
Escherichia coli (Lilly) and Saccharomyces cerevisiae (Novo) rDNA Glucagon: An Assessment of Their Actions When Supplied Selectively to Periportal Cells in the Bivascularly Perfused Rat Liver


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
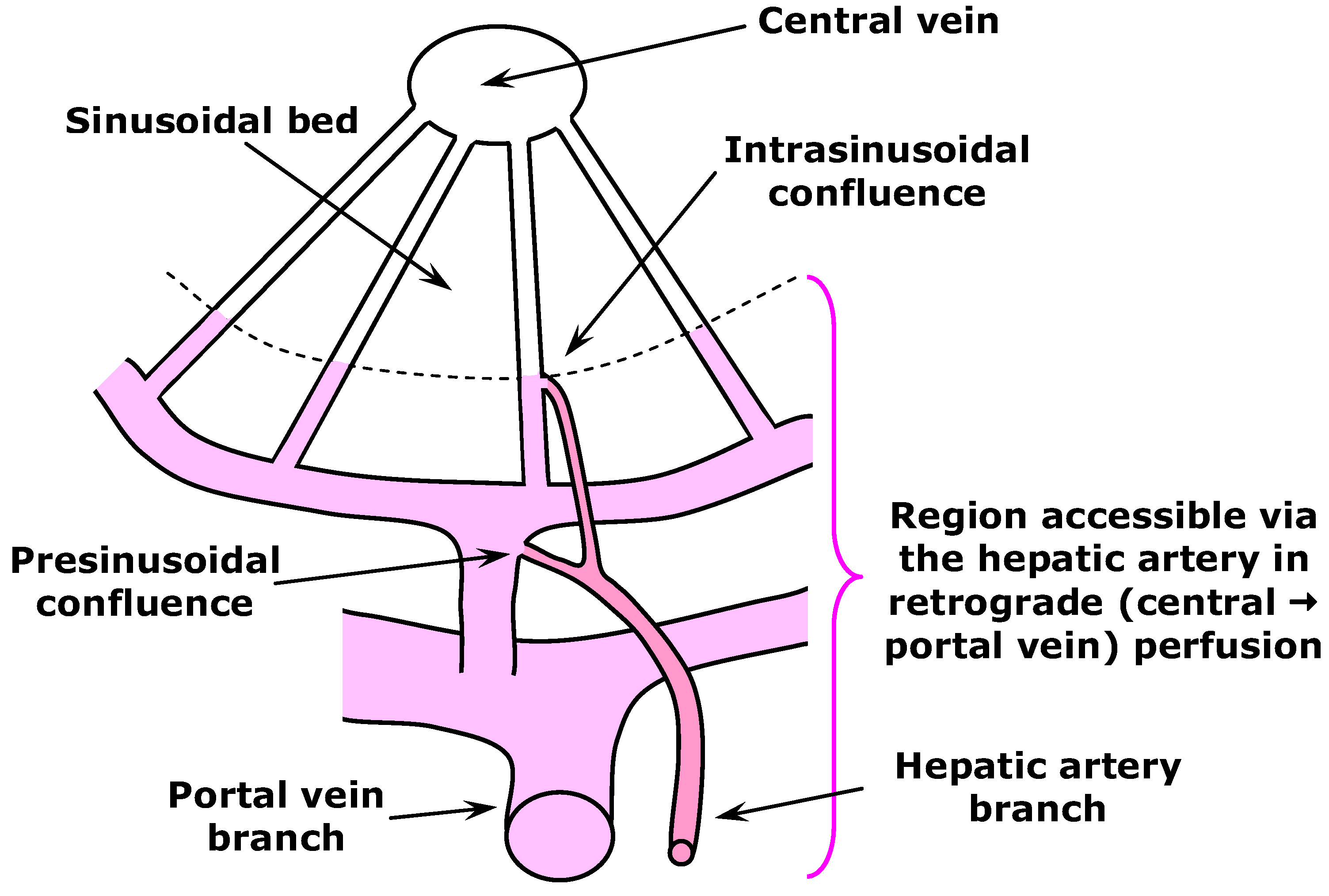
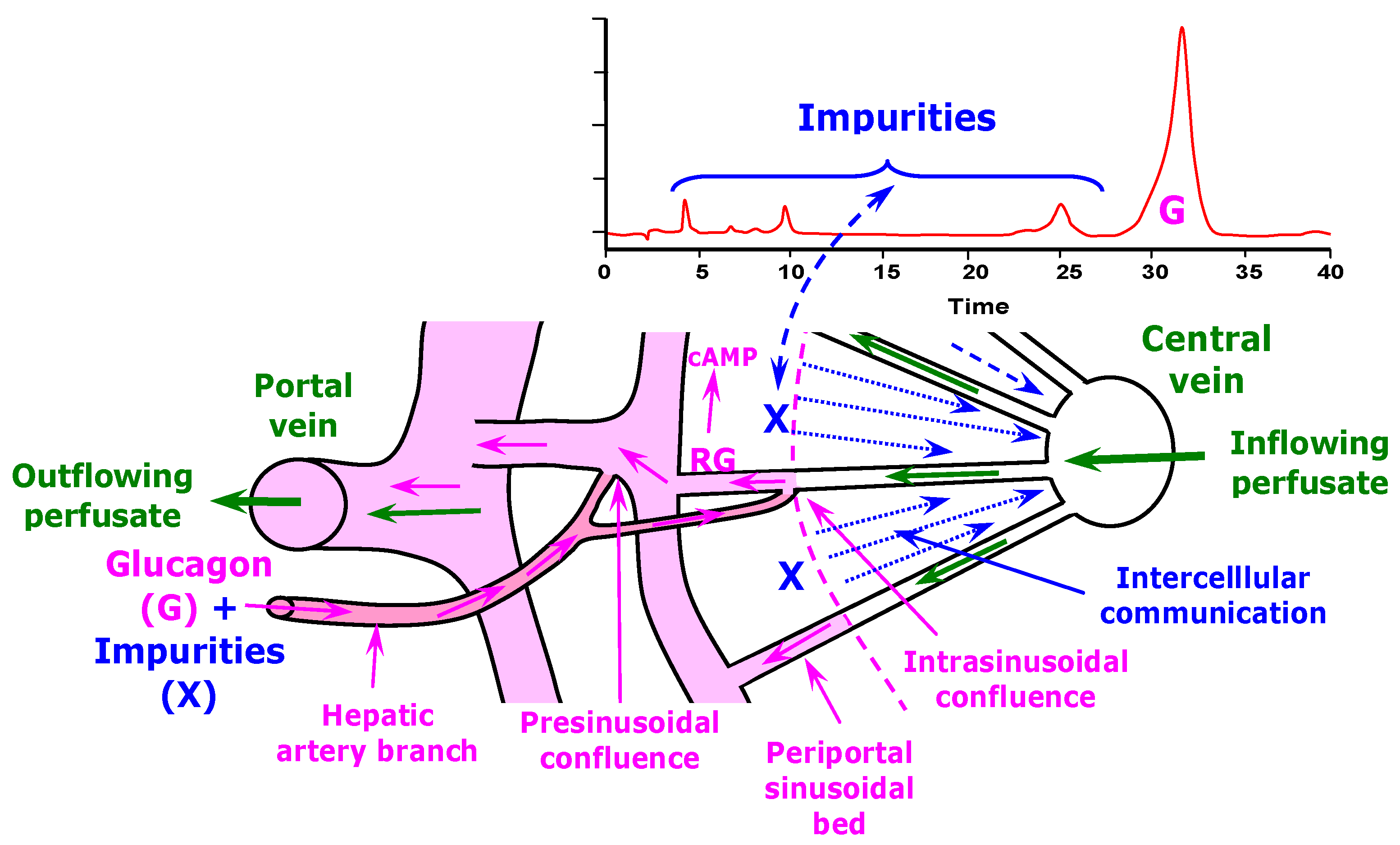
2.2. Liver Perfusion
2.3. Analytical
2.4. Chromatographic Analyses
2.5. Treatment of Data
3. Results
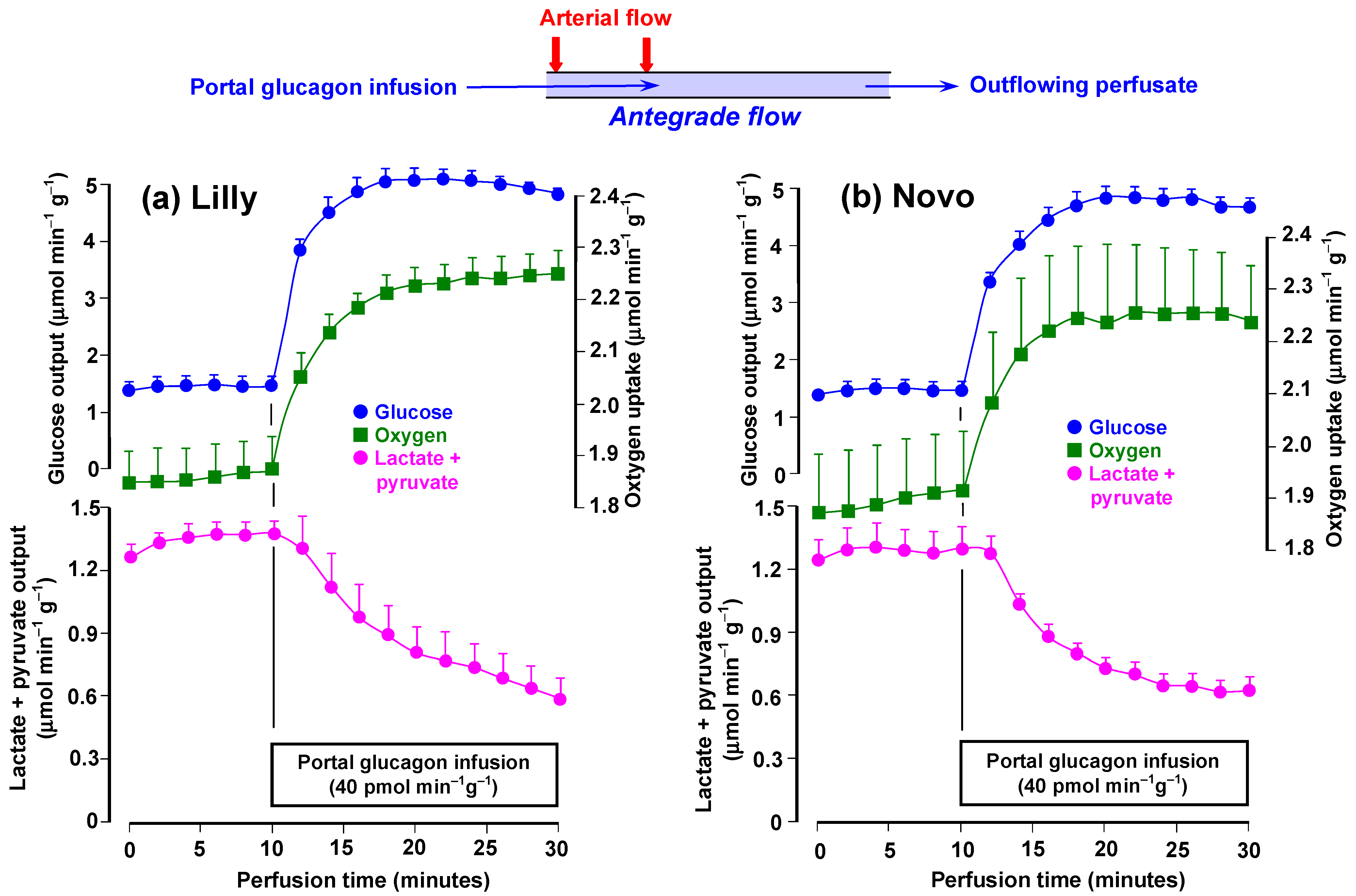
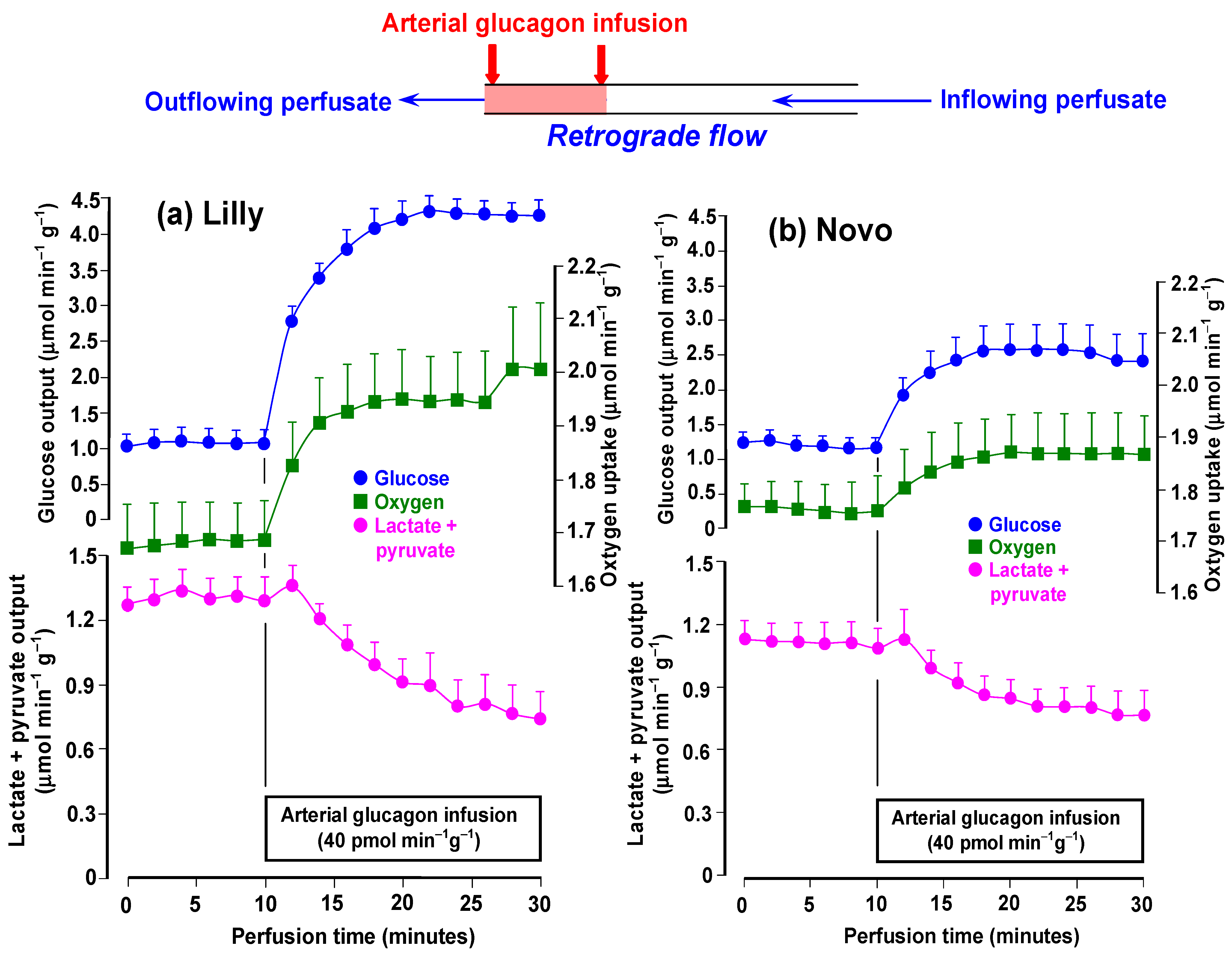
3.1. Time Courses of the Effects of rDNA Glucagon Preparations
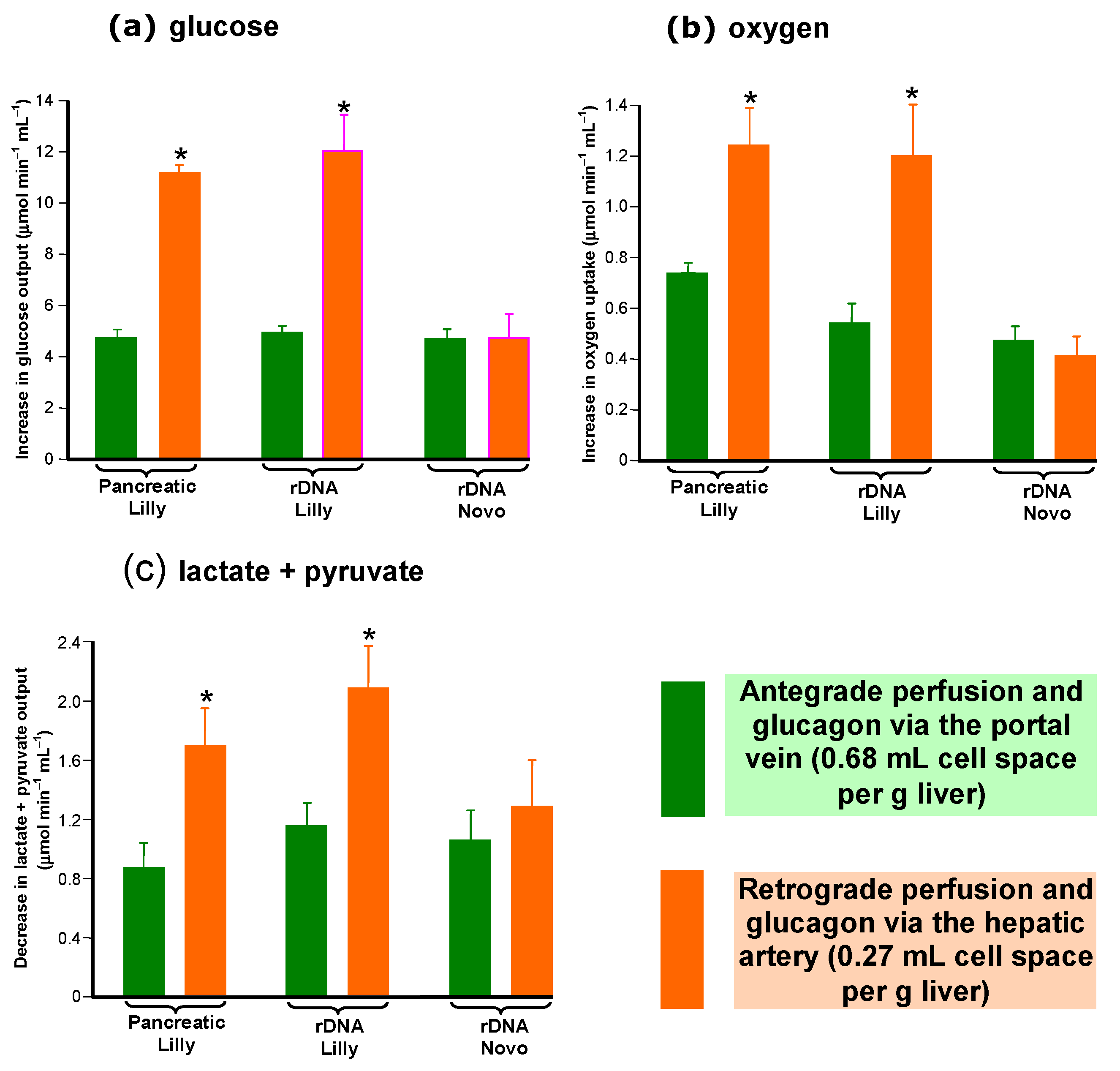
3.2. Cell Space-Normalized Changes of the rDNA Glucagon Preparations
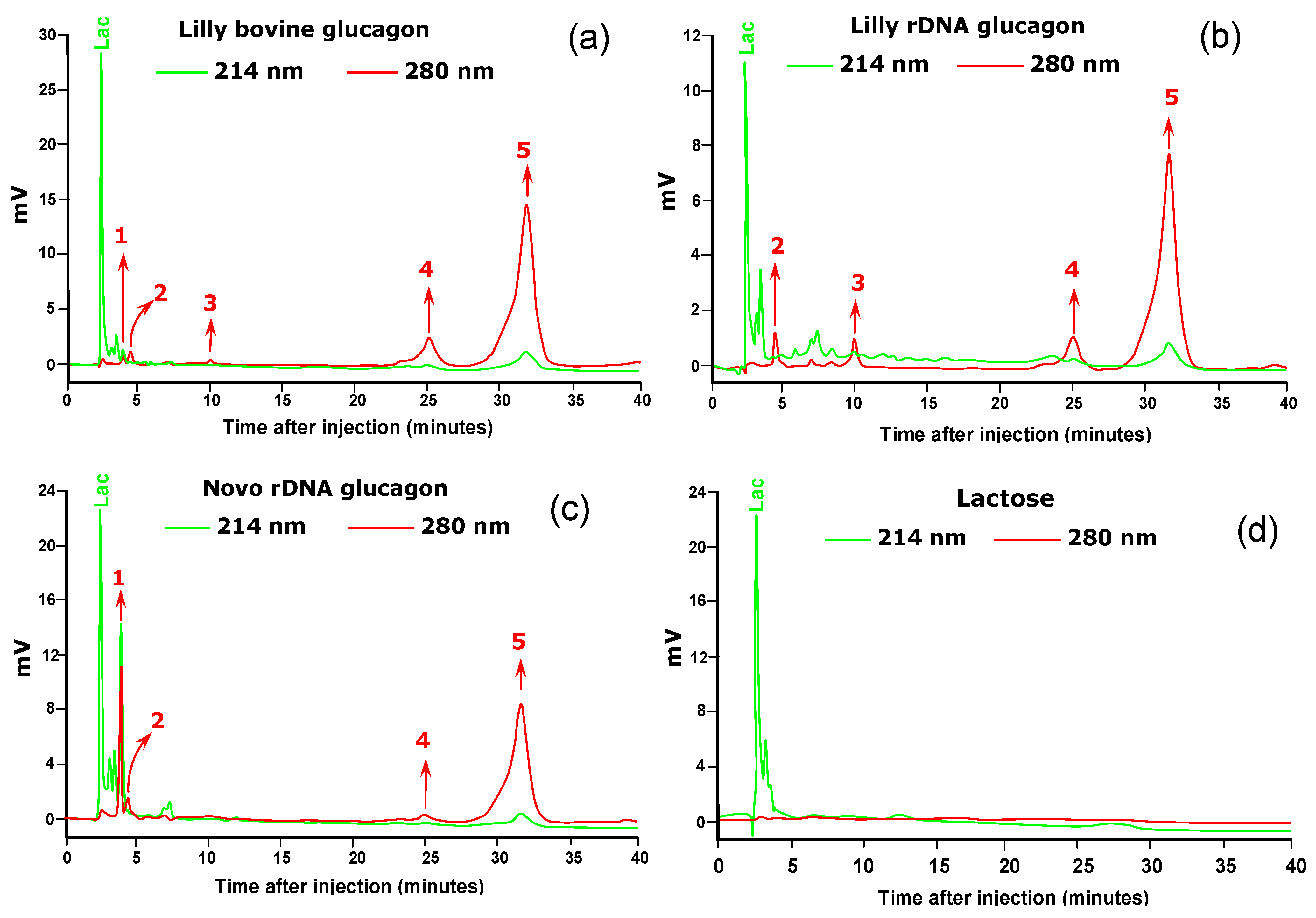
3.3. HPLC Chromatography of the Glucagon Preparations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pang, S.; Cherry, W.F.; Accaputo, J.; Schwab, A.J.; Goresky, C.A. Combined hepatic arterial-portal venous and hepatic arterial-hepatic venous perfusion to probe the abundance of drug metabolizing activities: Perihepatic venous O-deethylation activity for phenacetin and periportal sulfation activity for acetaminophen in the once-through rat liver preparation. J. Pharmacol. Exp. Ther. 1988, 247, 690–700. [Google Scholar] [PubMed]
- Schmeisch, A.P.; Oliveira, D.S.; Ide, L.T.; Suzuki-Kemmelmeier, F.; Bracht, A. Zonation of the metabolic action of vasopressin in the bivascularly perfused rat liver. Regul. Pept. 2005, 129, 233–243. [Google Scholar] [CrossRef]
- Fernandes, T.R.L.; Suzuki-Kemmelmeier, F.; Bracht, A. The hemodynamic effects of ATP in retrograde perfusion of the bivascularly perfused rat liver. Liver Int. 2003, 23, 371–378. [Google Scholar] [CrossRef]
- Jungermann, K.; Katz, N. Metabolism of carbohydrates. In Regulation of Hepatic Metabolism (Intra- and Intercellular Compart-Mentation), 1st ed.; Jungermann, K., Kauffman, F.C., Thurman, R.G., Eds.; Plenum Press: New York, NY, USA, 1986; pp. 211–235. [Google Scholar]
- Bracht, A.; Constantin, J.; Ishii-Iwamoto, E.L.; Suzuki-Kemmelmeier, F. Zonation of gluconeogenesis from lactate and pyruvate in the rat liver studied by means of anterograde and retrograde bivascular perfusion. Biochim. Biophys. Acta 1994, 1199, 298–304. [Google Scholar] [CrossRef]
- Botini, F.F.; Suzuki-Kemmelmeier, F.; Nascimento, E.A.; Ide, L.T.; Bracht, A. Zonation of alanine metabolism in the bivascularly perfused rat liver. Liver Int. 2005, 25, 861–871. [Google Scholar] [CrossRef]
- Comar, J.F.; Suzuki-Kemmelmeier, F.; Constantin, J.; Bracht, A. Hepatic zonation of carbon and nitrogen fluxes derived from glutamine and ammonia transformations. J. Biomed. Sci. 2010, 17, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constantin, J.; Ishii-Iwamoto, E.L.; Suzuki-Kemmelmeier, F.; Bracht, A. Zonation of the action of glucagon on gluconeogenesis studied in the bivascularly perfused rat liver. FEBS Lett. 1994, 352, 24–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maskalick, D.G.; Anderson, M.T. Process for Recovering Glucagon from Pancreas Glands. U.S. Patent 4617376, 25 July 1986. Available online: https://patents.google.com/patent/EP0207727A3/en (accessed on 30 March 2023).
- Constantin, J.; Ishii-Iwamoto, E.L.; Suzuki-Kemmelmeier, F.; Yamamoto, N.S.; Bracht, A. The action of glucagon infused via the hepatic artery in anterograde and retrograde perfusion of the rat liver is not a function of the accessible cellular spaces. Biochim. Biophys. Acta 1995, 1244, 169–178. [Google Scholar] [CrossRef]
- Sáez, J.C.; Spray, D.C.; Nairn, A.C.; Hertzeberg, E.L.; Greengard, P.; Bennett, M.V.L. cAMP increases junctional conductance and stimulates phosphorylation of the 27-kDa principal gap junction polypeptide. Proc. Natl. Acad. Sci. USA 1986, 83, 2473–2477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constantin, J.; Suzuki-Kemmelmeier, F.; Yamamoto, N.S.; Bracht, A. Production, uptake, and metabolic effects of cyclic AMP in the bivascularly perfused rat liver. Biochem. Pharmacol. 1997, 54, 1115–1125. [Google Scholar] [CrossRef]
- Fonjallaz, P.; Loumaye, E. Glucagon rDNA origin (GlucaGen®) and recombinant LH. J. Biotechnol. 2000, 79, 185–189. [Google Scholar] [CrossRef]
- Ishizaki, J.; Tamaki, M.; Shin, M.; Usuzuki, H.; Yoshikawa, K.; Teraoka, H.; Yoshida, N. Production of recombinant human glucagon in the form of a fusion protein in Escherichia coli; recovery of glucagon by sequence-specific digestion. Appl. Microbiol. Biotechnol. 1992, 36, 483–486. [Google Scholar] [CrossRef]
- Norris, K.; Thim, L.; Norris, F.; Hansen, M.T.; Moody, A.J. Process for preparing glucagon or fragments or derivatives thereof in yeast. U.S. Patent 4826763, 14 January 1989. Available online: https://patents.google.com/patent/CA1294232C/en (accessed on 29 March 2023).
- Suzuki-Kemmelmeier, F.; Ishii-Iwamoto, E.L.; Bracht, A. The metabolism of fructose in the bivascularly perfused rat liver. Biochim. Biophys. Acta 1992, 1116, 275–282. [Google Scholar] [CrossRef]
- Clark, L.C. Monitor and control of blood O2 tension. ASAIO J. 1956, 2, 41–48. [Google Scholar]
- Bergmeyer, H.U.; Bernt, E. Determination of glucose with glucose oxidase and peroxidase. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Verlag Chemie-Academic Press: Weinheim, Germany; London, UK, 1974; pp. 1205–1215. [Google Scholar]
- Gutmann, I.; Wahlefeld, A.W. L-(+)-Lactate determination with lactate dehydrogenase and NAD. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Verlag Chemie-Academic Press: Weinheim, Germany; London, UK, 1974; pp. 1464–1468. [Google Scholar]
- Czok, R.; Lamprecht, W. Pyruvate, phosphoenolpyruvate and D-glycerate-2-phosphate. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Verlag Chemie-Academic Press: Weinheim, Germany; London, UK, 1974; pp. 1446–1451. [Google Scholar]
- Kimmig, R.; Mauch, T.J.; Kerzl, W.; Schwabe, U.; Scholz, R. Actions of glucagon on flux rates in perfused rat liver 1. Kinetics of the inhibitory effect on glycolysis and the stimulatory effect on glycogenolysis. Eur. J. Biochem. 1983, 136, 609–616. [Google Scholar] [CrossRef]
- Comar, J.F.; de Oliveira, D.S.; Bracht, L.; Kemmelmeier, F.S.; Peralta, R.M.; Bracht, A. The metabolic responses to L-glutamine of livers from rats with diabetes types 1 and 2. PLoS ONE 2016, 11, e0160067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimmig, R.; Mauch, T.J.; Scholz, R. Actions of glucagon on flux rates in perfused rat liver 2. Relationship between inhibition of glycolysis and stimulation of respiration by glucagon. Eur. J. Biochem. 1983, 136, 617–620. [Google Scholar] [CrossRef]
- Mojsov, S.; Merrifield, R.B. An improved synthesis of crystalline mammalian glucagon. Eur. J. Biochem. 1984, 145, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Joshi, A.B.; Rus, E.; Kirsch, L.E. The degradation pathways of glucagon in acidic solutions. Int. J. Pharm. 2000, 203, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Sethi, M.K.; Thakan, A.; Mahajan, S.; Mara, B.; Yerramalla, R.; Vemula, L.; Thirunavukarasu, J.; Bandi, N.; Vakale, K.A.; Reddy, V.R.; et al. Preparation of glucagon impurities related to lactose formulations. Asian J. Pharm. Anal. Med. Chem. 2020, 8, 137–146. [Google Scholar]
- Han, Q.; Bao, Z.; Luo, M.Z.; Zhang, J.Y. Assessment of innate immune response modulating impurities in glucagon for injection. PLoS ONE 2022, 17, e0277922. [Google Scholar] [CrossRef]
- Bao, Z.; Cheng, Y.C.; Luo, M.Z.; Zhang, Y. Comparison of the purity and impurity of glucagon-for-injection products under various stability conditions. Sci. Pharm. 2022, 90, 32. [Google Scholar] [CrossRef]
- Stümpel, F.; Ott, T.; Willecke, K.; Jungermann, K. Connexin 3 gap junctions enhance stimulation of glucose output by glucagon and noradrenaline in mouse liver. Hepatology 1998, 28, 1616–1620. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Clemens, M.G. Glucagon increases gap junctional intercellular communication via cAMP in the isolated perfused rat liver. Shock 2004, 22, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Habegger, K.M. Cross talk between insulin and glucagon receptor signaling in the hepatocyte. Diabetes 2022, 71, 1842–1851. [Google Scholar] [CrossRef]
- Kawamori, D.; Sasaki, S. Newly Discovered Knowledge Pertaining to Glucagon and Its Clinical Applications. J. Diabetes Investig. 2023; online ahead of print. [Google Scholar] [CrossRef]
- Kaur, J.; Seaquist, E.R. Translational aspects of glucagon: Current use and future prospects. J. Endocrinol. 2023, 257, e22078. [Google Scholar] [CrossRef]
- Novikoff, A.; Müller, T.M. The molecular pharmacology of glucagon in diabetes and obesity. Peptides 2023, 165, 171003. [Google Scholar] [CrossRef]
- Ahrén, B. Glucagon—Early breakthroughs and recent discoveries. Peptides 2015, 67, 74–81. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peaks at 280 nm | Pancreatic Glucagon Elli Lilly | rDNA Glucagon Elli Lilly | rDNA Glucagon Novo | |||
|---|---|---|---|---|---|---|
| Rf (min) | Relative Area (%) | Rf (min) | Relative Area (%) | Rf (min) | Relative Area (%) | |
| 1 | 4.02 ± 0.01 | 1.12 ± 0.08 a | - | - | 3.98 ± 0.02 | 15.08 ± 0.12 a |
| 2 | 4.52 ± 0.01 | 1.29 ± 0.01 a | 4.50 ± 0.01 | 4.03 ± 0.90 a | 4.49 ± 0.01 | 2.49 ± 0.38 |
| 3 | 10.0 ± 0.03 | 0.56 ± 0.08 a | 10.10 ± 0.03 | 4.02 ± 0.43 a | - | - |
| 4 | 25.34 ± 0.03 | 12.33 ± 0.11 a,b | 25.54 ± 0.19 | 9.04 ± 0.29 a,c | 25.26 ± 0.05 | 2.36 ± 0.25 b,c |
| 5 | 32.09 ± 0.08 | 83.42 ± 0.11 a,b | 32.15 ± 0.14 | 78.14 ± 2.44 a | 32.02 ± 0.04 | 77.15 ± 0.05 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bracht, L.; Constantin, J.; Peralta, R.M.; Bracht, A. Escherichia coli (Lilly) and Saccharomyces cerevisiae (Novo) rDNA Glucagon: An Assessment of Their Actions When Supplied Selectively to Periportal Cells in the Bivascularly Perfused Rat Liver. Sci. Pharm. 2023, 91, 29. https://doi.org/10.3390/scipharm91030029
Bracht L, Constantin J, Peralta RM, Bracht A. Escherichia coli (Lilly) and Saccharomyces cerevisiae (Novo) rDNA Glucagon: An Assessment of Their Actions When Supplied Selectively to Periportal Cells in the Bivascularly Perfused Rat Liver. Scientia Pharmaceutica. 2023; 91(3):29. https://doi.org/10.3390/scipharm91030029
Chicago/Turabian StyleBracht, Lívia, Jorgete Constantin, Rosane Marina Peralta, and Adelar Bracht. 2023. "Escherichia coli (Lilly) and Saccharomyces cerevisiae (Novo) rDNA Glucagon: An Assessment of Their Actions When Supplied Selectively to Periportal Cells in the Bivascularly Perfused Rat Liver" Scientia Pharmaceutica 91, no. 3: 29. https://doi.org/10.3390/scipharm91030029