Retinoic Acid: A Key Regulator of Lung Development

 ,
,
Abstract
:1. General Introduction
2. Retinoic Acid Signaling Pathway Overview
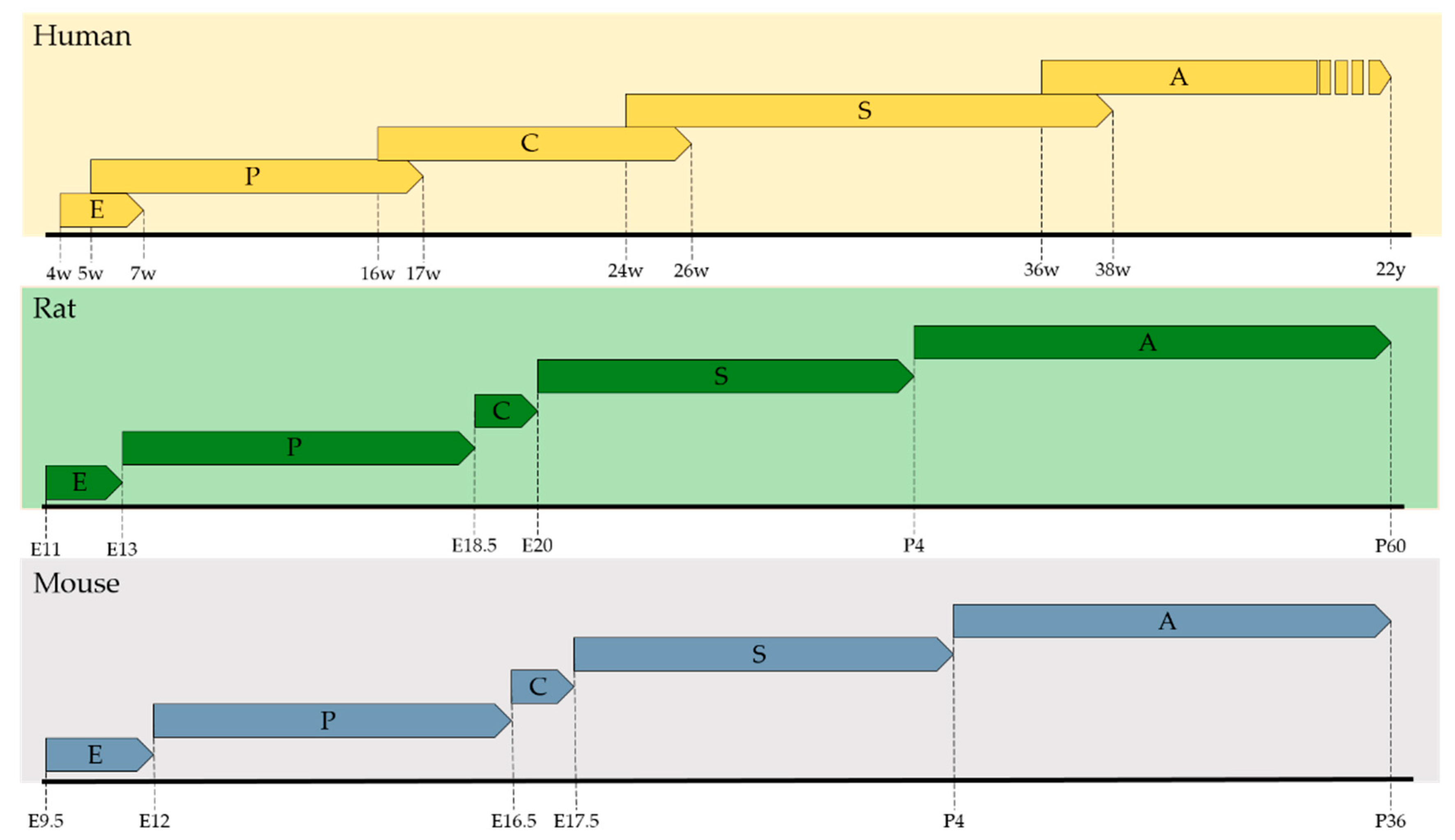
3. Lung Development and Retinoic Acid
3.1. Embryonic Phase
3.2. Pseudoglandular Phase
3.3. Canalicular Phase
3.4. Saccular Phase
3.5. Alveolar Phase
3.6. Vascular Development
4. RA and Lung Disease
5. Final Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Cunningham, T.J.; Duester, G. Mechanisms of retinoic acid signalling and its roles in organ and limb development. Nat. Rev. Mol. Cell Biol. 2015, 16, 110–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghyselinck, N.B.; Duester, G. Retinoic acid signaling pathways. Development 2019, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timoneda, J.; Rodríguez-Fernández, L.; Zaragozá, R.; Marín, M.P.; Cabezuelo, M.T.; Torres, L.; Viña, J.R.; Barber, T. Vitamin A Deficiency and the Lung. Nutrients 2018, 10, 1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, W.V.; Chen, F. Retinoic Acid in the Developing Lung and Other Foregut Derivatives. In The Retinoids: Biology, Biochemistry, and Disease; Dollé, P., Neiderreither, K., Eds.; John Wiley & Sons: IncHoboken, NJ, USA, 2015. [Google Scholar] [CrossRef]
- Quadro, L.; Blaner, W.S.; Salchow, D.J.; Vogel, S.; Piantedosi, R.; Gouras, P.; Freeman, S.; Cosma, M.P.; Colantuoni, V.; Gottesman, M.E. Impaired retinal function and vitamin A availability in mice lacking retinol-binding protein. EMBO J. 1999, 18, 4633–4644. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, R.; Yu, J.; Honda, J.; Hu, J.; Whitelegge, J.; Ping, P.; Wiita, P.; Bok, D.; Sun, H. A membrane receptor for retinol binding protein mediates cellular uptake of vitamin A. Science 2007, 315, 820–825. [Google Scholar] [CrossRef]
- Fex, G.; Johannesson, G. Retinol transfer across and between phospholipid bilayer membranes. Biochim. Biophys. Acta 1988, 944, 249–255. [Google Scholar] [CrossRef]
- Napoli, J.L. Cellular retinoid binding-proteins, CRBP, CRABP, FABP5: Effects on retinoid metabolism, function and related diseases. Pharm. Ther. 2017, 173, 19–33. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.K.; Wassef, L.; Hamberger, L.; Piantedosi, R.; Palczewski, K.; Blaner, W.S.; Quadro, L. Retinyl ester formation by lecithin: Retinol acyltransferase is a key regulator of retinoid homeostasis in mouse embryogenesis. J. Biol. Chem. 2008, 283, 5611–5621. [Google Scholar] [CrossRef] [Green Version]
- Sandell, L.L.; Sanderson, B.W.; Moiseyev, G.; Johnson, T.; Mushegian, A.; Young, K.; Rey, J.P.; Ma, J.X.; Staehling-Hampton, K.; Trainor, P.A. RDH10 is essential for synthesis of embryonic retinoic acid and is required for limb, craniofacial, and organ development. Genes Dev. 2007, 21, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Shannon, S.R.; Moise, A.R.; Trainor, P.A. New insights and changing paradigms in the regulation of vitamin A metabolism in development. Wiley Interdiscip. Rev. Dev. Biol. 2017, 6. [Google Scholar] [CrossRef] [Green Version]
- Napoli, J.L. Physiological insights into all-trans-retinoic acid biosynthesis. Biochim. Biophys Acta 2012, 1821, 152–167. [Google Scholar] [CrossRef] [Green Version]
- Feng, L.; Hernandez, R.E.; Waxman, J.S.; Yelon, D.; Moens, C.B. Dhrs3a regulates retinoic acid biosynthesis through a feedback inhibition mechanism. Dev. Biol. 2010, 338, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Duester, G. Retinoic acid synthesis and signaling during early organogenesis. Cell 2008, 134, 921–931. [Google Scholar] [CrossRef] [Green Version]
- Rhinn, M.; Dollé, P. Retinoic acid signalling during development. Development 2012, 139, 843–858. [Google Scholar] [CrossRef] [Green Version]
- Niederreither, K.; Fraulob, V.; Garnier, J.M.; Chambon, P.; Dollé, P. Differential expression of retinoic acid-synthesizing (RALDH) enzymes during fetal development and organ differentiation in the mouse. Mech. Dev. 2002, 110, 165–171. [Google Scholar] [CrossRef]
- Ross, A.C.; Zolfaghari, R. Cytochrome P450s in the regulation of cellular retinoic acid metabolism. Annu. Rev. Nutr. 2011, 31, 65–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, D.E. Cellular transport and metabolism of vitamin A: Roles of the cellular retinoid-binding proteins. Nutr. Rev. 1994, 5, S24–S31. [Google Scholar] [CrossRef] [PubMed]
- Chambon, P. A decade of molecular biology of retinoic acid receptors. FASEB J. 1996, 10, 940–954. [Google Scholar] [CrossRef]
- Metzler, M.A.; Sandell, L.L. Enzymatic Metabolism of Vitamin A in Developing Vertebrate Embryos. Nutrients 2016, 8, 812. [Google Scholar] [CrossRef]
- Quanjer, P.H.; Stanojevic, S.; Cole, T.J.; Baur, X.; Hall, G.L.; Culver, B.H.; Enright, P.L.; Hankinson, J.L.; Ip, M.S.M.; Zheng, J.; et al. Multi-ethnic reference values for spirometry for the 3-95-yr age range: The global lung function 2012 equations. Eur. Respir. J. 2012, 40, 1324–1343. [Google Scholar] [CrossRef]
- Stocks, J.; Hislop, A.; Sonnappa, S. Early lung development: Lifelong effect on respiratory health and disease. Lancet Respir. Med. 2013, 1, 728–742. [Google Scholar] [CrossRef]
- Burri, P.H. Structural Aspects of Postnatal Lung Development–Alveolar Formation and Growth. Biol. Neonate 2006, 89, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Hogan, B.L.M.; Barkauskas, C.E.; Chapman, H.A.; Epstein, J.A.; Jain, R.; Hsia, C.C.W.; Niklason, L.; Calle, E.; Le, A.; Randell, S.H.; et al. Repair and regeneration of the respiratory system: Complexity, plasticity, and mechanisms of lung stem cell function. Cell Stem Cell 2014, 15, 123–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrisey, E.E.; Hogan, B.L.M. Preparing for the First Breath: Genetic and Cellular Mechanisms in Lung Development. Dev. Cell 2010, 1, 8–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCulley, D.; Wienhold, M.; Sun, X. The pulmonary mesenchyme directs lung development. Curr. Opin. Genet. Dev. 2015, 32, 98–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warburton, D.; Perin, L.; Defilippo, R.; Bellusci, S.; Shi, W.; Driscoll, B. Stem/progenitor cells in lung development, injury repair, and regeneration. Proc. Am. Thorac. Soc. 2008, 5, 703–706. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.Y.; Pang, M.F.; Varner, V.D.; Kojima, L.; Miller, E.; Radisky, D.C.; Nelson, C.M. Localized Smooth Muscle Differentiation Is Essential for Epithelial Bifurcation during Branching Morphogenesis of the Mammalian Lung. Dev. Cell 2015, 34, 719–726. [Google Scholar] [CrossRef] [Green Version]
- Warburton, D.; El-Hashash, A.; Carraro, G.; Tiozzo, C.; Sala, F.; Rogers, O.; De Langhe, S.; Kemp, P.J.; Riccardi, D.; Torday, J.; et al. Chapter Three-Lung Organogenesis. In Current Topics in Developmental Biology; Koopman, P., Ed.; Academic Press: Cambridge, MA, USA, 2010; Volume 90, pp. 73–158. [Google Scholar] [CrossRef]
- Schittny, J.C. Development of the lung. Cell Tissue Res. 2017, 367, 427–444. [Google Scholar] [CrossRef] [Green Version]
- Herriges, M.; Morrisey, E.E. Lung development: Orchestrating the generation and regeneration of a complex organ. Development 2014, 141, 502–513. [Google Scholar] [CrossRef] [Green Version]
- Fernandes-Silva, H.; Correia-Pinto, J.; Moura, R.S. Canonical Sonic Hedgehog Signaling in Early Lung Development. J. Dev. Biol. 2017, 5, 3. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.; Xu, C.; Lu, M.; Wu, X.; Tang, L.; Wu, X. Wnt/β-catenin signaling links embryonic lung development and asthmatic airway remodeling. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 3226–3242. [Google Scholar] [CrossRef] [PubMed]
- Ning, J.; Zhao, Y.; Ye, Y.; Yu, J. Opposing roles and potential antagonistic mechanism between TGF-β and BMP pathways: Implications for cancer progression. EBioMedicine 2019, 41, 702–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danopoulos, S.; Shiosaki, J.; Al Alam, D. FGF Signaling in Lung Development and Disease: Human Versus Mouse. Front. Genet. 2019, 10, 170. [Google Scholar] [CrossRef] [PubMed]
- Yeung, B.; Yu, J.; Yang, X. Roles of the Hippo pathway in lung development and tumorigenesis. Int. J. Cancer 2016, 138, 533–539. [Google Scholar] [CrossRef]
- Lazzaro, D.; Price, M.; Felice, M.D.E.; Lauro, R.D.I. The transcription factor TTF-1 is expressed at the onset of thyroid and lung morphogenesis and in restricted regions of the foetal brain. Development 1991, 113, 1093–1104. [Google Scholar]
- Goss, A.M.; Tian, Y.; Tsukiyama, T.; Cohen, E.D.; Zhou, D.; Lu, M.M.; Yamaguchi, T.P.; Morrisey, E.E. Wnt2/2b and β-Catenin Signaling Are Necessary and Sufficient to Specify Lung Progenitors in the Foregut. Dev. Cell 2009, 17, 290–298. [Google Scholar] [CrossRef] [Green Version]
- Domyan, E.T.; Ferretti, E.; Throckmorton, K.; Mishina, Y.; Nicolis, S.K.; Sun, X. Signaling through BMP receptors promotes respiratory identity in the foregut via repression of Sox2. Development 2011, 138, 971–981. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Desai, T.J.; Qian, J.; Niederreither, K.; Lü, J.; Cardoso, W.V. Inhibition of Tgfβ signaling by endogenous retinoic acid is essential for primary lung bud induction. Development 2007, 134, 2969–2979. [Google Scholar] [CrossRef] [Green Version]
- Malpel, S.; Mendelsohn, C.; Cardoso, W.V. Regulation of retinoic acid signaling during lung morphogenesis. Development 2000, 127, 3057–3067. [Google Scholar]
- Desai, T.J.; Malpel, S.; Flentke, G.R.; Smith, S.M.; Cardoso, W.V. Retinoic acid selectively regulates Fgf10 expression and maintains cell identity in the prospective lung field of the developing foregut. Dev. Biol. 2004, 273, 402–415. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Dollé, P.; Cardoso, W.V.; Niederreither, K. Retinoic acid regulates morphogenesis and patterning of posterior foregut derivatives. Dev. Biol. 2006, 297, 433–445. [Google Scholar] [CrossRef] [PubMed]
- Desai, T.J.; Chen, F.; Lü, J.; Qian, J.; Niederreither, K.; Dollé, P.; Chambon, P.; Cardoso, W.V. Distinct roles for retinoic acid receptors alpha and beta in early lung morphogenesis. Dev. Biol. 2006, 291, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Cao, Y.; Qian, J.; Shao, F.; Niederreither, K.; Cardoso, W.V. A retinoic acid–dependent network in the foregut controls formation of the mouse lung primordium. J. Clin. Investig. 2010, 120, 2040–2048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rankin, S.A.; Han, L.; Kyle, W.; Wells, J.M.; John, M.; Zorn, A.M.; Rankin, S.A.; Han, L.; Mccracken, K.W.; Kenny, A.P.; et al. Article A Retinoic Acid-Hedgehog Cascade Coordinates Mesoderm-Inducing Signals and Endoderm Competence during Lung Specification. Cell Rep. 2016, 16, 66–78. [Google Scholar] [CrossRef] [Green Version]
- Rankin, S.A.; McCracken, K.W.; Luedeke, D.M.; Han, L.; Wells, J.M.; Shannon, J.M.; Zorn, A.M. Timing is everything: Reiterative Wnt, BMP and RA signaling regulate developmental competence during endoderm organogenesis. Dev. Biol. 2018, 434, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Metzger, R.J.; Klein, O.D.; Martin, G.R.; Krasnow, M.A. The branching programme of mouse lung development. Nature 2008, 453, 745–750. [Google Scholar] [CrossRef] [Green Version]
- Chazaud, C.; Dollé, P.; Rossant, J.; Mollard, R. Retinoic acid signaling regulates murine bronchial tubule formation. Mech. Dev. 2003, 120, 691–700. [Google Scholar] [CrossRef]
- Fernandes-Silva, H.; Vaz-Cunha, P.; Barbosa, V.B.; Silva-Gonçalves, C.; Correia-Pinto, J.; Moura, R.S. Retinoic acid regulates avian lung branching through a molecular network. Cell Mol. Life Sci. 2017, 74, 4599–4619. [Google Scholar] [CrossRef] [Green Version]
- Rhinn, M.; Schuhbaur, B.; Niederreither, K.; Dollé, P. Involvement of retinol dehydrogenase 10 in embryonic patterning and rescue of its loss of function by maternal retinaldehyde treatment. Proc. Natl. Acad. Sci. USA 2011, 108, 16687–16692. [Google Scholar] [CrossRef] [Green Version]
- Bouillet, P.; Sapin, V.; Chazaud, C.; Messaddeq, N.; Décimo, D.; Dollé, P.; Chambon, P. Developmental expression pattern of Stra6, a retinoic acid-responsive gene encoding a new type of membrane protein. Mech. Dev. 1997, 63, 173–186. [Google Scholar] [CrossRef]
- Dollé, P.; Ruberte, E.; Leroy, P.; Morriss-Kay, G.; Chambon, P. Retinoic acid receptors and cellular retinoid binding proteins. I. A systematic study of their differential pattern of transcription during mouse organogenesis. Development 1990, 110, 1133–1151. [Google Scholar] [PubMed]
- Bellusci, S.; Grindley, J.; Emoto, H.; Itoh, N.; Hogan, B.L. Fibroblast growth factor 10 (FGF10) and branching morphogenesis in the embryonic mouse lung. Development 1997, 124, 4867–4878. [Google Scholar] [PubMed]
- Mendelsohn, C.; Lohnes, D.; Décimo, D.; Lufkin, T.; LeMeur, M.; Chambon, P.; Mark, M. Function of the retinoic acid receptors (RARs) during development (II). Multiple abnormalities at various stages of organogenesis in RAR double mutants. Development 1994, 120, 2749–2771. [Google Scholar] [PubMed]
- Mollard, R.; Ghyselinck, N.B.; Wendling, O.; Chambon, P.; Mark, M. Stage-dependent responses of the developing lung to retinoic acid signaling. Int. J. Dev. Biol. 2000, 44, 457–462. [Google Scholar]
- Lohnes, D.; Mark, M.; Mendelsohn, C.; Dollé, P.; Dierich, A.; Gorry, P.; Gansmuller, A.; Chambon, P. Function of the retinoic acid receptors (RARs) during development (I). Craniofacial and skeletal abnormalities in RAR double mutants. Development 1994, 120, 2723–2748. [Google Scholar]
- Lü, J.; Qian, J.; Izvolsky, K.I.; Cardoso, W.V. Global analysis of genes differentially expressed in branching and non-branching regions of the mouse embryonic lung. Dev. Biol. 2004, 273, 418–435. [Google Scholar] [CrossRef] [Green Version]
- Moura, R.S.; Coutinho-Borges, J.P.; Pacheco, A.P.; Damota, P.O.; Correia-Pinto, J. FGF signaling pathway in the developing chick lung: Expression and inhibition studies. PLoS ONE 2011, 6, e17660. [Google Scholar] [CrossRef] [Green Version]
- Schuger, L.; Varani, J.; Mitra, R., Jr.; Gilbride, K. Retinoic acid stimulates mouse lung development by a mechanism involving epithelial-mesenchymal interaction and regulation of epidermal growth factor receptors. Dev. Biol. 1993, 159, 462–473. [Google Scholar] [CrossRef]
- Montedonico, S.; Nakazawa, N.; Puri, P. Retinoic acid rescues lung hypoplasia in nitrofen-induced hypoplastic foetal rat lung explants. Pediatr. Surg. Int. 2006, 22, 2–8. [Google Scholar] [CrossRef]
- Pereira-Terra, P.; Moura, R.S.; Nogueira-Silva, C.; Correia-Pinto, J. Neuroendocrine factors regulate retinoic acid receptors in normal and hypoplastic lung development. J. Physiol. 2015, 593, 3301–3311. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, W.V.; Williams, M.C.; Mitsialis, S.A.; Joyce-Brady, M.; Rishi, A.K.; Brody, J.S. Retinoic acid induces changes in the pattern of airway branching and alters epithelial cell differentiation in the developing lung in vitro. Am. J. Respir. Cell Mol. Biol. 1995, 12, 464–476. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, W.V.; Mitsialis, S.A.; Brody, J.S.; Williams, M.C. Retinoic acid alters the expression of pattern-related genes in the developing rat lung. Dev. Dyn. 1996, 207, 47–59. [Google Scholar] [CrossRef]
- Urase, K.; Mukasa, T.; Igarashi, H.; Ishii, Y.; Yasugi, S.; Momoi, M.Y.; Momoi, T. Spatial expression of Sonic hedgehog in the lung epithelium during branching morphogenesis. Biochem. Biophys. Res. Commun. 1996, 225, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Lebeche, D.; Malpel, S.; Cardoso, W.V. Fibroblast growth factor interactions in the developing lung. Mech. Dev. 1999, 86, 125–136. [Google Scholar] [CrossRef]
- Weaver, M.; Yingling, J.M.; Dunn, N.R.; Bellusci, S.; Hogan, B.L. Bmp signaling regulates proximal-distal differentiation of endoderm in mouse lung development. Development 1999, 126, 4005–4015. [Google Scholar]
- Zhou, L.; Dey, C.R.; Wert, S.E.; Yan, C.; Costa, R.H.; Whitsett, J.A. Hepatocyte nuclear factor-3beta limits cellular diversity in the developing respiratory epithelium and alters lung morphogenesis in vivo. Dev. Dyn. 1997, 210, 305–314. [Google Scholar] [CrossRef]
- Bogue, C.W.; Gross, I.; Vasavada, H.; Dynia, D.W.; Wilson, C.M.; Jacobs, H.C. Identification of Hox genes in newborn lung and effects of gestational age and retinoic acid on their expression. Am. J. Physiol. 1994, 266, L448–L454. [Google Scholar] [CrossRef]
- Volpe, M.V.; Vosatka, R.J.; Nielsen, H.C. Hoxb-5 control of early airway formation during branching morphogenesis in the developing mouse lung. Biochim. Biophys. Acta 2000, 1475, 337–345. [Google Scholar] [CrossRef]
- Chen, F.; Marquez, H.; Kim, Y.K.; Qian, J.; Shao, F.; Fine, A.; Cruikshank, W.W.; Quadro, L.; Cardoso, W.V. Prenatal retinoid deficiency leads to airway hyperresponsiveness in adult mice. J. Clin. Investig. 2014, 124, 801–811. [Google Scholar] [CrossRef] [Green Version]
- Gontan, C.; de Munck, A.; Vermeij, M.; Grosveld, F.; Tibboel, D.; Rottier, R. Sox2 is important for two crucial processes in lung development: Branching morphogenesis and epithelial cell differentiation. Dev. Biol. 2008, 317, 296–309. [Google Scholar] [CrossRef] [Green Version]
- Rockich, B.; Hrycaj, S.M.; Shih, H.P.; Nagy, M.S.; Ferguson, M.A.; Kopp, J.L.; Sander, M.; Wellik, D.M.; Spence, J.R. Sox9 plays multiple roles in the lung epithelium during branching morphogenesis. Proc. Natl. Acad. Sci. USA 2013, 110, E4456–E4464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- deMello, D.E.; Reid, L.M. Embryonic and early fetal development of human lung vasculature and its functional implications. Pediatr. Dev. Pathol. 2000, 3, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Ornitz, D.M.; Yin, Y. Signaling networks regulating development of the lower respiratory tract. Cold Spring Harb. Perspect. Biol. 2012, 4. [Google Scholar] [CrossRef] [PubMed]
- Boyden, E.A. The structure of the pulmonary acinus in a child of six years and eight months. Am. J. Anat. 1971, 132, 275–299. [Google Scholar] [CrossRef]
- Kitaoka, H.; Burri, P.H.; Weibel, E.R. Development of the human fetal airway tree: Analysis of the numerical density of airway endtips. Anat. Rec. 1996, 244, 207–213. [Google Scholar] [CrossRef]
- DiMaio, M.; Gil, J.; Ciurea, D.; Kattan, M. Structural maturation of the human fetal lung: A morphometric study of the development of air-blood barriers. Pediatr. Res. 1989, 26, 88–93. [Google Scholar] [CrossRef] [Green Version]
- McQualter, J.L.; Yuen, K.; Williams, B.; Bertoncello, I. Evidence of an epithelial stem/progenitor cell hierarchy in the adult mouse lung. Proc. Natl. Acad. Sci. USA 2010, 107, 1414–1419. [Google Scholar] [CrossRef] [Green Version]
- Nolen-Walston, R.D.; Kim, C.F.; Mazan, M.R.; Ingenito, E.P.; Gruntman, A.M.; Tsai, L.; Boston, R.; Woolfenden, A.E.; Jacks, T.; Hoffman, A.M. Cellular kinetics and modeling of bronchioalveolar stem cell response during lung regeneration. Am. J. Physiol. Lung Cell Mol. Physiol. 2008, 294, 1158–1166. [Google Scholar] [CrossRef] [Green Version]
- Barré, S.F.; Haberthür, D.; Cremona, T.P.; Stampanoni, M.; Schittny, J.C. The total number of acini remains constant throughout postnatal rat lung development. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 311, L1082–L1089. [Google Scholar] [CrossRef] [Green Version]
- DeMello, D.E.; Sawyer, D.; Galvin, N.; Reid, L.M. Early Fetal Development of Lung Vasculature. Am. J. Respir. Cell Mol. Biol. 1997, 16, 568–581. [Google Scholar] [CrossRef]
- Vu, T.H.; Alemayehu, Y.; Werb, Z. New insights into saccular development and vascular formation in lung allografts under the renal capsule. Mech. Dev. 2003, 120, 305–313. [Google Scholar] [CrossRef]
- Grummer, M.A.; Thet, L.A.; Zachman, R.D. Expression of retinoic acid receptor genes in fetal and newborn rat lung. Pediatr. Pulmonol. 1994, 17, 234–238. [Google Scholar] [CrossRef]
- Pawlikowski, B.; Wragge, J.; Siegenthaler, J.A. Retinoic acid signaling in vascular development. Genesis 2019, 57, e23287. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.H.; Lakshminrusimha, S.; El, Abiad, M.T.; Chess, P.R.; Ryan, R.M. Growth factors in lung development. Adv. Clin. Chem. 2005, 40, 261–316. [Google Scholar] [CrossRef] [PubMed]
- Nikolić, M.Z.; Sun, D.; Rawlins, E.L. Human lung development: Recent progress and new challenges. Development 2018, 145. [Google Scholar] [CrossRef] [Green Version]
- Wongtrakool, C.; Malpel, S.; Gorenstein, J.; Sedita, J.; Ramirez, M.I.; Underhill, T.M.; Cardoso, W.V. Down-regulation of retinoic acid receptor alpha signaling is required for sacculation and type I cell formation in the developing lung. J. Biol. Chem. 2003, 278, 46911–46928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massaro, G.D. Formation of Pulmonary Alveoli and Gas-Exchange Surface Area: Quantitation and Regulation. Annu. Rev. Physiol. 1996, 58, 73–92. [Google Scholar] [CrossRef]
- Boström, H.; Willetts, K.; Pekny, M.; Levéen, P.; Lindahl, P.; Hedstrand, H.; Pekna, M.; Hellström, M.; Gebre-Medhin, S.; Schalling, M.; et al. PDGF-A signaling is a critical event in lung alveolar myofibroblast development and alveogenesis. Cell 1996, 85, 863–873. [Google Scholar] [CrossRef] [Green Version]
- Lindahl, P.; Karlsson, L.; Hellström, M.; Gebre-Medhin, S.; Willetts, K.; Heath, J.K.; Betsholtz, C. Alveogenesis failure in PDGF-A-deficient mice is coupled to lack of distal spreading of alveolar smooth muscle cell progenitors during lung development. Development 1997, 124, 3943–3953. [Google Scholar]
- Burri, P.H. Postnatal growth and maturation of the lung. Chest 1975, 67, 2S–3S. [Google Scholar] [CrossRef]
- Caduff, J.H.; Fischer, L.C.; Burri, P.H. Scanning electron microscope study of the developing microvasculature in the postnatal rat lung. Anat. Rec. 1986, 216, 154–164. [Google Scholar] [CrossRef] [PubMed]
- McGowan, S.; Jackson, S.K.; Jenkins-Moore, M.; Dai, H.H.; Chambon, P.; Snyder, J.M. Mice bearing deletions of retinoic acid receptors demonstrate reduced lung elastin and alveolar numbers. Am. J. Respir. Cell Mol. Biol. 2000, 23, 162–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinstein, M.; Xu, X.; Ohyama, K.; Deng, C.X. FGFR-3 and FGFR-4 function cooperatively to direct alveogenesis in the murine lung. Development 1998, 125, 3615–3623. [Google Scholar] [PubMed]
- Massaro, G.D.; Massaro, D. Treatment with retinoic acid increases the number of pulmonary alveoli. FASEB J. 1996, 270, L305–L310. [Google Scholar] [CrossRef]
- Hind., M.; Corcoran., J.; Maden., M. Temporal/spatial expression of retinoid binding proteins and RAR isoforms in the postnatal lung. Am. J. Physiol. Lung Cell Mol. Physiol. 2002, 282, L468–L476. [Google Scholar] [CrossRef]
- Geevarghese, S.K.; Chytil, F. Depletion of retinyl esters in the lungs coincides with lung prenatal morphological maturation. Biochem. Biophys. Res. Commun. 1994, 200, 529–535. [Google Scholar] [CrossRef]
- McGowan, S.E. Contributions of retinoids to the generation and repair of the pulmonary alveolus. Chest 2002, 121, 206S–208S. [Google Scholar] [CrossRef] [Green Version]
- Baybutt, R.C.; Hu, L.; Molteni, A. Vitamin A Deficiency Injures Lung and Liver Parenchyma and Impairs Function of Rat Type II Pneumocytes. J. Nutr. 2000, 130, 1159–1165. [Google Scholar] [CrossRef]
- Biesalski, H.K.; Nohr, D. Importance of vitamin-A for lung function and development. Mol. Asp. Med. 2003, 24, 431–440. [Google Scholar] [CrossRef]
- Verma, R.P.; Mcculloch, K.M.; Worrell, L.; Vidyasagar, D. Vitamin A deficiency and severe bronchopulmonary dysplasia in very low birthweight infants. Am. J. Perinatol. 1996, 13, 389–393. [Google Scholar] [CrossRef]
- Albertine, K.H.; Jones, G.P.; Starcher, B.C.; Bohnsack, J.F.; Davis, P.L.; Cho, S.C.; Carlton, D.P.; Bland, R.D. Chronic lung injury in preterm lambs: Disordered respiratory tract development. Am. J. Respir. Crit. Care Med. 1999, 159, 945–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGowan, S.E.; Harvey, C.S.; Jackson, S.K. Retinoids, retinoic acid receptors, and cytoplasmic retinoid binding proteins in perinatal rat lung fibroblasts. Am. J. Physiol. Lung Cell Mol. Physiol. 1995, 269, L463–L472. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.T.; Whitney, P.L. Postnatal development of rat lung. Changes in lung lectin, elastin, acetylcholinesterase and other enzymes. Biochem. J. 1980, 188, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Ong, D.E.; Chytil, F. Changes in levels of cellular retinol and retinoic acid binding proteins of liver and lung during perinatal development of rat. Proc. Natl. Acad. Sci. USA 1976, 73, 3976–3978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hind, M.; Corcoran, J.; Maden, M. Alveolar proliferation, retinoid synthesizing enzymes, and endogenous retinoids in the postnatal mouse lung different roles for Aldh-1 and Raldh-2. Am. J. Respir Cell Mol. Biol. 2002, 26, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Yun, E.J.; Lorizio, W.; Seedorf, G.; Abman, S.H.; Vu, T.H. VEGF and endothelium-derived retinoic acid regulate lung vascular and alveolar development. Am. J. Physiol. Lung Cell Mol. Physiol. 2016, 310, L287–L298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massaro, G.D.; Massaro, D.; Chambon, P. Retinoic acid receptor-α regulates pulmonary alveolus formation in mice after, but not during, perinatal period. Am. J. Physiol. Lung Cell Mol. Physiol. 2003, 284, 2002–2004. [Google Scholar] [CrossRef] [Green Version]
- Massaro, G.D.; Massaro, D.; Chan, W.Y.; Clerch, L.B.; Ghyselinck, N.; Chambon, P.; Chandraratna, R.A.S. Retinoic acid receptor-β: An endogenous inhibitor of the perinatal formation of pulmonary alveoli. Physiol. Genom. 2001, 4, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Snyder, J.M.; Jenkins-Moore, M.; Jackson, S.K.; Goss, K.L.; Dai, H.H.; Bangsund, P.J.; Giguere, V.; Mcgowan, S.E. Alveolarization in retinoic acid receptor-β-deficient mice. Pediatr. Res. 2005, 57, 384–391. [Google Scholar] [CrossRef] [Green Version]
- Massaro, G.D.; Massaro, D. Retinoic acid treatment abrogates elastase-induced pulmonary emphysema in rats. Nat. Med. 1997, 3, 675–677. [Google Scholar] [CrossRef]
- Shenai, J.P.; Chytil, F. Effect of maternal vitamin-A administration on fetal lung vitamin-A stores in the perinatal rat. Biol. Neonate 1990, 58, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Okabe, T.; Yorifuji, H.; Yamada, E.; Takaku, F. Isolation and characterization of vitamin-A-storing lung cells. Exp. Cell Res. 1984, 154, 125–135. [Google Scholar] [CrossRef]
- Dirami, G.; Massaro, G.D.; Clerch, L.B.; Ryan, U.S.; Reczek, P.R.; Massaro, D. Lung retinol storing cells synthesize and secrete retinoic acid, an inducer of alveolus formation. Am. J. Physiol. Lung Cell Mol. Physiol. 2018, 286, 249–256. [Google Scholar] [CrossRef] [Green Version]
- McGowan, S.E.; Torday, J.S. The pulmonary lipofibroblast (lipid interstitial cell) and its contributions to alveolar development. Annu. Rev. Physiol. 1997, 59, 43–62. [Google Scholar] [CrossRef] [PubMed]
- Shenai, J.P.; Chytil, F. Vitamin A storage in lungs during perinatal development in the rat. Biol. Neonate 1990, 57, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Burri, P.H. The postnatal growth of the rat lung III. Morphology. Anat. Rec. 1974, 180, 77–98. [Google Scholar] [CrossRef]
- Mcgowan, S.E.; Doro, M.M.; Jackson, S.K. Endogenous retinoids increase perinatal elastin gene expression in rat lung fibroblasts and fetal explants. Am. J. Physiol. Lung Cell Mol. Physiol. 1997, 273, L410–L416. [Google Scholar] [CrossRef]
- Liu, R.; Harvey, C.S.; McGowan, S.E. Retinoic acid increases elastin in neonatal rat lung fibroblast cultures. Am. J. Physiol. Lung Cell Mol. Physiol. 1993, 265, L430–L437. [Google Scholar] [CrossRef]
- Nabeyrat, E.; Besnard, V.; Corroyer, S.; Cazals, V.; Clement, A. Retinoic acid-induced proliferation of lung alveolar epithelial cells: Relation with the IGF system. Am. J. Physiol. Lung Cell Mol. Physiol. 1998, 275, 71–79. [Google Scholar] [CrossRef]
- Nabeyrat, E.; Corroyer, S.; Epaud, R.; Besnard, R.I.E.; Corroyer, S.; Epaud, R. Retinoic acid-induced proliferation of lung alveolar epithelial cells is linked to p21(CIP1) downregulation. Am. J. Physiol. Lung Cell Mol. Physiol. 2000, 278, L42–L50. [Google Scholar] [CrossRef]
- Ng-Blichfeldt, J.P.; Alçada, J.; Montero, M.A.; Dean, C.H.; Griesenbach, U.; Griffiths, M.J.; Hind, M. Deficient retinoid-driven angiogenesis may contribute to failure of adult human lung regeneration in emphysema. Thorax 2017, 72, 510–521. [Google Scholar] [CrossRef] [PubMed]
- Zachman, R.D.; Kakkad, B.; Chytil, F. Perinatal rat lung retinol (vitamin A) and retinyl palmitate. Pediatr. Res. 1984, 18, 1297–1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolbach, S.B.; Howe, P.R. Tissue Changes Following Deprivation of Fat-Soluble A Vitamin. J. Exp. Med. 1925, 42, 753–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraslon, C.; Bourbon, J.R. Retinoids control surfactant phospholipid biosynthesis in fetal rat lung. Am. J. Physiol. Lung Cell Mol. Physiol. 1994, 266, L705–L712. [Google Scholar] [CrossRef] [PubMed]
- Metzler, M.D.; Snyder, J.M. Retinoic acid differentially regulates expression of surfactant-associated proteins in human fetal lung. Endocrinology 1993, 133, 1990–1998. [Google Scholar] [CrossRef]
- Maden, M.; Hind, M. Retinoic acid in alveolar development, maintenance and regeneration. Philos. Trans. R. Soc. B Biol. Sci. 2004, 359, 799–808. [Google Scholar] [CrossRef] [Green Version]
- Hind, M.; Maden, M. Retinoic acid induces alveolar regeneration in the adult mouse lung. Eur. Respir J. 2004, 23, 20–27. [Google Scholar] [CrossRef]
- Maden, M. Retinoids have differing efficacies on alveolar regeneration in a dexamethasone-treated mouse. Am. J. Respir Cell Mol. Biol. 2006, 35, 260–267. [Google Scholar] [CrossRef] [Green Version]
- Belloni, P.N.; Garvin, L.; Mao, C.P.; Bailey-Healy, I.; Leaffer, D. Effects of all-trans-retinoic acid in promoting alveolar repair. Chest 2000, 117, 235S–241S. [Google Scholar] [CrossRef]
- Srinivasan, G.; Bruce, E.N.; Houtz, P.K.; Bruce, M.C. Dexamethasone-induced changes in lung function are not prevented by concomitant treatment with retinoic acid. Am. J. Physiol. Lung Cell Mol. Physiol. 2002, 283, 275–287. [Google Scholar] [CrossRef] [Green Version]
- Ng-Blichfeldt, J.P.; Gosens, R.; Dean, C.; Griffiths, M.; Hind, M. Regenerative pharmacology for COPD: Breathing new life into old lungs. Thorax 2019, 74, 890–897. [Google Scholar] [CrossRef] [PubMed]
- Uniyal, S.; Dhasmana, A.; Tyagi, A.; Muyal, J.P. ATRA reduces inflammation and improves alveolar epithelium regeneration in emphysematous rat lung. Biomed. Pharmacother. 2018, 108, 1435–1450. [Google Scholar] [CrossRef] [PubMed]
- Parera, M.C.; Van Dooren, M.; Van Kempen, M.; De Krijger, R.; Grosveld, F.; Tibboel, D.; Rottier, R. Distal angiogenesis: A new concept for lung vascular morphogenesis. Am. J. Physiol. Lung Cell Mol. Physiol. 2005, 288, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Kool, H.; Mous, D.; Tibboel, D.; de Klein, A.; Rottier, R.J. Pulmonary vascular development goes awry in congenital lung abnormalities. Birth Defects Res. Part. C Embryo Today Rev. 2014, 102, 343–358. [Google Scholar] [CrossRef]
- Yamamoto, H.; Jun Yun, E.; Gerber, H.P.; Ferrara, N.; Whitsett, J.A.; Vu, T.H. Epithelial-vascular cross talk mediated by VEGF-A and HGF signaling directs primary septae formation during distal lung morphogenesis. Dev. Biol. 2007, 308, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Armulik, A.; Abramsson, A.; Betsholtz, C. Endothelial/pericyte interactions. Circ. Res. 2005, 97, 512–523. [Google Scholar] [CrossRef] [Green Version]
- Lai, L.; Bohnsack, B.L.; Niederreither, K.; Hirschi, K.K. Retinoic acid regulates endothelial cell proliferation during vasculogenesis. Development 2003, 130, 6465–6474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, A.F.; Gonçalves, F.L.L.; Regis, A.C.; Gallindo, R.M.; Sbragia, L. Prenatal retinoic acid improves lung vascularization and VEGF expression in CDH rat. Am. J. Obs. Gynecol. 2012, 207, 76. [Google Scholar] [CrossRef]
- Kool, H.M.; Bürgisser, P.E.; Edel, G.G.; de Kleer, I.; Boerema-de Munck, A.; de Laat, I.; Chrifi, I.; Cheng, C.; van Cappellen, W.A.; Kremers, G.J.; et al. Inhibition of retinoic acid signaling induces aberrant pericyte coverage and differentiation resulting in vascular defects in congenital diaphragmatic hernia. Am. J. Physiol. Lung Cell Mol. Physiol. 2019, 317, 317–331. [Google Scholar] [CrossRef]
- Beurskens, N.; Klaassens, M.; Rottier, R.; de Klein, A.; Tibboel, D. Linking animal models to human congenital diaphragmatic hernia. Birth Defects Res. A Clin. Mol. Teratol. 2007, 79, 565–572. [Google Scholar] [CrossRef]
- Kardon, G.; Ackerman, K.G.; McCulley, D.J.; Shen, Y.; Wynn, J.; Shang, L.; Bogenschutz, E.; Sun, X.; Chung, W.K. Congenital diaphragmatic hernias: From genes to mechanisms to therapies. Dis. Model. Mech. 2017, 10, 955–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Major, D.; Cadenas, M.; Fournier, L.; Leclerc, S.; Lefebvre, M.; Cloutier, R. Retinol status of newborn infants with congenital diaphragmatic hernia. Pediatr. Surg. Int. 1998, 13, 547–549. [Google Scholar] [CrossRef] [PubMed]
- Marquez, H.A.; Cardoso, W.V. Vitamin A-retinoid signaling in pulmonary development and disease. Mol. Cell Pediatr. 2016, 3, 28. [Google Scholar] [CrossRef] [Green Version]
- Pierce, R.A.; Joyce, B.; Officer, S.; Heintz, C.; Moore, C.; McCurnin, D.; Johnston, C.; Maniscalco, W. Retinoids increase lung elastin expression but fail to alter morphology or angiogenesis genes in premature ventilated baboons. Pediatr. Res. 2007, 61, 703–709. [Google Scholar] [CrossRef] [Green Version]
- Hind, M.; Gilthorpe, A.; Stinchcombe, S.; Maden, M. Retinoid induction of alveolar regeneration: From mice to man? Thorax 2009, 64, 451–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedmacher, F.; Hofmann, A.D.; Takahashi, T.; Takahashi, H.; Kutasy, B.; Puri, P. Prenatal administration of all-trans retinoic acid upregulates leptin signaling in hypoplastic rat lungs with experimental congenital diaphragmatic hernia. Pediatr. Surg. Int. 2014, 30, 1183–1190. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Lung Developmental Phase | RA Pathway Machinery | RA Pathway Targets | Knockouts with Lung Phenotype | References |
|---|---|---|---|---|
| Embryonic | cyp26 raldh2 RARα, β, γ | fgf10, FGFR2 WNT, wnt2/2b TGFβ HH bmp4 nkx2.1 | raldh2−/− (lung agenesis) | [40,41,42,43,44,45,46,47] |
| Pseudoglandular | stra6 crbp raldh1, 2, 3 cyp26 rarα1, α2, β, γ coup-tfII | fgf10, fgfr2 shh bmp4 foxa2 tgfβ2, β3 cftr hoxa2, a5, b5, b6 HOXB5 acta myh11 sox2, sox9, id2 meis2 crbpI raldh1 rarβ, RARα, β, γ | rdh10−/− (absence of lung, primary lung bud growth arrested and branching impaired) raldh2−/− (hypoplastic lungs, defective growth and branching) rarαβ2−/− (lung agenesis and hypoplasia, absence of lung budding, altered branching) | [16,41,43,49,50,51,52,53,54,55,56,57,58,62,64,69,70,71] |
| Canalicular | RARα | - | - | [84] |
| Saccular | rarα, β, γ | - | - | [84,88] |
| Alveolar | CRBPI, II CRABP RALDH1, 2 RARα, β, γ RXR | VEGFA FGF signaling SPA, B, C, spa YAP TNF-α, TNFR1 IL6, IL6ST | rarα−/− (↓ alveolar number and surface area) rarβ−/− (early & faster septation; ↓ surface area) rarγ−/− (↓ elastic tissue, ↓ alveoli number, ↑ distance between the alveolar walls) rxrα−/− (embryonic lethality) rarγ−/−, rxrα+/− (↓ elastic tissue at the alveolar septa) | [88,94,97,104,107,108,109,110,111,112,123,126,127,130,134] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes-Silva, H.; Araújo-Silva, H.; Correia-Pinto, J.; Moura, R.S. Retinoic Acid: A Key Regulator of Lung Development. Biomolecules 2020, 10, 152. https://doi.org/10.3390/biom10010152
Fernandes-Silva H, Araújo-Silva H, Correia-Pinto J, Moura RS. Retinoic Acid: A Key Regulator of Lung Development. Biomolecules. 2020; 10(1):152. https://doi.org/10.3390/biom10010152
Chicago/Turabian StyleFernandes-Silva, Hugo, Henrique Araújo-Silva, Jorge Correia-Pinto, and Rute S Moura. 2020. "Retinoic Acid: A Key Regulator of Lung Development" Biomolecules 10, no. 1: 152. https://doi.org/10.3390/biom10010152