.4. Analysis of Phenolic Profile
2.5. Cell Culture and Treatment
2.6. Sulforhodamine B Assay
2.7. Wound-Healing Migration Assay
2.8. Transwell Invasion Assay
2.9. Apoptosis Analysis
2.10. Cell-Cycle Analysis
2.11. Statistical Analysis
3. Results
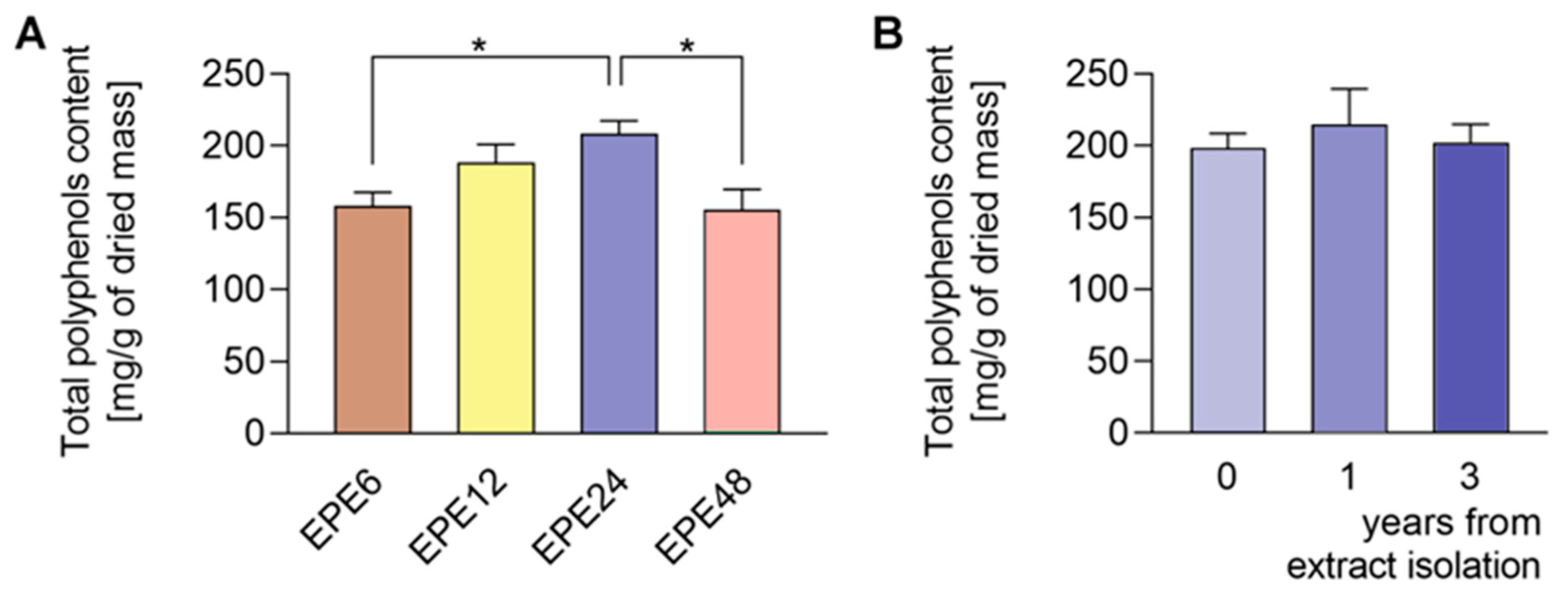
3.1. Qualitative and Quantitative Composition of EPE Extract
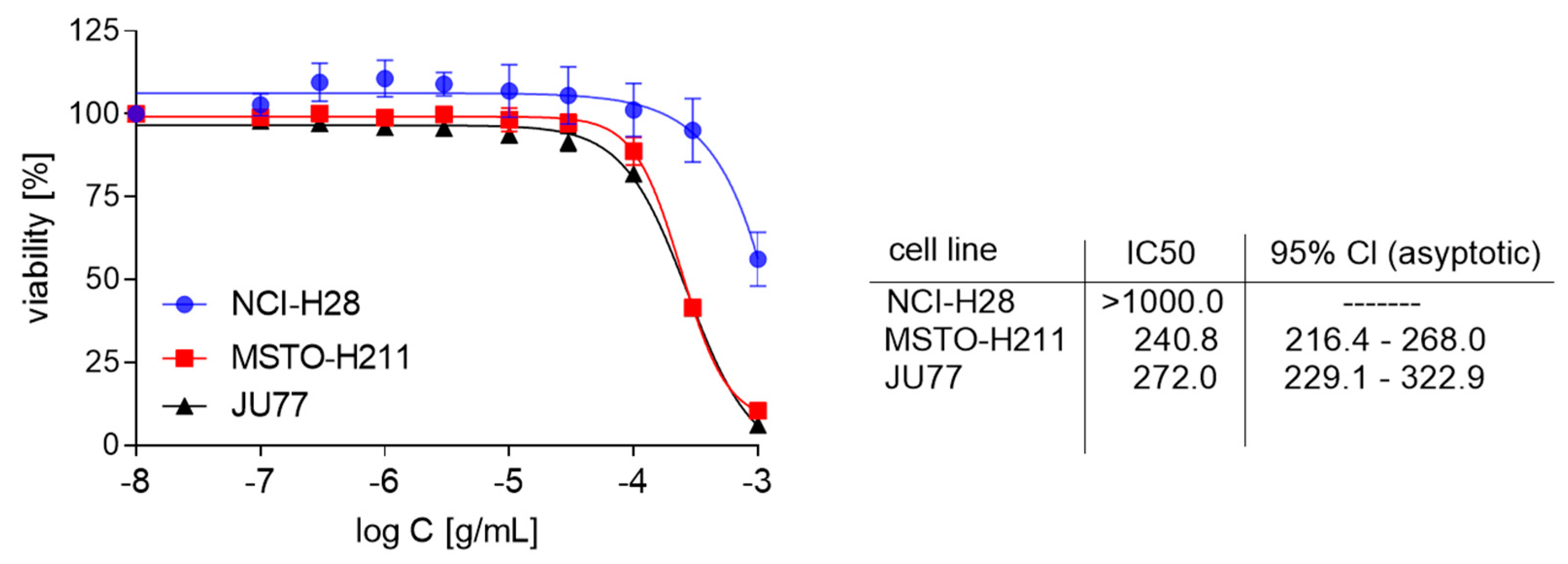
3.2. The Growth Inhibitory Effect of EPE on Mesothelioma Cells
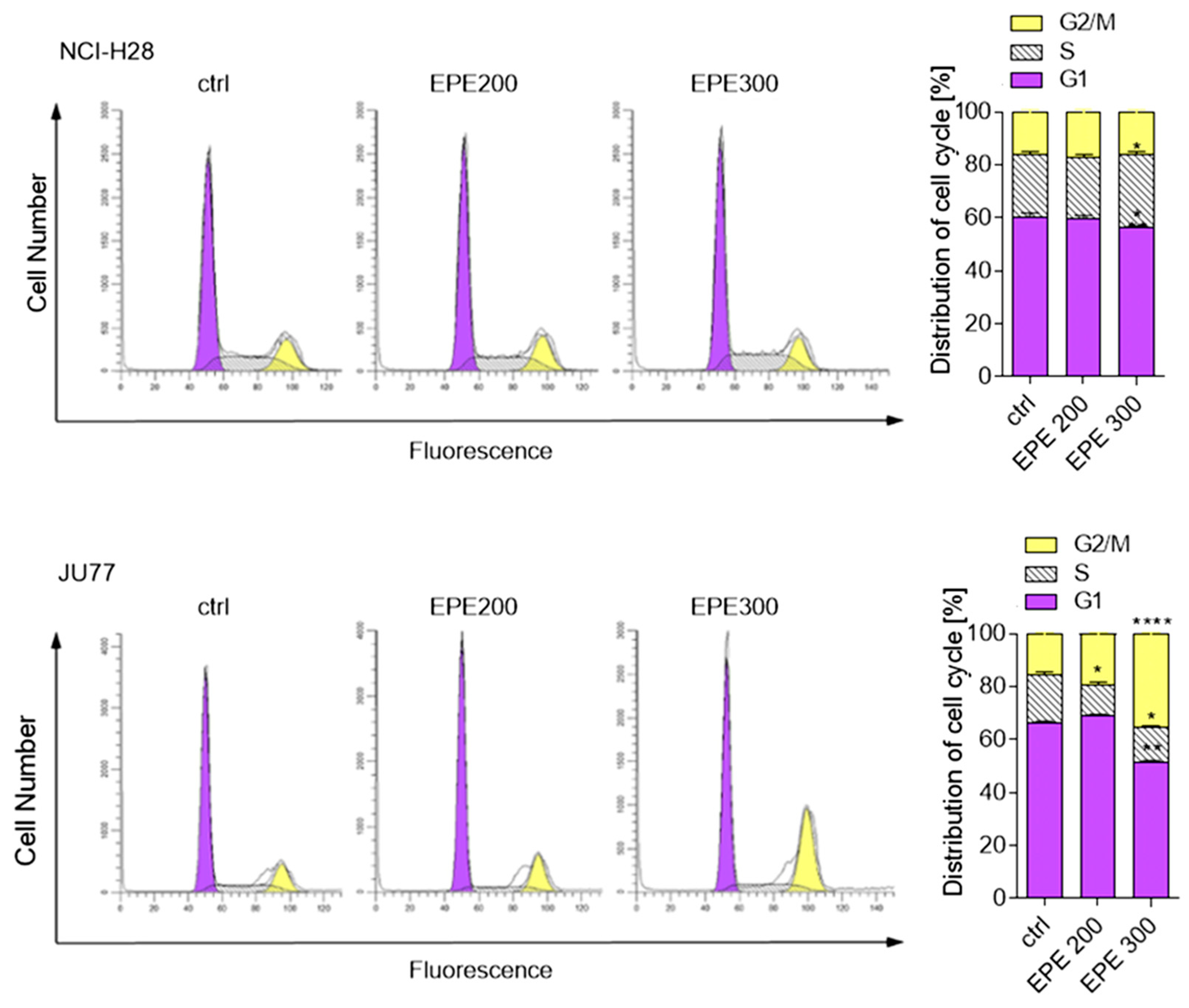
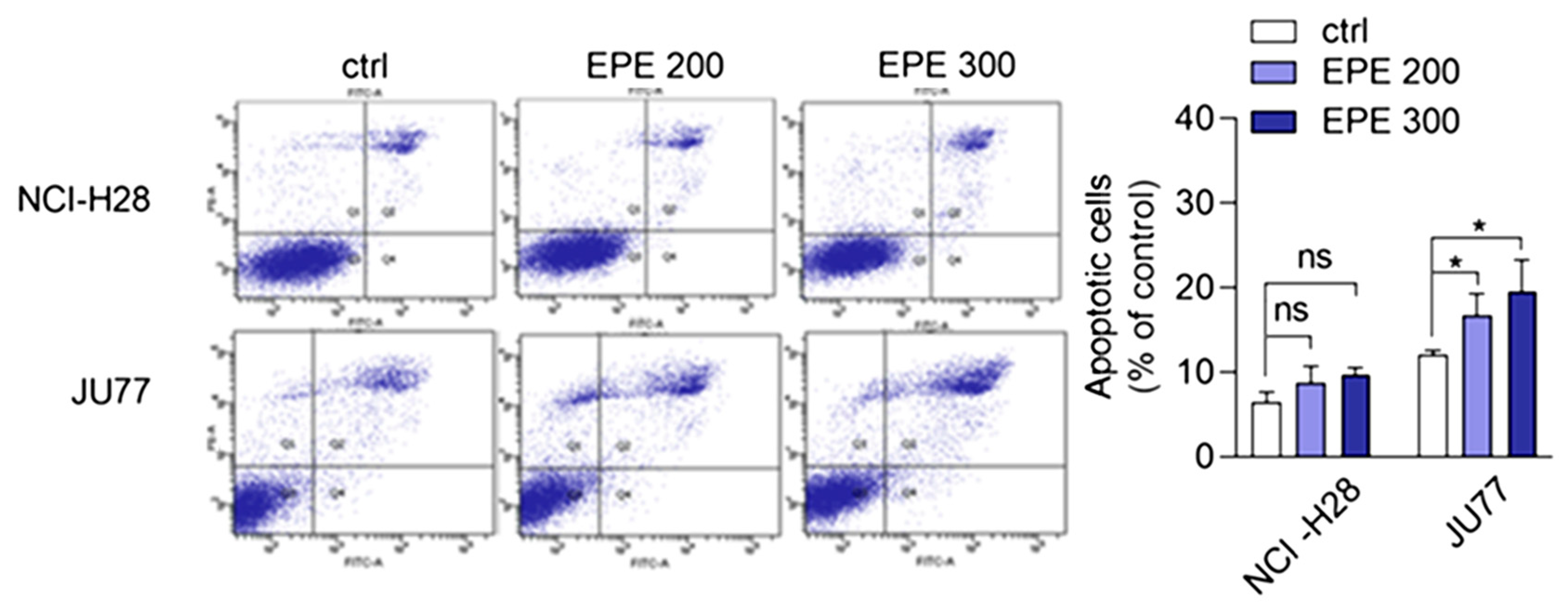
3.3. EPE Induced Cell Cycle Arrest and Apoptosis in JU77 Cells
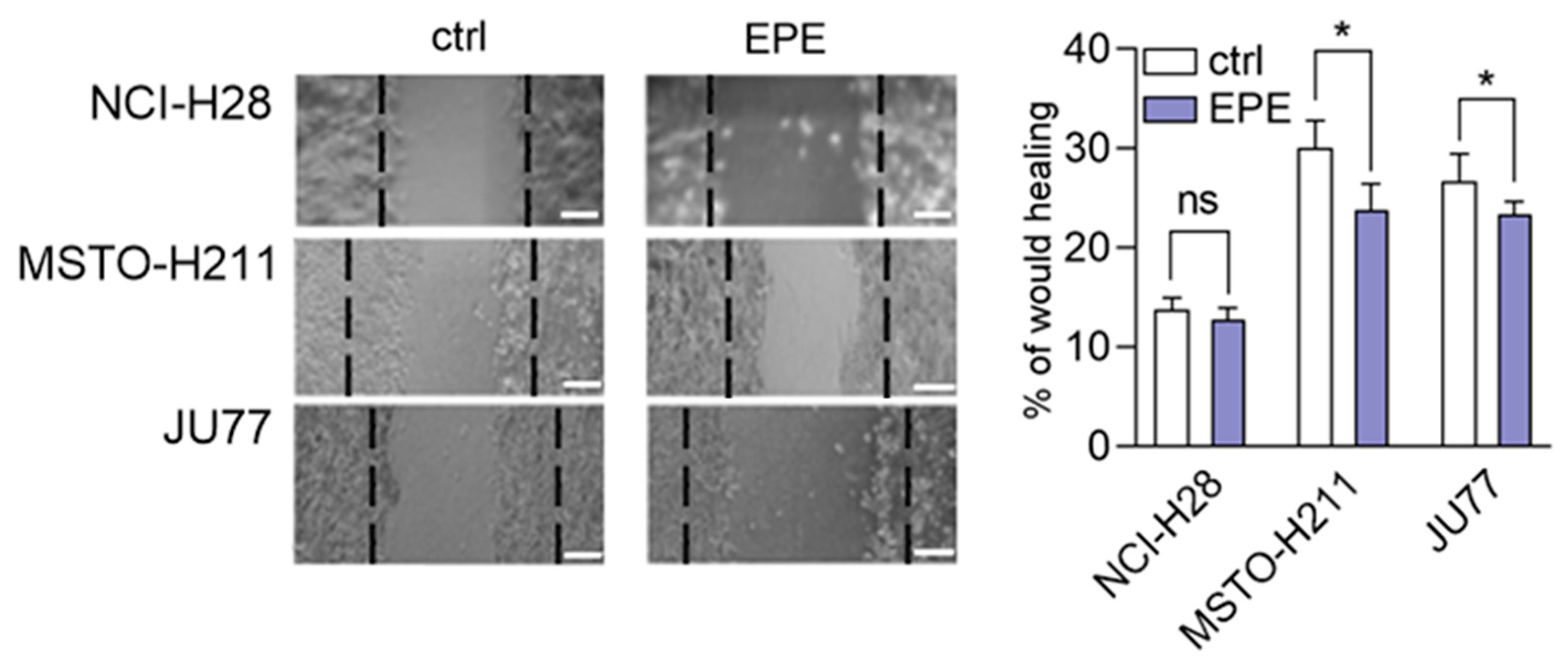
3.4. EPE Weakly Inhibited Cohort Cell Migration of MPM Cells Lines
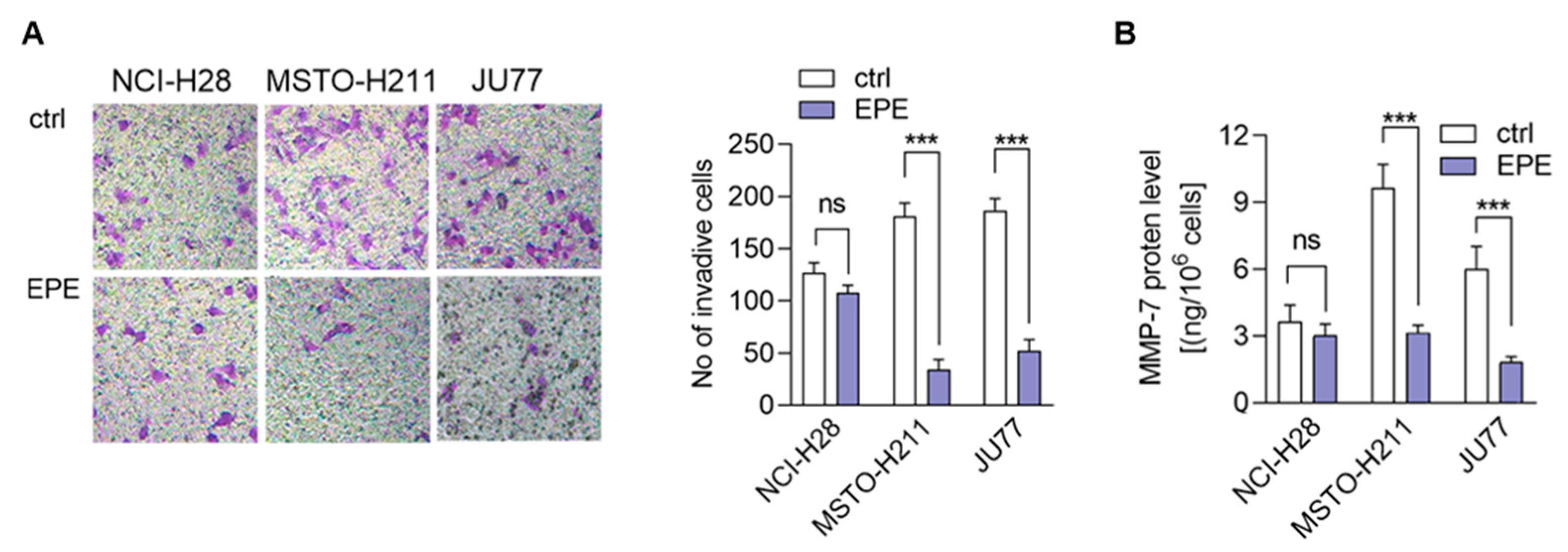
3.5. EPE Treatment Caused a Lower Invasion of More Invasive MPM Cells Lines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wojcik, M.; Burzynska-Pedziwiatr, I.; A Wozniak, L. A Review of natural and synthetic antioxidants important for health and longevity. Curr. Med. Chem. 2010, 17, 3262–3288. [Google Scholar] [CrossRef] [PubMed]
- Burzynska-Pedziwiatr, I.; Bukowiecka-Matusiak, M.; Wojcik, M.; Machala, W.; Bienkiewicz, M.; Spolnik, G.; Danikiewicz, W.; Wozniak, L.A. Dual stimulus-dependent effect of Oenothera paradoxa extract on the respiratory burst in human leukocytes: Suppressing for Escherichia coli and phorbol myristate acetate and stimulating for formyl-methionyl-leucyl-phenylalanine. Oxid. Med. Cell. Longev. 2014, 2014, 764367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Vallinas, M.; González-Castejón, M.; Rodríguez-Casado, A.; De Molina, A.R. Dietary phytochemicals in cancer prevention and therapy: A complementary approach with promising perspectives. Nutr. Rev. 2013, 71, 585–599. [Google Scholar] [CrossRef] [PubMed]
- Dalla Pellegrina, C.; Padovani, G.; Mainente, F.; Zoccatelli, G.; Bissoli, G.; Mosconi, S.; Veneri, G.; Peruffo, A.; Andrighetto, G.; Rizzi, C.; et al. Anti-tumour potential of a gallic acid-containing phenolic fraction from Oenothera biennis. Cancer Lett. 2005, 226, 17–25. [Google Scholar] [CrossRef]
- Zhang, J.; Li, L.; Kim, S.H.; Hagerman, A.E.; Lü, J. Anti-cancer, Anti-diabetic and other pharmacologic and Biological activities of penta-galloyl-glucose. Pharm. Res. 2009, 26, 2066–2080. [Google Scholar] [CrossRef] [Green Version]
- Ceci, C.; Lacal, P.M.; Tentori, L.; De Martino, M.G.; Miano, R.; Graziani, G. Experimental evidence of the antitumor, antimetastatic and antiangiogenic activity of ellagic acid. Nutrients 2018, 10, 1756. [Google Scholar] [CrossRef] [Green Version]
- Kuo, P.-L.; Hsu, Y.-L.; Lin, T.-C.; Chang, J.-K.; Lin, C.-C. Induction of cell cycle arrest and apoptosis in human non-small cell lung cancer A549 cells by casuarinin from the bark of Terminalia arjuna Linn. Anti Cancer Drugs 2005, 16, 409–415. [Google Scholar] [CrossRef]
- Hsu, J.D.; Kao, S.H.; Ou, T.T.; Chen, Y.J.; Li, Y.J.; Wang, C.J. Gallic acid induces G2/M phase arrest of breast cancer cell MCF-7 through stabilization of p27Kip1 attributed to disruption of p27Kip1/Skp2 complex. J. Agric. Food Chem. 2011, 59, 1996–2003. [Google Scholar] [CrossRef]
- Meng, M.; Liu, K.C.; Huang, A.C.; Wu, P.P.; Lin, H.Y.; Chueh, F.S.; Yang, J.S.; Lu, C.C.; Chiang, J.H.; Chung, J.G.; et al. Gallic acid suppresses the migration and invasion of PC-3 human prostate cancer cells via inhibition of matrix metalloproteinase-2 and-9 signaling pathways. Oncol. Rep. 2011, 26, 177–184. [Google Scholar] [CrossRef]
- Lo, C.; Lai, T.Y.; Yang, J.S.; Yang, J.H.; Ma, Y.S.; Weng, S.W.; Lin, H.Y.; Chen, H.Y.; Lin, J.G.; Chung, J.G.; et al. Gallic acid inhibits the migration and invasion of A375.S2 human melanoma cells through the inhibition of matrix metalloproteinase-2 and Ras. Melanoma Res. 2011, 21, 267–273. [Google Scholar] [CrossRef]
- Ji, B.C.; Hsu, W.H.; Yang, J.S.; Hsia, T.C.; Lu, C.C.; Chiang, J.H.; Yang, J.L.; Lin, C.H.; Lin, J.J.; Suen, L.J.W.; et al. Gallic acid induces apoptosis via caspase-3 and mitochondrion-dependent pathways in vitro and suppresses lung xenograft tumor growth in vivo. J. Agric. Food Chem. 2009, 57, 7596–7604. [Google Scholar] [CrossRef]
- Lu, Y.; Jiang, F.; Jiang, H.; Wu, K.; Zheng, X.; Cai, Y.; Katakowski, M.; Chopp, M.; To, S.S.T. Gallic acid suppresses cell viability, proliferation, invasion and angiogenesis in human glioma cells. Eur. J. Pharmacol. 2010, 641, 102–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, J.; Li, Y.; Gao, H.; Yang, D.; He, X.; Fang, Y.; Zhou, G. Phenolic compound ellagic acid inhibits mitochondrial respiration and tumor growth in lung cancer. Food Funct. 2020, 11, 6332–6339. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, B.A.; Re, G.G. IGF-II down regulation associated cell cycle arrest in colon cancer cells exposed to phenolic antioxidant ellagic acid. Anticancer. Res. 2001, 21, 359–364. [Google Scholar] [PubMed]
- Losso, J.N.; Bansode, R.R.; Trappey, A.; A Bawadi, H.; Truax, R. In vitro anti-proliferative activities of ellagic acid. J. Nutr. Biochem. 2004, 15, 672–678. [Google Scholar] [CrossRef]
- Umesalma, S.; Nagendraprabhu, P.; Sudhandiran, G. Ellagic acid inhibits proliferation and induced apoptosis via the Akt signaling pathway in HCT-15 colon adenocarcinoma cells. Mol. Cell. Biochem. 2014, 399, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Pitchakarn, P.; Chewonarin, T.; Ogawa, K.; Suzuki, S.; Asamoto, M.; Takahashi, S.; Shirai, T.; Limtrakul, P. Ellagic acid inhibits migration and invasion by prostate cancer cell lines. Asian Pac. J. Cancer Prev. 2013, 14, 2859–2863. [Google Scholar] [CrossRef] [Green Version]
- Ceci, C.; Tentori, L.; Atzori, M.G.; Lacal, P.M.; Bonanno, E.; Scimeca, M.; Cicconi, R.; Mattei, M.; De Martino, M.G.; Vespasiani, G.; et al. Ellagic Acid inhibits bladder cancer invasiveness and in vivo tumor growth. Nutrients 2016, 8, 744. [Google Scholar] [CrossRef]
- Zhang, L.; Duo, J.; Ying, G.G.; Wang, G. Quercetin inhibits human breast cancer cell proliferation and induces apoptosis via Bcl-2 and Bax regulation. Mol. Med. Rep. 2012, 5, 1453–1456. [Google Scholar] [CrossRef]
- Khan, N.; Afaq, F.; Saleem, M.; Ahmad, N.; Mukhtar, H. Targeting multiple signaling pathways by green tea polyphenol (−)-epigallocatechin-3-gallate. Cancer Res. 2006, 66, 2500–2505. [Google Scholar] [CrossRef] [Green Version]
- Neuhaus, T.; Pabst, S.; Stier, S.; Weber, A.A.; Schrör, K.; Sachinidis, A.; Vetter, H.; Ko, Y.D. Inhibition of the vascular-endothelial growth factor-induced intracellular signaling and mitogenesis of human endothelial cells by epigallocatechin-3 gallate. Eur. J. Pharmacol. 2004, 483, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska, U.; Szewczyk, K.; Owczarek, K.; Hrabec, Z.; Podsędek, A.; Sosnowska, D.; Hrabec, E. Procyanidins from evening primrose (Oenothera paradoxa) defatted seeds inhibit invasiveness of breast cancer cells and modulate the expression of selected genes involved in angiogenesis, metastasis, and apoptosis. Nutr. Cancer 2013, 65, 1219–1231. [Google Scholar] [CrossRef] [PubMed]
- Jaszewska, E.; Kosmider, A.; Kiss, A.K.; Naruszewicz, M. Oenothera paradoxa defatted seeds extract containing pentagalloylglucose and procyanidins potentiates the cytotoxicity of vincristine. J. Physiol. Pharmacol. 2010, 61, 637–643. [Google Scholar] [PubMed]
- Ettinger, D.S.; Akerley, W.; Borghaei, H.; Chang, A.; Cheney, R.T.; Chirieac, L.R.; D’Amico, T.A.; Demmy, T.L.; Ganti, A.K.P.; Govindan, R.; et al. Malignant pleural mesothelioma. J. Natl. Compr. Canc. Netw. 2012, 10, 26–41. [Google Scholar] [CrossRef] [Green Version]
- Howlader, N.; Noone, A.M.; Krapcho, M.; Garshell, J.; Miller, D.; Altekruse, S.F.; Kosary, C.L.; Yu, M.; Ruhl, J.; Tatalovich, Z.; et al. SEER Cancer Statistics Review, 1975–2015; National Cancer Institute: Bethesda, MA, USA, 2015. [Google Scholar]
- Hyland, R.A.; Ware, S.; Johnson, A.R.; Yates, D.H. Incidence trends and gender differences in malignant mesothelioma in New South Wales, Australia. Scand. J. Work. Environ. Health 2007, 33, 286–292. [Google Scholar] [CrossRef] [Green Version]
- Price, B.P. Mesothelioma Trends in the US: An update based on surveillance, epidemiology, and end results program data for 1973 through 2003. Am. J. Epidemiol. 2004, 159, 107–112. [Google Scholar] [CrossRef]
- Maurya, S.; Singh, D. Quantitative analysis of total phenolic content in Adhatoda vasica Nees extracts. Int. J. PharmTech Res. 2010, 2, 2403–2406. [Google Scholar]
- Gorlach, S.; Wagner, W.; Podsȩdek, A.; Sosnowska, D.; Dastych, J.; Koziołkiewicz, M. Polyphenols from evening primrose (Oenothera paradoxa) defatted seeds induce apoptosis in human colon cancer caco-2 cells. J. Agric. Food Chem. 2011, 59, 6985–6997. [Google Scholar] [CrossRef]
- Jaszewska, E.; Kosśmider, A.; Kiss, A.K.; Naruszewicz, M. Pro-oxidative and pro-apoptotic action of defatted seeds of Oenothera paradoxa on human skin melanoma cells. J. Agric. Food Chem. 2009, 57, 8282–8289. [Google Scholar] [CrossRef]
- Kiss, A.K.; Naruszewicz, M. Polyphenolic compounds characterization and reactive nitrogen species scavenging capacity of Oenothera paradoxa defatted seed extracts. Food Chem. 2012, 131, 485–492. [Google Scholar] [CrossRef]
- Lu, X.; Errington, J.; Curtin, N.J.; Lunec, J.; Newell, D.R. The impact of p53 status on cellular sensitivity to antifolate drugs. Clin. Cancer Res. 2001, 7, 2114–2123. [Google Scholar] [PubMed]
- Plażuk, D.; Wieczorek, A.; Ciszewski, W.M.; Kowalczyk, K.; Błauż, A.; Pawlędzio, S.; Makal, A.; Eurtivong, C.; Arabshahi, H.J.; Reynisson, J.; et al. Synthesis and in vitro biological evaluation of ferrocenyl side-chain-functionalized paclitaxel derivatives. ChemMedChem 2017, 12, 1882–1892. [Google Scholar] [CrossRef] [PubMed]
- Timoszuk, M.; Bielawska, K.; Skrzydlewska, E. Evening primrose (Oenothera biennis) biological activity dependent on chemical composition. Antioxidants 2018, 7, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Middleton, E.; Kandaswami, C.; Theoharides, T.C. The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharmacol. Rev. 2000, 52, 673–675. [Google Scholar]
- Fantini, M.; Benvenuto, M.; Masuelli, L.; Frajese, G.V.; Tresoldi, I.; Modesti, A.; Bei, R. In vitro and in vivo antitumoral effects of combinations of polyphenols, or polyphenols and anticancer drugs: Perspectives on cancer treatment. Int. J. Mol. Sci. 2015, 16, 9236–9282. [Google Scholar] [CrossRef] [Green Version]
- Kiss, A.K.; Kaplon-Cieslicka, A.; Filipiak, K.J.; Opolski, G.; Naruszewicz, M. Ex vivo effects of an Oenothera paradoxa extract on the reactive oxygen species generation and neutral endopeptidase activity in neutrophils from patients after acute myocardial infarction. Phytotherapy Res. 2011, 26, 482–487. [Google Scholar] [CrossRef]
- Granica, S.; Czerwińska, M.E.; Piwowarski, J.P.; Ziaja, M.; Kiss, A.K. Chemical composition, antioxidative and Anti-inflammatory activity of extracts prepared from aerial parts of Oenothera biennis L. and Oenothera paradoxa hudziok obtained after seeds cultivation. J. Agric. Food Chem. 2013, 61, 801–810. [Google Scholar] [CrossRef]
- Sałaga, M.; Lewandowska, U.; Sosnowska, D.; Zakrzewski, P.K.; Cygankiewicz, A.I.; Piechota-Polańczyk, A.; Sobczak, M.; Mosinska, P.; Chen, C.; Krajewska, W.M.; et al. Polyphenol extract from evening primrose pomace alleviates experimental colitis after intracolonic and oral administration in mice. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2014, 387, 1069–1078. [Google Scholar] [CrossRef] [Green Version]
- Christie, W.W. The analysis of evening primrose oil. Ind. Crop. Prod. 1999, 10, 73–83. [Google Scholar] [CrossRef]
- Lewandowska, U.; Owczarek, K.; Szewczyk, K.; Podsędek, A.; Koziolkiewicz, M.; Hrabec, E. Influence of polyphenol extract from evening primrose (Oenothera paradoxa) seeds on human prostate and breast cancer cell lines. Postępy Higieny i Medycyny Doświadczalnej 2014, 68, 110–118. [Google Scholar] [CrossRef]
- Azwanida, N.N. A review on the extraction methods use in medicinal plants, principle, strength and limitation. Med. Aromat. Plants 2015, 4, 2167–2412. [Google Scholar]
- De Castro, M.L.; García-Ayuso, L. Soxhlet extraction of solid materials: An outdated technique with a promising innovative future. Anal. Chim. Acta 1998, 369, 1–10. [Google Scholar] [CrossRef]
- Zhang, Q.W.; Lin, L.G.; Ye, W.C. Techniques for extraction and isolation of natural products: A comprehensive review. Chin. Med. 2018, 13, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Castro, M.D.L.; Priego-Capote, F. Soxhlet extraction: Past and present panacea. J. Chromatogr. A 2010, 1217, 2383–2389. [Google Scholar] [CrossRef] [PubMed]
- Kiss, A.K.; Filipek, A.; Czerwinska, M.; Naruszewicz, M. Oenothera paradoxa defatted seeds extract and ist bioactive component penta-O-galloyl-β-d-glucose decreased production of reactive oxygen species and infibited release of leukotriene B4, interleukin-8, elastase, and myeloperoxidase in human neutrophils. J. Agirc. Food Chemi. 2010, 58, 9960–9966. [Google Scholar] [CrossRef] [PubMed]
- Oh, G.S.; Pae, H.O.; Oh, H.; Hong, S.G.; Kim, I.K.; Chai, K.Y.; Yun, Y.G.; Kwon, T.O.; Chung, H.T. In vitro anti-proliferative effect of 1,2,3,4,5-penta-O-galloyl-beta-d-glucose on human hepatocellular carcinoma cell line, SK-HEP-1 cells. Cancer Lett. 2001, 174, 17–24. [Google Scholar] [CrossRef]
- Ryu, H.G.; Jeong, S.J.; Kwon, H.Y.; Lee, H.J.; Lee, E.O.; Lee, M.H.; Choi, S.H.; Ahn, K.S.; Kim, S.H. Penta-O-galloyl-β-d-glucose attenuates cisplatin-induced nephrotoxicity via reactive oxygen species reduction in renal epithelial cells and enhances antitumor activity in Caki-2 renal cancer cells. Toxicol. Vitr. 2012, 26, 206–214. [Google Scholar] [CrossRef]
- Niedzwiecki, A.; Roomi, M.W.; Kalinovsky, T.; Rath, M. Anticancer efficacy of polyphenols and their combinations. Nutrientrs 2016, 8, 552. [Google Scholar] [CrossRef] [Green Version]
- Shim, J.H.; Lee, K.A.; Lee, Y.J.; Ban, J.O.; Lee, S.H.; Cho, M.K.; Nam, H.S.; Hong, J.T. The flavonoid resveratrol suppresses growth of human malignant pleural mesothelioma cells through direct inhibition of specificity protein 1. Int. J. Mol. Med. 2012, 30, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Chae, J.I.; Cho, J.H.; Lee, K.A.; Choi, N.J.; Seo, K.S.; Kim, S.B.; Lee, S.H.; Shim, J.H. Role of transcription factor Sp1 in the quercetin-mediated inhibitory effect on human malignant pleural mesothelioma. Int. J. Mol. Med. 2012, 30, 835–841. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.M.; Thompson, J.K.; MacPherson, M.B.; Beuschel, S.L.; Westbom, C.M.; Sayan, M.; Shukla, A. Curcumin: A double hit on malignant mesothelioma. Cancer Prev. Res. 2014, 7, 330–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valenti, D.; De Bari, L.; Manente, G.A.; Rossi, L.; Mutti, L.; Moro, L.; Vacca, R.A. Negative modulation of mitochondrial oxidative phosphorylation by epigallocatechin-3 gallate leads to growth arrest and apoptosis in human malignant pleural mesothelioma cells. Biochimica et Biophysica Acta Mol. Basis Dis. 2013, 1832, 2085–2096. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska, U.; Owczarek, K.; Szewczyk, K.; Sosnowska, D.; Koziolkiewicz, M.; Hrabec, E. Differentiated impact of procyanidins from evening primrose on human breast cancer cells. Open Life Sci. 2014, 9, 647–658. [Google Scholar] [CrossRef] [Green Version]
- American Cancer Society. Malignant Mesothelioma Early Detection, Diagnosis, and Staging; American Cancer Society: Atlanta, GA, USA, 2018. [Google Scholar]
- Fecker, R.; Buda, V.; Alexa, E.; Avram, S.; Pavel, I.Z.; Muntean, D.; Ileana, C.; Watz, C.; Minda, D.; Dehelean, C.; et al. Phytochemical and biological screening of Oenothera biennis L. hydroalcoholic extract. Biomol. 2020, 10, 818. [Google Scholar] [CrossRef] [PubMed]
- Dawczynski, C.; Hackermeier, U.; Viehweger, M.; Stange, R.; Springer, M.; Jahreis, G. Incorporation of n-3 PUFA and gamma-linolenic acid in blood lipids and red blood cell lipids together with their influence on disease activity in patients with chronic inflammatory arthritis—A randomized controlled human intervention trial. Lipids Health Dis. 2011, 10, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taweechaisupapong, S.; Srisuk, N.; Nimitpornsuko, C.; Vattraphoudes, T.; Rattanayatikul, C.; Godfrey, K. Evening primrose oil effects on osteoclasts during tooth movement. Angle Orthod. 2005, 75, 356–361. [Google Scholar]
- Owczarek, K.; Hrabec, E.; Fichna, J.; Sosnowska, D.; Koziołkiewicz, M.; Szymański, J.; Lewandowska, U. Inhibition of nuclear factor-kappaB, cyclooxygenase-2, and metalloproteinase-9 expression by flavanols from evening primrose (Oenothera paradoxa) in human colon cancer SW-480 cells. J. Funct. Foods 2017, 37, 553–563. [Google Scholar] [CrossRef]
- Lewandowska, U.; Szewczyk, K.; Owczarek, K.; Hrabec, Z.; Podsedek, A.; Koziołkiewicz, M.; Hrabec, E. Flavanols from evening primrose (Oenothera paradoxa) defatted seeds inhibit prostate cells invasiveness and cause changes in Bcl-2/Bax mRNA ratio. J. Agric. Food Chem. 2013, 61, 2987–2998. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spectrophotometric Analysis (n = 3) A | ||||||
|---|---|---|---|---|---|---|
| Total Polyphenols Content (mg/g) | 208.24 ± 8.92 | |||||
| Qualitative microLC-TOF 4600 MS Analysis B | ||||||
| No. a | tR (min) b | Assigned identity c | MW d | [M − H]¯ (m/z) e | Chemical Entity f | MS/MS Ions (m/z) g |
| 1. | 0.52 | Gallic acid | 170.12 | 169.0134 | Phenolic acid | 125.0235, 107.0136 |
| 2. | 0.52 | Caffeic acid | 180.16 | 179.0546 | Phenolic acid | 161.0432, 119.0325, 89.0238 |
| 3. | 0.53 | Digalloyl glucose | 484.37 | 483.0782 | Hydrolyzable tannins | 331.0660, 313.0562, 169.0136 |
| 4. | 0.58 | Procyanidin dimer | 578.52 | 577.1369 | Flavan-3-ol | 451.1028, 425.0867, 289.0706 |
| 5. | 0.91 | Catechin | 290.26 | 289.0708 | Flavan-3-ol | 245.0805, 109.0292 |
| 6. | 0.98 | Quercetin pentoside | 434.09 | 433.1120 | Flavonols | 343.0803, 181.0496 |
| 7. | 1.06 | Protocatechuic acid | 154.12 | 152.9175 | Phenolic acid | 135.9133, 122.0367, 109.0291 |
| 8 | 1.10 | Procyanidin tetramer | 1154.36 | 1153.2692 | Flavon-3-ol | 863.1865, 575.1145, 287.0532 |
| 9 | 1.16 | Procyanidin trimer | 866.21 | 865.1989 | Flavan-3-ol | 695.1420, 577.1371, 287.0514 |
| 10 | 1.20 | Procyanidin trimer gallate | 1018.34 | 1017.2184 | Flavan-3-ol | 865.2041, 287.0569 |
| 11. | 1.21 | Quercetin/Ellagic acid | 302.24/302.20 | 300.9980 | Phenolic acid | 286.0818, 257.0354, 185.0230 |
| 12. | 1.28 | Penta-O-galloyl β-d-glucose | 940.68 | 939.1179 | Hydrolyzable tannins | 787.1009, 769.0939, 617.0820 |
| 13. | 1.32 | Procyanidin dimer gallate | 730.15 | 729.1476 | Flavan-3-ol | 577.1382, 559.1232, 407.0792, 289.0707 |
| 14. | 1.64 | Tetragalloyl glucose | 788.60 | 787.1025 | Hydrolyzable tannins | 617.0801, 465.0645, 295.0421 |
| 15. | 1.71 | Methyl gallate | 184.15 | 183.1025 | Phenolic acid | 139.1131, 111.0821 |
| LC-TripleTOF-MS Analysis (n = 3) | |||||
|---|---|---|---|---|---|
| Compound | Structure | RT | m/z | mg/g | |
| gallic acid |  | 0.97 | 169.0501 # | 51.40 ± 0.13 | |
| catechin |  | 2.03 | 291.0917 * | 23.23 ± 0.09 | |
| procyanidin B2 |  | 2.15 | 579.1384 * | 0.98 ± 0.28 | |
| penta-O-galloyl-β-d-glucose |  | 2.23 | 939.1206 # | 3.70 ± 0.06 | |
| ellagic acid |  | 2.25 | 303.0145 * | 77.67 ± 0.48 | |
| epicatechin gallate |  | 2.41 | 443.0941 * | 3.96 ± 0.01 | |
| caffeic acid |  | 2.49 | 181.0501 * | 1.31 ± 0.02 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chmielewska-Kassassir, M.; Sobierajska, K.; Ciszewski, W.M.; Bukowiecka-Matusiak, M.; Szczesna, D.; Burzynska-Pedziwiatr, I.; Wiczkowski, W.; Wagner, W.; Wozniak, L.A. Polyphenol Extract from Evening Primrose (Oenothera paradoxa) Inhibits Invasion Properties of Human Malignant Pleural Mesothelioma Cells. Biomolecules 2020, 10, 1574. https://doi.org/10.3390/biom10111574
Chmielewska-Kassassir M, Sobierajska K, Ciszewski WM, Bukowiecka-Matusiak M, Szczesna D, Burzynska-Pedziwiatr I, Wiczkowski W, Wagner W, Wozniak LA. Polyphenol Extract from Evening Primrose (Oenothera paradoxa) Inhibits Invasion Properties of Human Malignant Pleural Mesothelioma Cells. Biomolecules. 2020; 10(11):1574. https://doi.org/10.3390/biom10111574
Chicago/Turabian StyleChmielewska-Kassassir, Malgorzata, Katarzyna Sobierajska, Wojciech M. Ciszewski, Malgorzata Bukowiecka-Matusiak, Dorota Szczesna, Izabela Burzynska-Pedziwiatr, Wieslaw Wiczkowski, Waldemar Wagner, and Lucyna A. Wozniak. 2020. "Polyphenol Extract from Evening Primrose (Oenothera paradoxa) Inhibits Invasion Properties of Human Malignant Pleural Mesothelioma Cells" Biomolecules 10, no. 11: 1574. https://doi.org/10.3390/biom10111574