Discovery of New Antibacterial Accramycins from a Genetic Variant of the Soil Bacterium, Streptomyces sp. MA37




Abstract
:1. Introduction
2. Materials and Methods
2.1. General Experimental Procedures
2.2. Strain, Genomic DNA, and Media
2.3. Construction of Knockout Vector
2.4. Conjugations between Escherichia coli and Streptomyces sp. MA37
2.5. Screening for the Mutant Strain
2.6. Metabolic Profile of Mutant and WT Strains
2.7. Fermentation, Extraction, Metabolite Screening
2.8. Semi-Prep HPLC Isolation
2.9. Minimum Inhibitory Concentration
2.10. GenBank Accession Number
3. Results and Discussion
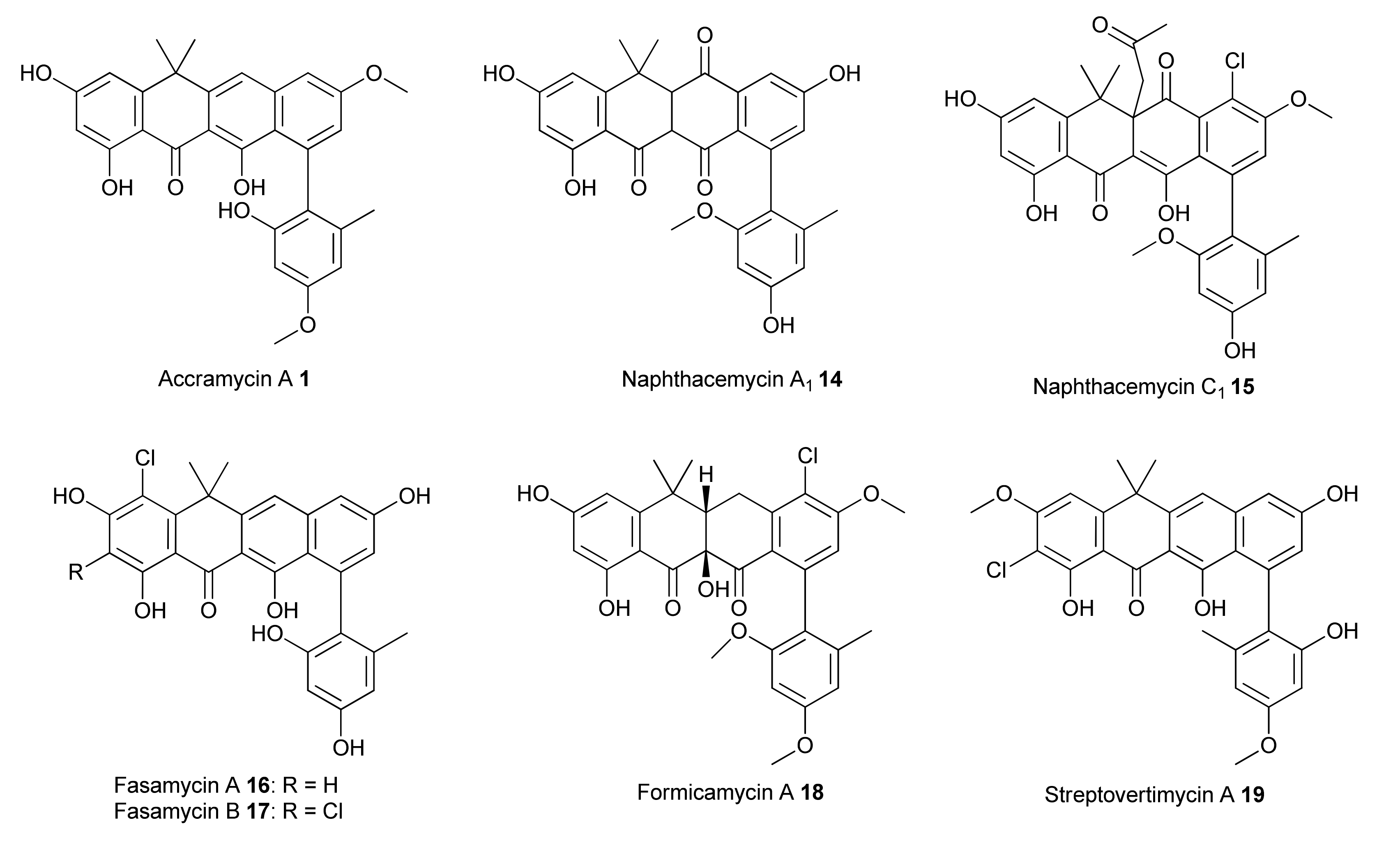
3.1. Structure Elucidation
3.2. Biological Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fukumoto, A.; Kim, Y.P.; Iwatsuki, M.; Hirose, T.; Sunazuka, T.; Hanaki, H.; Omura, S.; Shiomi, K. Naphthacemycins, novel circumventors of β-lactam resistance in MRSA, produced by Streptomyces sp. KB-3346-5. II. Structure elucidation. J. Antibiot. 2017, 70, 568–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huo, C.; Zheng, Z.; Xu, Y.; Ding, Y.; Zheng, H.; Mu, Y.; Niu, Y.; Gao, J.; Lu, X. Naphthacemycins from a Streptomyces sp. as Protein-Tyrosine Phosphatase Inhibitors. J. Nat. Prod. 2020, 83, 1394–1399. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Lu, X.; Zhu, J.; Mu, Y.; Xu, Y.; Gao, J.; Zhang, X.; Zheng, Z. Discovery of naphthacemycins as a novel class of PARP1 inhibitors. Bioorg. Med. Chem. Lett. 2019, 29, 1904–1908. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.; Munnoch, J.T.; Devine, R.; Holmes, N.A.; Seipke, R.F.; Wilkinson, K.A.; Wilkinson, B.; Hutchings, M.I. Formicamycins, antibacterial polyketides produced by Streptomyces formicae isolated from African Tetraponera plant-ants. Chem. Sci. 2017, 8, 3218–3227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Z.; Chakraborty, D.; Dewell, S.B.; Reddy, B.V.B.; Brady, S.F. Environmental DNA-encoded antibiotics fasamycins A and B inhibit FabF in type II fatty acid biosynthesis. J. Am. Chem. Soc. 2012, 134, 2981–2987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Z.; Kallifidas, D.; Brady, S.F. Functional analysis of environmental DNA-derived type II polyketide synthases reveals structurally diverse secondary metabolites. Proc. Natl. Acad. Sci. USA 2011, 108, 12629–12634. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Li, X.; Wu, P.; Xue, J.; Xu, L.; Li, H.; Wei, X. Streptovertimycins A–H, new fasamycin-type antibiotics produced by a soil-derived Streptomyces morookaense strain. J. Antibiot. 2020, 73, 283–289. [Google Scholar] [CrossRef]
- Hirose, T.; Kojima, Y.; Matsui, H.; Hanaki, H.; Iwatsuki, M.; Shiomi, K.; Omura, S.; Sunazuka, T. Total synthesis of (±)-naphthacemycin A 9, possessing both antibacterial activity against methicillin-resistant Staphylococcus aureus and circumventing effect of β-lactam resistance. J. Antibiot. 2017, 70, 574–581. [Google Scholar]
- Huang, J.K.; Yang Lauderdale, T.L.; Lin, C.C.; Shia, K.S. Total Synthesis of tetarimycin A, (±)-naphthacemycin A9, and (±)-fasamycin A: Structure-activity relationship studies against drug-resistant bacteria. J. Org. Chem. 2018, 83, 6508–6523. [Google Scholar] [CrossRef]
- Jiang, D.; Xin, K.; Yang, B.; Chen, Y.; Zhang, Q.; He, H.; Gao, S. Total synthesis of three families of natural antibiotics: Anthrabenzoxocinones, fasamycins/naphthacemycins, and benastatins. CCS Chem. 2020, 2, 800–812. [Google Scholar] [CrossRef]
- Huang, S.; Tabudravu, J.; Elsayed, S.S.; Travert, J.; Peace, D.; Tong, M.H.; Kyeremeh, K.; Kelly, S.M.; Trembleau, L.; Ebel, R.; et al. Discovery of a single monooxygenase that catalyzes carbamate formation and ring contraction in the biosynthesis of the legonmycins. Angew. Chem. Int. Ed. 2015, 54, 12697–12701. [Google Scholar] [CrossRef] [PubMed]
- Maglangit; Fang; Kyeremeh; Sternberg; Ebel; Deng A co-culturing approach enables discovery and biosynthesis of a bioactive indole alkaloid metabolite. Molecules 2020, 25, 256. [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Su, L.; Fang, Q.; Tabudravu, J.; Yang, X.; Rickaby, K.; Trembleau, L.; Kyeremeh, K.; Deng, Z.; Deng, H.; et al. Enzymatic reconstitution and biosynthetic investigation of the bacterial carbazole neocarazostatin A. J. Org. Chem. 2019, 84, 16323–16328. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Lv, M.; Kyeremeh, K.; Deng, Z.; Deng, H.; Yu, Y. A ThDP-dependent enzymatic carboligation reaction involved in neocarazostatin A tricyclic carbazole formation. Org. Biomol. Chem. 2016, 14, 8679–8684. [Google Scholar] [CrossRef] [Green Version]
- Su, L.; Zhang, R.; Kyeremeh, K.; Deng, Z.; Deng, H.; Yu, Y. Dissection of the neocarazostatin: A C4 alkyl side chain biosynthesis by in vitro reconstitution. Org. Biomol. Chem. 2017, 15, 3843–3848. [Google Scholar] [CrossRef]
- Huang, S.; Elsayed, S.S.; Lv, M.; Tabudravu, J.; Rateb, M.E.; Gyampoh, R.; Kyeremeh, K.; Ebel, R.; Jaspars, M.; Deng, Z.; et al. Biosynthesis of neocarazostatin A reveals the sequential carbazole prenylation and hydroxylation in the tailoring steps. Chem. Biol. 2015, 22, 1633–1642. [Google Scholar] [CrossRef] [PubMed]
- Maglangit, F.; Alrashdi, S.; Renault, J.; Trembleau, L.; Victoria, C.; Tong, M.H.; Wang, S.; Kyeremeh, K.; Deng, H. Characterization of the promiscuous N-acyl CoA transferase, LgoC, in legonoxamine biosynthesis. Org. Biomol. Chem. 2020, 18, 2219–2222. [Google Scholar] [CrossRef]
- Maglangit, F.; Tong, M.H.; Jaspars, M.; Kyeremeh, K.; Deng, H. Legonoxamines A-B, two new hydroxamate siderophores from the soil bacterium, Streptomyces sp. MA37. Tetrahedron Lett. 2019, 60, 75–79. [Google Scholar] [CrossRef]
- Wu, L.; Tong, M.H.; Raab, A.; Fang, Q.; Wang, S.; Kyeremeh, K.; Yu, Y.; Deng, H. An unusual metal-bound 4-fluorothreonine transaldolase from Streptomyces sp. MA37 catalyses promiscuous transaldol reactions. Appl. Microbiol. Biotechnol. 2020, 104, 3385–3396. [Google Scholar] [CrossRef] [Green Version]
- Deng, H.; Ma, L.; Bandaranayaka, N.; Qin, Z.; Mann, G.; Kyeremeh, K.; Yu, Y.; Shepherd, T.; Naismith, J.H.; O’Hagan, D. Identification of fluorinases from Streptomyces sp. MA37, Norcardia brasiliensis, and Actinoplanes sp. N902-109 by genome mining. ChemBioChem 2014, 15, 364–368. [Google Scholar] [CrossRef]
- Ma, L.; Bartholome, A.; Tong, M.H.; Qin, Z.; Yu, Y.; Shepherd, T.; Kyeremeh, K.; Deng, H.; O’Hagan, D.; Su, L.; et al. Identification of a fluorometabolite from Streptomyces sp. MA37: (2R3S4S)-5-fluoro-2,3,4-trihydroxypentanoic acid. Chem. Sci. 2017, 14, 8679–8684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Maglangit, F.; Deng, H. Fluorine biocatalysis. Curr. Opin. Chem. Biol. 2020, 55, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Deng, H. Defluorination of 4-fluorothreonine by threonine deaminase. Org. Biomol. Chem. 2020, 18, 6236–6240. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Tong, M.H.; Kyeremeh, K.; Deng, H. Identification of 5-fluoro-5-deoxy-ribulose as a Shunt fluorometabolite in Streptomyces sp. MA37. Biomolecules 2020, 10, 1023. [Google Scholar] [CrossRef] [PubMed]
- Maglangit, F.; Fang, Q.; Leman, V.; Soldatou, S.; Ebel, R.; Kyeremeh, K.; Deng, H. Accramycin A, a new aromatic polyketide, from the soil bacterium, Streptomyces sp. MA37. Molecules 2019, 24, 3384. [Google Scholar] [CrossRef] [Green Version]
- Fukumoto, A.; Kim, Y.P.; Matsumoto, A.; Takahashi, Y.; Suzuki, M.; Onodera, H.; Tomoda, H.; Matsui, H.; Hanaki, H.; Iwatsuki, M.; et al. Naphthacemycins, novel circumventors of β-lactam resistance in MRSA, produced by Streptomyces sp. KB-3346-5. I. The taxonomy of the producing strain, and the fermentation, isolation and antibacterial activities. J. Antibiot. 2017, 70, 562–567. [Google Scholar] [CrossRef] [Green Version]
- Devine, R.; Mcdonald, H.; Qin, Z.; Arnold, C.; Noble, K.; Chandra, G.; Wilkinson, B.; Hutchings, M.I. Refactoring the formicamycin biosynthetic gene cluster to make high-level producing strains and new molecules. BioRxiv 2020, 1–30. [Google Scholar] [CrossRef]
- Wagner, T.; Joshi, B.; Janice, J.; Askarian, F.; Škalko-Basnet, N.; Hagestad, O.C.; Mekhlif, A.; Wai, S.N.; Hegstad, K.; Johannessen, M. Enterococcus faecium produces membrane vesicles containing virulence factors and antimicrobial resistance related proteins. J. Proteom. 2018, 187, 28–38. [Google Scholar] [CrossRef]
- Rosvoll, T.C.S.; Lindstad, B.L.; Lunde, T.M.; Hegstad, K.; Aasnæs, B.; Hammerum, A.M.; Lester, C.H.; Simonsen, G.S.; Sundsfjord, A.; Pedersen, T. Increased high-level gentamicin resistance in invasive Enterococcus faecium is associated with aac(6′)Ie-aph(2″)Ia-encoding transferable megaplasmids hosted by major hospital-adapted lineages. FEMS Immunol. Med. Microbiol. 2012, 66, 166–176. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Suppl. M100; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2020. [Google Scholar]
- Fang, Q.; Maglangit, F.; Wu, L.; Ebel, R.; Kyeremeh, K.; Andersen, J.H.; Annang, F.; Pérez-Moreno, G.; Reyes, F.; Deng, H. Signalling and bioactive metabolites from Streptomyces sp. RK44. Molecules 2020, 25, 460. [Google Scholar] [CrossRef] [Green Version]
- Fang, Q.; Maglangit, F.; Mugat, M.; Urwald, C.; Kyeremeh, K.; Deng, H. Targeted isolation of indole alkaloids from Streptomyces sp. CT37. Molecules 2020, 25, 1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Compound Name | MIC (µg/mL) | ||||
|---|---|---|---|---|---|
| S. aureus ATCC 25923) | E. faecalis (ATCC 29212) | E. faecium K59–68 * | E. faecium K60–39 * | S. haemolyticus 8-7A * | |
| Accramycin A 1 | 12.5 | 3.1 | 6.3 | 6.3 | 12.5 |
| Accramycin B 2 | 12.5 | 6.3 | 12.5 | 6.3 | 12.5 |
| Accramycin C 3 | 6.3 | 3.1 | 6.3 | 6.3 | 12.5 |
| Accramycin D 4 | 12.5 | 1.5 | 6.3 | 6.3 | 12.5 |
| Accramycin E 5 | 12.5 | 1.5 | 6.3 | 6.3 | 12.5 |
| Accramycin F 6 | 12.5 | 1.5 | 6.3 | 6.3 | 12.5 |
| Accramycin G 7 | 12.5 | 1.5 | 6.3 | 6.3 | 12.5 |
| Accramycin H 8 | 6.3 | 1.5 | 6.3 | 6.3 | 12.5 |
| Accramycin I 9 | 6.3 | 1.5 | 3.1 | 6.3 | 12.5 |
| Accramycin J 10 | 3.1 | 1.5 | 1.5 | 3.1 | 6.3 |
| Accramycin K 11 | 6.3 | 3.1 | 6.3 | 6.3 | 12.5 |
| Naphthacemycin B1 12 | 3.1 | 6.3 | 6.3 | 3.1 | 12.5 |
| Fasamycin C 13 | 6.3 | 6.3 | 6.3 | 3.1 | 12.5 |
| Ampicillin | 0.5 | 1.0 | 1.5 | 25 | 12.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maglangit, F.; Zhang, Y.; Kyeremeh, K.; Deng, H. Discovery of New Antibacterial Accramycins from a Genetic Variant of the Soil Bacterium, Streptomyces sp. MA37. Biomolecules 2020, 10, 1464. https://doi.org/10.3390/biom10101464
Maglangit F, Zhang Y, Kyeremeh K, Deng H. Discovery of New Antibacterial Accramycins from a Genetic Variant of the Soil Bacterium, Streptomyces sp. MA37. Biomolecules. 2020; 10(10):1464. https://doi.org/10.3390/biom10101464
Chicago/Turabian StyleMaglangit, Fleurdeliz, Yuting Zhang, Kwaku Kyeremeh, and Hai Deng. 2020. "Discovery of New Antibacterial Accramycins from a Genetic Variant of the Soil Bacterium, Streptomyces sp. MA37" Biomolecules 10, no. 10: 1464. https://doi.org/10.3390/biom10101464