
Collateral Damage in the Placenta during Viral Infection in Pregnancy: A Possible Mechanism for Vertical Transmission and an Adverse Pregnancy Outcome

 , and
, and 
Abstract
:1. Introduction
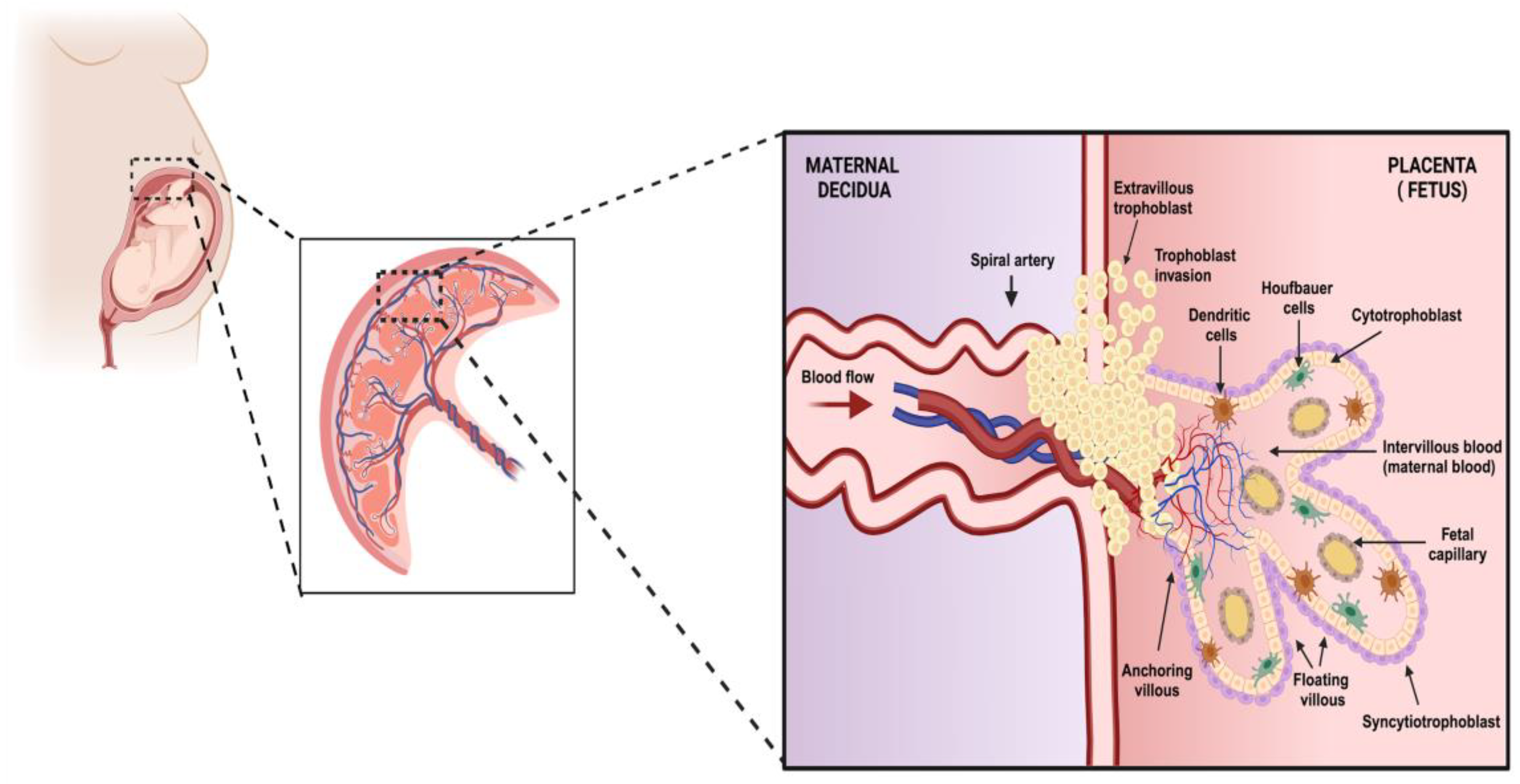
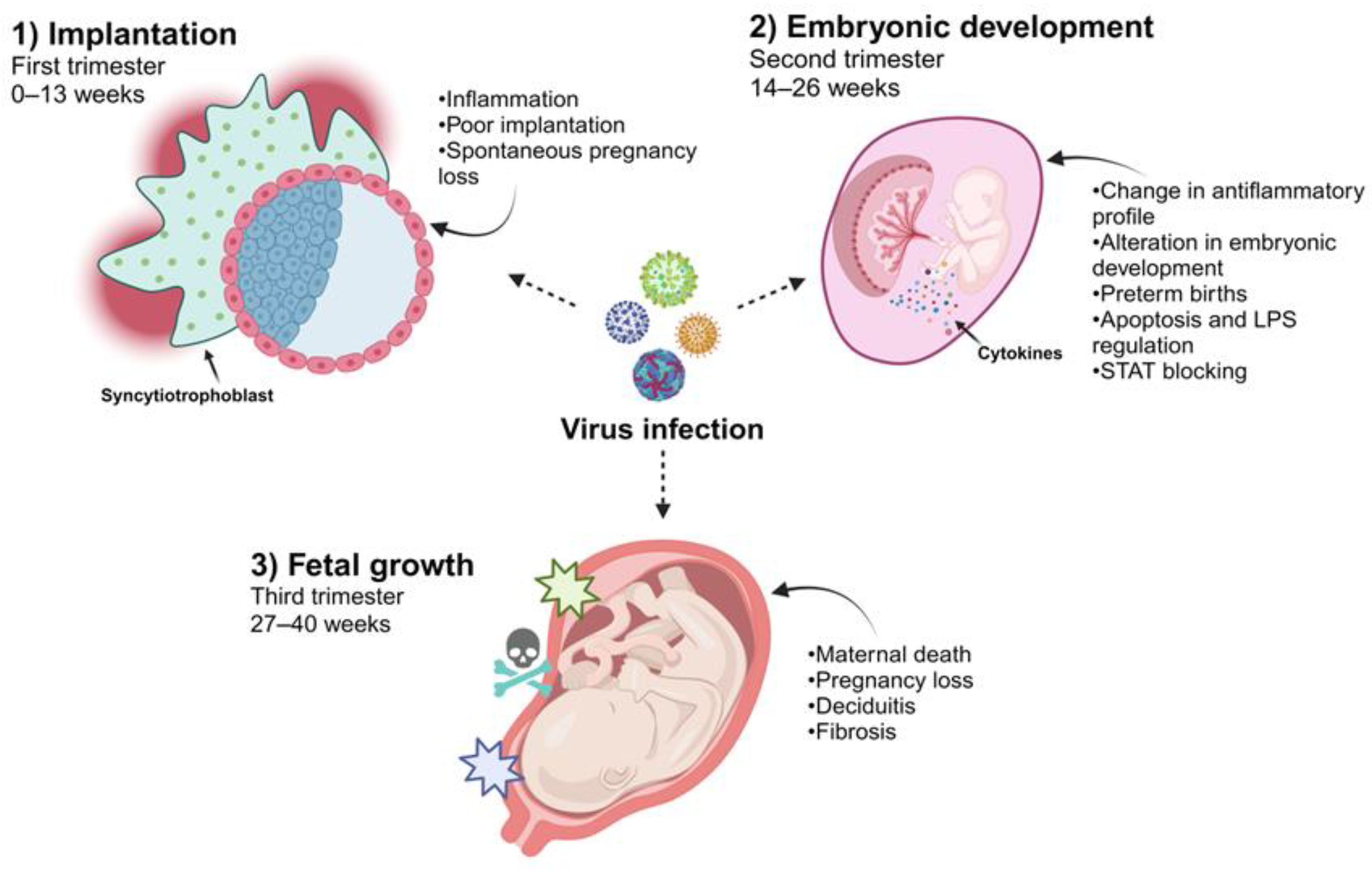
2. The Temporality of Pregnancy and the Effect of Viral Infection on the Placenta
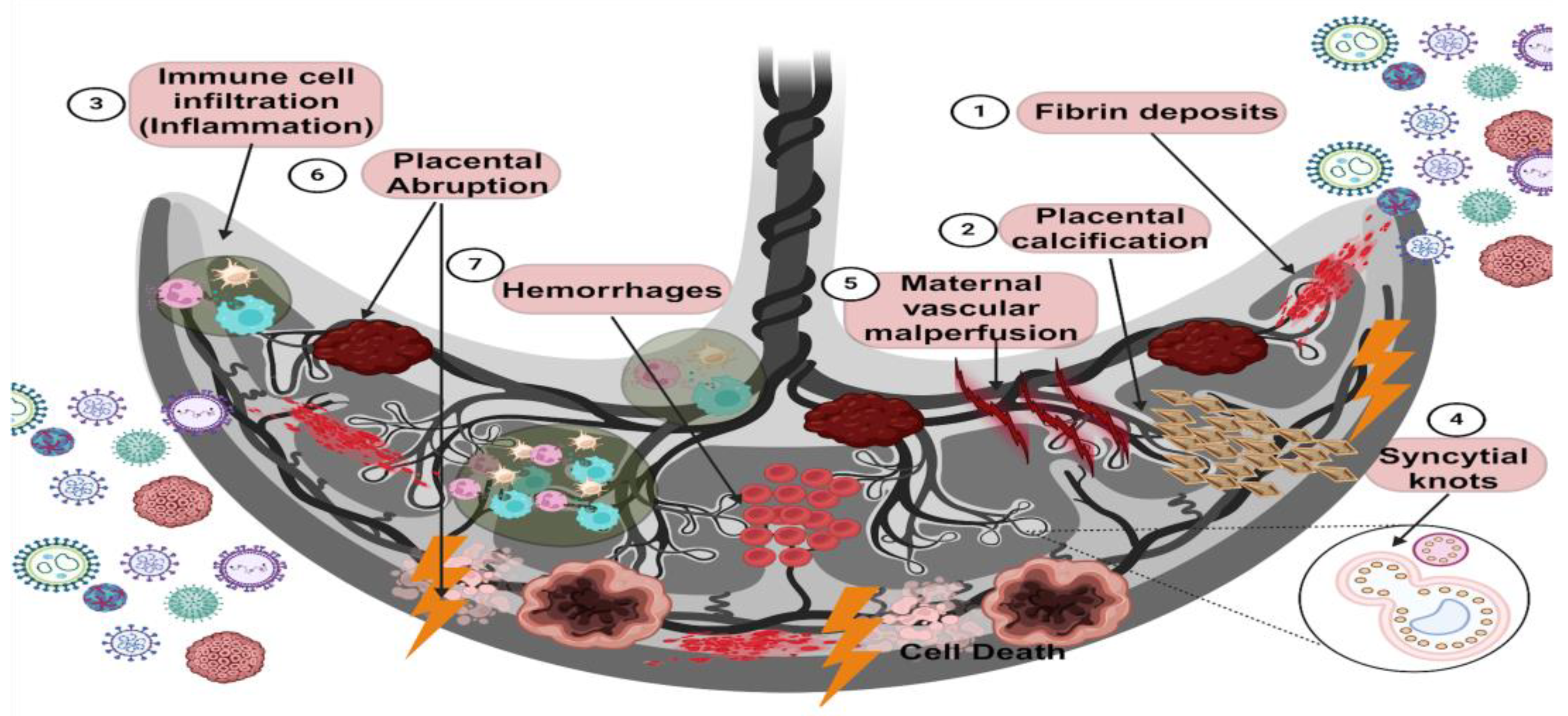
3. Histopathological Changes in the Placenta Caused by Viral Infections
4. Viral Infections That Cannot Be Transmitted Vertically during Pregnancy
4.1. Hepatitis E Virus (HEV)
4.2. Dengue Virus (DENV)
4.3. Chikungunya Virus (CHIKV)
4.4. Respiratory Syncytial Virus (RSV)
4.5. Human Bocavirus (HBoV)
4.6. Ebola Virus (EBOV) and Other Hemorrhagic Fever Viruses
4.7. Human Coronavirus
5. Viral Mechanisms Could Cause Potential Damage to the Placenta
5.1. HEV
5.2. DENV
5.3. CHIKV
5.4. RSV
5.5. EBOV and Other Hemorrhagic Fever Viruses
5.6. Human Coronaviruses
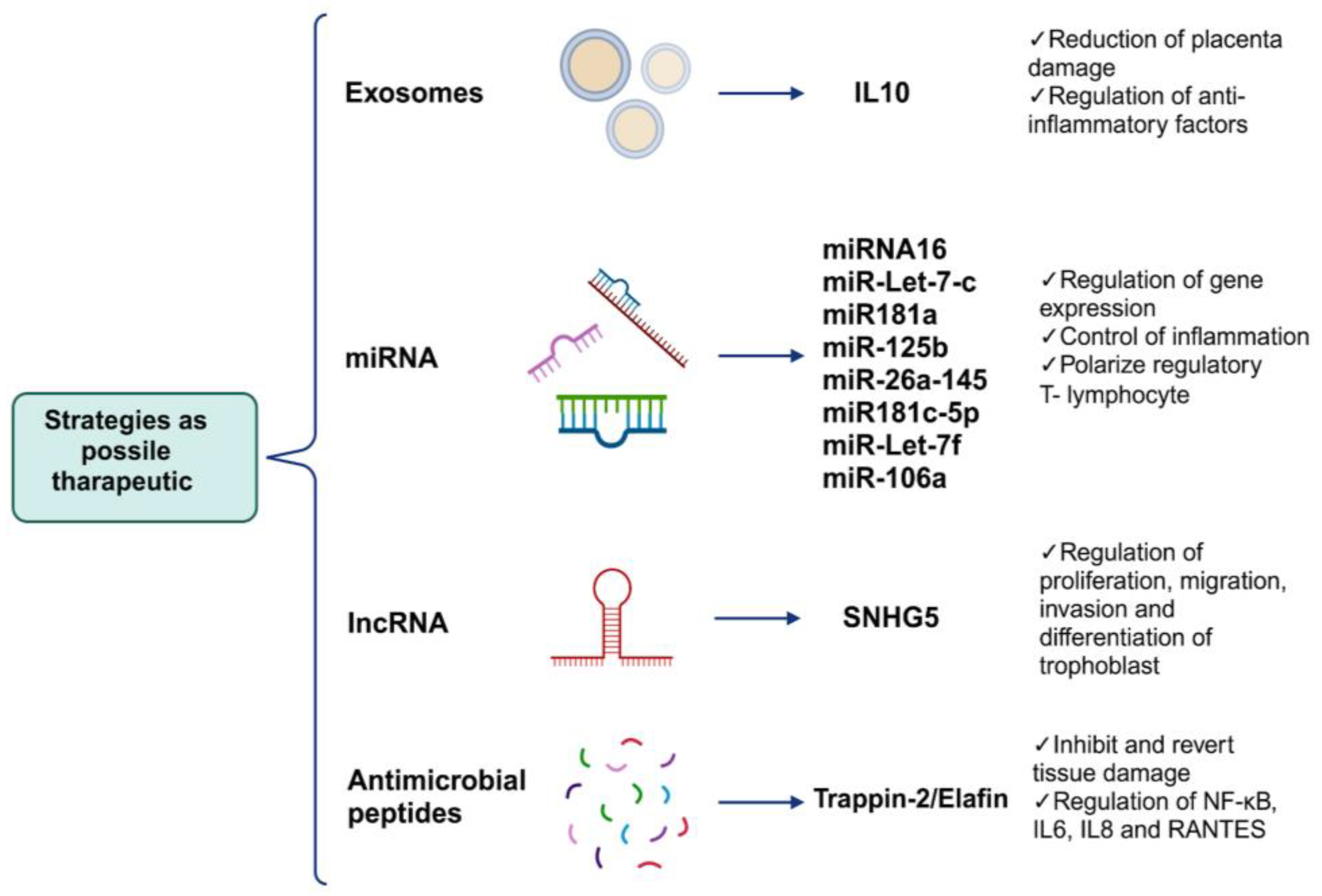
6. Alternative Strategies as Possible Therapeutic Targets Can Regulate the Mechanisms of Placental Damage
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Knöfler, M.; Haider, S.; Saleh, L.; Pollheimer, J.; Gamage, T.K.J.B.; James, J. Human Placenta and Trophoblast Development: Key Molecular Mechanisms and Model Systems. Cell. Mol. Life Sci. 2019, 76, 3479–3496. [Google Scholar] [CrossRef]
- Cindrova-Davies, T.; Sferruzzi-Perri, A.N. Human Placental Development and Function. Semin. Cell Dev. Biol. 2022, 131, 66–77. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.; Wang, Y. The Placenta: A Maternofetal Interface. Annu. Rev. Nutr. 2023, 43, 301–325. [Google Scholar] [CrossRef] [PubMed]
- Walentin, K.; Hinze, C.; Schmidt-Ott, K.M. The Basal Chorionic Trophoblast Cell Layer: An Emerging Coordinator of Placenta Development. BioEssays 2016, 38, 254–265. [Google Scholar] [CrossRef]
- Xiao, Z.; Yan, L.; Liang, X.; Wang, H. Progress in Deciphering Trophoblast Cell Differentiation during Human Placentation. Curr. Opin. Cell Biol. 2020, 67, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Moser, G.; Windsperger, K.; Pollheimer, J.; de Sousa Lopes, S.C.; Huppertz, B. Human Trophoblast Invasion: New and Unexpected Routes and Functions. Histochem. Cell Biol. 2018, 150, 361–370. [Google Scholar] [CrossRef]
- Taniguchi, K.; Kawai, T.; Hata, K. Placental Development and Nutritional Environment. Adv. Exp. Med. Biol. 2018, 1012, 63–73. [Google Scholar] [CrossRef]
- Yockey, L.J.; Iwasaki, A. Interferons and Proinflammatory Cytokines in Pregnancy and Fetal Development. Immunity 2018, 49, 397–412. [Google Scholar] [CrossRef]
- Megli, C.J.; Coyne, C.B. Infections at the Maternal-Fetal Interface: An Overview of Pathogenesis and Defence. Nat. Rev. Microbiol. 2022, 20, 67–82. [Google Scholar] [CrossRef]
- Arora, N.; Sadovsky, Y.; Dermody, T.S.; Coyne, C.B. Microbial Vertical Transmission during Human Pregnancy. Cell Host Microbe 2017, 21, 561–567. [Google Scholar] [CrossRef]
- León-Juárez, M.; Martínez-Castillo, M.; González-García, L.D.; Helguera-Repetto, A.C.; Zaga-Clavellina, V.; García-Cordero, J.; Flores-Pliego, A.; Herrera-Salazar, A.; Vázquez-Martínez, E.R.; Reyes-Muñoz, E. Cellular and Molecular Mechanisms of Viral Infection in the Human Placenta. Pathog. Dis. 2017, 75, ftx093. [Google Scholar] [CrossRef]
- Kim, C.J.; Romero, R.; Chaemsaithong, P.; Kim, J.-S. Chronic Inflammation of the Placenta: Definition, Classification, Pathogenesis, and Clinical Significance. Am. J. Obs. Gynecol. 2015, 213, S53–S69. [Google Scholar] [CrossRef] [PubMed]
- Chudnovets, A.; Liu, J.; Narasimhan, H.; Liu, Y.; Burd, I. Role of Inflammation in Virus Pathogenesis during Pregnancy. J. Virol. 2020, 95, e01381-19. [Google Scholar] [CrossRef] [PubMed]
- Narang, K.; Cheek, E.H.; Enninga, E.A.L.; Theiler, R.N. Placental Immune Responses to Viruses: Molecular and Histo-Pathologic Perspectives. Int. J. Mol. Sci. 2021, 22, 2921. [Google Scholar] [CrossRef] [PubMed]
- Creisher, P.S.; Lei, J.; Sherer, M.L.; Dziedzic, A.; Jedlicka, A.E.; Narasimhan, H.; Chudnovets, A.; Campbell, A.D.; Liu, A.; Pekosz, A.; et al. Downregulation of Transcriptional Activity, Increased Inflammation, and Damage in the Placenta Following in Utero Zika Virus Infection Is Associated with Adverse Pregnancy Outcomes. Front. Virol. 2022, 2, 782906. [Google Scholar] [CrossRef] [PubMed]
- Salomão, N.; Rabelo, K.; Avvad-Portari, E.; Basílio-de-Oliveira, C.; Basílio-de-Oliveira, R.; Ferreira, F.; Ferreira, L.; de Souza, T.M.; Nunes, P.; Lima, M.; et al. Histopathological and Immunological Characteristics of Placentas Infected with Chikungunya Virus. Front. Microbiol. 2022, 13, 1055536. [Google Scholar] [CrossRef] [PubMed]
- de Noronha, L.; Zanluca, C.; Burger, M.; Suzukawa, A.A.; Azevedo, M.; Rebutini, P.Z.; Novadzki, I.M.; Tanabe, L.S.; Presibella, M.M.; Duarte Dos Santos, C.N. Zika Virus Infection at Different Pregnancy Stages: Anatomopathological Findings, Target Cells and Viral Persistence in Placental Tissues. Front. Microbiol. 2018, 9, 2266. [Google Scholar] [CrossRef] [PubMed]
- Koi, H.; Zhang, J.; Parry, S. The Mechanisms of Placental Viral Infection. Ann. N. Y. Acad. Sci. 2001, 943, 148–156. [Google Scholar] [CrossRef]
- Costa, M.L.; Nobrega, G.d.M.; Antolini-Tavares, A. Key Infections in the Placenta. Obstet. Gynecol. Clin. N. Am. 2020, 47, 133–146. [Google Scholar] [CrossRef]
- Tabata, T.; Petitt, M.; Puerta-Guardo, H.; Michlmayr, D.; Wang, C.; Fang-Hoover, J.; Harris, E.; Pereira, L. Zika Virus Targets Different Primary Human Placental Cells, Suggesting Two Routes for Vertical Transmission. Cell Host Microbe 2016, 20, 155–166. [Google Scholar] [CrossRef]
- Buxmann, H.; Hamprecht, K.; Meyer-Wittkopf, M.; Friese, K. Primary Human Cytomegalovirus (HCMV) Infection in Pregnancy. Dtsch. Arztebl. Int. 2017, 114, 45–52. [Google Scholar] [CrossRef]
- Spano, L.C.; Lima Pereira, F.E.; Gomes da Silva Basso, N.; Mercon-de-Vargas, P.R. Human Cytomegalovirus Infection and Abortion: An Immunohistochemical Study. Med. Sci. Monit. 2002, 8, BR230–BR235. [Google Scholar]
- Kong, Q.; Li, J.; Zhao, L.; Shi, P.; Liu, X.; Bian, C.; Liu, J.; Liu, T. Human Cytomegalovirus Inhibits the Proliferation and Invasion of Extravillous Cytotrophoblasts via Hippo-YAP Pathway. Virol. J. 2021, 18, 214. [Google Scholar] [CrossRef]
- Liu, T.; Zheng, X.; Li, Q.; Chen, J.; Yin, Z.; Xiao, J.; Zhang, D.; Li, W.; Qiao, Y.; Chen, S. Role of Human Cytomegalovirus in the Proliferation and Invasion of Extravillous Cytotrophoblasts Isolated from Early Placentae. Int. J. Clin. Exp. Med. 2015, 8, 17248–17260. [Google Scholar]
- Zhuang, B.; Shang, J.; Yao, Y. HLA-G: An Important Mediator of Maternal-Fetal Immune-Tolerance. Front. Immunol. 2021, 12, 744324. [Google Scholar] [CrossRef]
- Easterfield, A.J.; Austen, B.M.; Westwood, O.M. Inhibition of Antigen Transport by Expression of Infected Cell Peptide 47 (ICP47) Prevents Cell Surface Expression of HLA in Choriocarcinoma Cell Lines. J. Reprod. Immunol. 2001, 50, 19–40. [Google Scholar] [CrossRef]
- Schust, D.J.; Tortorella, D.; Ploegh, H.L. HLA-G and HLA-C at the Feto-Maternal Interface: Lessons Learned from Pathogenic Viruses. Semin. Cancer Biol. 1999, 9, 37–46. [Google Scholar] [CrossRef]
- Schust, D.J.; Hill, A.B.; Ploegh, H.L. Herpes Simplex Virus Blocks Intracellular Transport of HLA-G in Placentally Derived Human Cells. J. Immunol. 1996, 157, 3375–3380. [Google Scholar] [CrossRef]
- Basonidis, A.; Liberis, A.; Daniilidis, A.; Petousis, S.; Dinas, K. Human Papilloma Virus Infection and Miscarriage: Is There an Association? Taiwan. J. Obs. Gynecol. 2020, 59, 656–659. [Google Scholar] [CrossRef]
- Giakoumelou, S.; Wheelhouse, N.; Cuschieri, K.; Entrican, G.; Howie, S.E.M.; Horne, A.W. The Role of Infection in Miscarriage. Hum. Reprod. Update 2016, 22, 116–133. [Google Scholar] [CrossRef]
- Condrat, C.E.; Filip, L.; Gherghe, M.; Cretoiu, D.; Suciu, N. Maternal HPV Infection: Effects on Pregnancy Outcome. Viruses 2021, 13, 2455. [Google Scholar] [CrossRef]
- Foresta, C.; Patassini, C.; Bertoldo, A.; Menegazzo, M.; Francavilla, F.; Barzon, L.; Ferlin, A. Mechanism of Human Papillomavirus Binding to Human Spermatozoa and Fertilizing Ability of Infected Spermatozoa. PLoS ONE 2011, 6, e15036. [Google Scholar] [CrossRef]
- Mastora, E.; Kitsou, C.; Evangelou, T.; Zikopoulos, A.; Zagorianakou, N.; Georgiou, I. Presence of HPV 16 and HPV 18 in Spermatozoa and Embryos of Mice. In Vivo 2021, 35, 3203–3209. [Google Scholar] [CrossRef]
- Calinisan, J.H.; Chan, S.R.; King, A.; Chan, P.J. Human Papillomavirus and Blastocyst Apoptosis. J. Assist. Reprod. Genet. 2002, 19, 132–136. [Google Scholar] [CrossRef]
- Ander, S.E.; Diamond, M.S.; Coyne, C.B. Immune Responses at the Maternal-Fetal Interface. Sci. Immunol. 2019, 4, eaat6114. [Google Scholar] [CrossRef]
- Kumpel, B.M.; Manoussaka, M.S. Placental Immunology and Maternal Alloimmune Responses. Vox Sang. 2012, 102, 2–12. [Google Scholar] [CrossRef]
- Morelli, S.S.; Mandal, M.; Goldsmith, L.T.; Kashani, B.N.; Ponzio, N.M. The Maternal Immune System during Pregnancy and Its Influence on Fetal Development. Res. Rep. Biol. 2015, 6, 171–189. [Google Scholar] [CrossRef]
- Price, M.E.; Fisher-Hoch, S.P.; Craven, R.B.; McCormick, J.B. A Prospective Study of Maternal and Fetal Outcome in Acute Lassa Fever Infection during Pregnancy. BMJ 1988, 297, 584–587. [Google Scholar] [CrossRef]
- Okogbenin, S.; Okoeguale, J.; Akpede, G.; Colubri, A.; Barnes, K.G.; Mehta, S.; Eifediyi, R.; Okogbo, F.; Eigbefoh, J.; Momoh, M.; et al. Retrospective Cohort Study of Lassa Fever in Pregnancy, Southern Nigeria. Emerg. Infect. Dis. 2019, 25, 1494–1500. [Google Scholar] [CrossRef]
- Adewole, A.A.; Amode, O.; Gbejegbe, E.H.; Musa, A.O.; Adamu, A.N.; Ojotule, A.; Ladan, A.A.; Ojiah, A.A.; Adesanya, A.; Olarinoye, G.M.; et al. Lassa Fever in Pregnancy: Report of Two Maternal Deaths in a Tertiary Center in the Middle-Belt Region of Nigeria. Afr. J. Reprod. Health 2022, 26, 137–143. [Google Scholar]
- Borrow, P.; Martínez-Sobrido, L.; De la Torre, J.C. Inhibition of the Type I Interferon Antiviral Response During Arenavirus Infection. Viruses 2010, 2, 2443–2480. [Google Scholar] [CrossRef]
- Hastie, K.M.; Kimberlin, C.R.; Zandonatti, M.A.; MacRae, I.J.; Saphire, E.O. Structure of the Lassa Virus Nucleoprotein Reveals a dsRNA-Specific 3′ to 5′ Exonuclease Activity Essential for Immune Suppression. Proc. Natl. Acad. Sci. USA 2011, 108, 2396–2401. [Google Scholar] [CrossRef]
- Reynard, S.; Russier, M.; Fizet, A.; Carnec, X.; Baize, S. Exonuclease Domain of the Lassa Virus Nucleoprotein Is Critical To Avoid RIG-I Signaling and To Inhibit the Innate Immune Response. J. Virol. 2014, 88, 13923–13927. [Google Scholar] [CrossRef]
- Bebell, L.M.; Oduyebo, T.; Riley, L.E. Ebola Virus Disease and Pregnancy: A Review of the Current Knowledge of Ebola Virus Pathogenesis, Maternal, and Neonatal Outcomes. Birth Defects Res. 2017, 109, 353–362. [Google Scholar] [CrossRef]
- Fallah, M.P.; Skrip, L.A.; Dahn, B.T.; Nyenswah, T.G.; Flumo, H.; Glayweon, M.; Lorseh, T.L.; Kaler, S.G.; Higgs, E.S.; Galvani, A.P. Pregnancy Outcomes in Liberian Women Who Conceived after Recovery from Ebola Virus Disease. Lancet Glob. Health 2016, 4, e678–e679. [Google Scholar] [CrossRef]
- Kayem, N.D.; Benson, C.; Aye, C.Y.L.; Barker, S.; Tome, M.; Kennedy, S.; Ariana, P.; Horby, P. Ebola Virus Disease in Pregnancy: A Systematic Review and Meta-Analysis. Trans. R. Soc. Trop. Med. Hyg. 2022, 116, 509–522. [Google Scholar] [CrossRef]
- He, F.; Melén, K.; Maljanen, S.; Lundberg, R.; Jiang, M.; Österlund, P.; Kakkola, L.; Julkunen, I. Ebolavirus Protein VP24 Interferes with Innate Immune Responses by Inhibiting Interferon-Λ1 Gene Expression. Virology 2017, 509, 23–34. [Google Scholar] [CrossRef]
- Falasca, L.; Agrati, C.; Petrosillo, N.; Di Caro, A.; Capobianchi, M.R.; Ippolito, G.; Piacentini, M. Molecular Mechanisms of Ebola Virus Pathogenesis: Focus on Cell Death. Cell Death Differ. 2015, 22, 1250–1259. [Google Scholar] [CrossRef]
- Ding, J.; Maxwell, A.; Adzibolosu, N.; Hu, A.; You, Y.; Liao, A.; Mor, G. Mechanisms of Immune Regulation by The Placenta: Role of Type I Interferon and Interferon Stimulated Genes Signaling During Pregnancy. Immunol. Rev. 2022, 308, 9–24. [Google Scholar] [CrossRef]
- Foley, J.H.; Conway, E.M. Cross Talk Pathways Between Coagulation and Inflammation. Circ. Res. 2016, 118, 1392–1408. [Google Scholar] [CrossRef]
- Antoniak, S.; Mackman, N. Multiple Roles of the Coagulation Protease Cascade during Virus Infection. Blood 2014, 123, 2605–2613. [Google Scholar] [CrossRef]
- Flores-Pliego, A.; Miranda, J.; Vega-Torreblanca, S.; Valdespino-Vázquez, Y.; Helguera-Repetto, C.; Espejel-Nuñez, A.; Borboa-Olivares, H.; Espino y Sosa, S.; Mateu-Rogell, P.; León-Juárez, M.; et al. Molecular Insights into the Thrombotic and Microvascular Injury in Placental Endothelium of Women with Mild or Severe COVID-19. Cells 2021, 10, 364. [Google Scholar] [CrossRef]
- Camen, I.V.; Istrate-Ofiţeru, A.M.; Novac, L.V.; Manolea, M.M.; Dijmărescu, A.L.; Neamţu, S.D.; Radu, L.; Boldeanu, M.V.; Şerbănescu, M.S.; Stoica, M.; et al. Analysis of the Relationship between Placental Histopathological Aspects of Preterm and Term Birth. Rom. J. Morphol. Embryol. 2022, 63, 357–367. [Google Scholar] [CrossRef]
- Khuroo, M.S.; Kamili, S.; Khuroo, M.S. Clinical Course and Duration of Viremia in Vertically Transmitted Hepatitis E Virus (HEV) Infection in Babies Born to HEV-Infected Mothers. J. Viral Hepat. 2009, 16, 519–523. [Google Scholar] [CrossRef]
- Galazios, G.; Tsoulou, S.; Zografou, C.; Tripsianis, G.; Koutlaki, N.; Papazoglou, D.; Tsikouras, P.; Maltezos, E.; Liberis, V. The Role of Cytokines IL-6 and IL-8 in the Pathogenesis of Spontaneous Abortions. J. Matern.-Fetal Neonatal Med. 2011, 24, 1283–1285. [Google Scholar] [CrossRef]
- Jilani, N.; Das, B.C.; Husain, S.A.; Baweja, U.K.; Chattopadhya, D.; Gupta, R.K.; Sardana, S.; Kar, P. Hepatitis E Virus Infection and Fulminant Hepatic Failure during Pregnancy. J. Gastroenterol. Hepatol. 2007, 22, 676–682. [Google Scholar] [CrossRef]
- Qian, Z.; Li, T.; Xia, Y.; Cong, C.; Chen, S.; Zhang, Y.; Gong, S.; Wang, W.; Liu, H.; Chen, D.; et al. Genotype 4 Hepatitis E Virus Replicates in the Placenta, Causes Severe Histopathological Damage, and Vertically Transmits to Fetuses. J. Infect. 2023, 87, 34–45. [Google Scholar] [CrossRef]
- Feitoza, H.A.C.; Koifman, S.; Koifman, R.J.; Saraceni, V. Dengue infection during pregnancy and adverse maternal, fetal, and infant health outcomes in Rio Branco, Acre State, Brazil, 2007–2012. Cad Saude Publica 2017, 33, e00178915. [Google Scholar] [CrossRef]
- Nunes, P.; Nogueira, R.; Coelho, J.; Rodrigues, F.; Salomão, N.; José, C.; de Carvalho, J.; Rabelo, K.; de Azeredo, E.; Basílio-de-Oliveira, R.; et al. A Stillborn Multiple Organs’ Investigation from a Maternal DENV-4 Infection: Histopathological and Inflammatory Mediators Characterization. Viruses 2019, 11, 319. [Google Scholar] [CrossRef]
- Fonceca, A.M.; Chopra, A.; Levy, A.; Noakes, P.S.; Poh, M.W.-P.; Bear, N.L.; Prescott, S.; Everard, M.L. Infective Respiratory Syncytial Virus Is Present in Human Cord Blood Samples and Most Prevalent during Winter Months. PLoS ONE 2017, 12, e0173738. [Google Scholar] [CrossRef]
- Hause, A.M.; Avadhanula, V.; Maccato, M.L.; Pinell, P.M.; Bond, N.; Santarcangelo, P.; Ferlic-Stark, L.; Ye, X.; Iwuchukwu, O.; Maurer, L.; et al. Clinical Characteristics and Outcomes of Respiratory Syncytial Virus Infection in Pregnant Women. Vaccine 2019, 37, 3464–3471. [Google Scholar] [CrossRef] [PubMed]
- Shabynina, N.R.; Tulakina, L.G.; Poluiakhtova, M.V.; Golovko, V.D. Ultrastructural pathology of the placenta in respiratory-syncytial viral infection and mycoplasmosis. Arkhiv Patol. 1993, 55, 54–57. [Google Scholar]
- Hansen, M.; Brockmann, M.; Schildgen, V.; Schildgen, O. Human Bocavirus Is Detected in Human Placenta and Aborted Tissues. Influenza Other Respir. Viruses 2019, 13, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Mupapa, K.; Mukundu, W.; Bwaka, M.A.; Kipasa, M.; De Roo, A.; Kuvula, K.; Kibadi, K.; Massamba, M.; Ndaberey, D.; Colebunders, R.; et al. Ebola Hemorrhagic Fever and Pregnancy. J. Infect. Dis. 1999, 179 (Suppl. S1), S11–S12. [Google Scholar] [CrossRef] [PubMed]
- Muehlenbachs, A.; de la Rosa Vázquez, O.; Bausch, D.G.; Schafer, I.J.; Paddock, C.D.; Nyakio, J.P.; Lame, P.; Bergeron, E.; McCollum, A.M.; Goldsmith, C.S.; et al. Ebola Virus Disease in Pregnancy: Clinical, Histopathologic, and Immunohistochemical Findings. J. Infect. Dis. 2017, 215, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Semerci, N.; De Assis, V.; Kayisli, U.A.; Schatz, F.; Steffensen, T.S.; Guzeloglu-Kayisli, O.; Lockwood, C.J. Regulation of Proinflammatory Molecules and Tissue Factor by SARS-CoV-2 Spike Protein in Human Placental Cells: Implications for SARS-CoV-2 Pathogenesis in Pregnant Women. Front. Immunol. 2022, 13, 876555. [Google Scholar] [CrossRef]
- Salomão, N.; Araújo, L.; Rabelo, K.; Avvad-Portari, E.; de Souza, L.; Fernandes, R.; Valle, N.; Ferreira, L.; Basílio-de-Oliveira, C.; Basílio-de-Oliveira, R.; et al. Placental Alterations in a Chikungunya-Virus-Infected Pregnant Woman: A Case Report. Microorganisms 2022, 10, 872. Available online: https://www.mdpi.com/2076-2607/10/5/872 (accessed on 22 December 2023). [CrossRef]
- Parcial, A.L.N.; Salomão, N.G.; Portari, E.A.; Arruda, L.V.; de Carvalho, J.J.; de Matos Guedes, H.L.; Conde, T.C.; Moreira, M.E.; Batista, M.M.; Paes, M.V.; et al. SARS-CoV-2 Is Persistent in Placenta and Causes Macroscopic, Histopathological, and Ultrastructural Changes. Viruses 2022, 14, 1885. [Google Scholar] [CrossRef]
- Yeşilbaş, O.; Kıhtır, H.S.; Talip Petmezci, M.; Balkaya, S.; Hatipoğlu, N.; Meşe, S.; Şevketoğlu, E. Very rare and life-threatening complications of bocavirus bronchiolitis: Pneumomediastinum and bilateral pneumothorax. Mikrobiyol. Bul. 2016, 50, 159–164. [Google Scholar] [CrossRef]
- Jamieson, D.J.; Uyeki, T.M.; Callaghan, W.M.; Meaney-Delman, D.; Rasmussen, S.A. What Obstetrician-Gynecologists Should Know about Ebola: A Perspective from the Centers for Disease Control and Prevention. Obs. Gynecol. 2014, 124, 1005–1010. [Google Scholar] [CrossRef]
- Ratho, R.K.; Thakur, V.; Arya, S.; Singh, M.P.; Suri, V.; Das, A. Placenta as a Site of HEV Replication and Inflammatory Cytokines Modulating the Immunopathogenesis of HEV in Pregnant Women. J. Med. Virol. 2022, 94, 3457–3463. [Google Scholar] [CrossRef] [PubMed]
- Patberg, E.T.; Adams, T.; Rekawek, P.; Vahanian, S.A.; Akerman, M.; Hernandez, A.; Rapkiewicz, A.V.; Ragolia, L.; Sicuranza, G.; Chavez, M.R.; et al. Coronavirus Disease 2019 Infection and Placental Histopathology in Women Delivering at Term. Am. J. Obs. Gynecol 2021, 224, 382.e1–382.e18. [Google Scholar] [CrossRef] [PubMed]
- Shchegolev, A.I.; Kulikova, G.V.; Lyapin, V.M.; Shmakov, R.G.; Sukhikh, G.T. The Number of Syncytial Knots and VEGF Expression in Placental Villi in Parturient Woman with COVID-19 Depends on the Disease Severity. Bull. Exp. Biol. Med. 2021, 171, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Eloundou, S.N.; Lee, J.; Wu, D.; Lei, J.; Feller, M.C.; Ozen, M.; Zhu, Y.; Hwang, M.; Jia, B.; Xie, H.; et al. Placental Malperfusion in Response to Intrauterine Inflammation and Its Connection to Fetal Sequelae. PLoS ONE 2019, 14, e0214951. [Google Scholar] [CrossRef]
- Valdespino-Vázquez, M.Y.; Helguera-Repetto, C.A.; León-Juárez, M.; Villavicencio-Carrisoza, O.; Flores-Pliego, A.; Moreno-Verduzco, E.R.; Díaz-Pérez, D.L.; Villegas-Mota, I.; Carrasco-Ramírez, E.; López-Martínez, I.E.; et al. Fetal and Placental Infection with SARS-CoV-2 in Early Pregnancy. J. Med. Virol. 2021, 93, 4480–4487. [Google Scholar] [CrossRef]
- Vivanti, A.J.; Vauloup-Fellous, C.; Prevot, S.; Zupan, V.; Suffee, C.; Do Cao, J.; Benachi, A.; De Luca, D. Transplacental Transmission of SARS-CoV-2 Infection. Nat. Commun. 2020, 11, 3572. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, J.; Liao, T.; Chen, S.; Yan, J.; Lin, X. Maternal HBsAg Carriers and Pregnancy Outcomes: A Retrospective Cohort Analysis of 85,190 Pregnancies. BMC Pregnancy Childbirth 2020, 20, 724. [Google Scholar] [CrossRef]
- Hosier, H.; Farhadian, S.F.; Morotti, R.A.; Deshmukh, U.; Lu-Culligan, A.; Campbell, K.H.; Yasumoto, Y.; Vogels, C.B.; Casanovas-Massana, A.; Vijayakumar, P.; et al. SARS-CoV-2 Infection of the Placenta. J. Clin. Investig. 2020, 130, 4947–4953. [Google Scholar] [CrossRef]
- Touret, Y.; Randrianaivo, H.; Michault, A.; Schuffenecker, I.; Kauffmann, E.; Lenglet, Y.; Barau, G.; Fourmaintraux, A. Early maternal-fetal transmission of the Chikungunya virus. Presse Med. 2006, 35, 1656–1658. [Google Scholar] [CrossRef]
- Bello, O.O.; Akinajo, O.R.; Odubamowo, K.H.; Oluwasola, T.A.O. Lassa Fever in Pregnancy: Report of 2 Cases Seen at the University College Hospital, Ibadan. Case Rep. Obstet. Gynecol. 2016, 2016, 9673683. Available online: https://www.hindawi.com/journals/criog/2016/9673683/ (accessed on 29 December 2023). [CrossRef]
- Ashworth, C.T.; Stouffer, J.G. A Study of Fibrin Deposition in the Placenta. Its Clinical Significance. Am. J. Clin. Pathol. 1956, 26, 1031–1043. [Google Scholar] [CrossRef]
- Stalker, A.L. Fibrin Deposition in Pregnancy. J. Clin. Pathol. Suppl. 1976, 10, 70–76. [Google Scholar] [CrossRef]
- Labarrere, C.; Mullen, E. Fibrinoid and Trophoblastic Necrosis with Massive Chronic Intervillositis: An Extreme Variant of Villitis of Unknown Etiology. Am. J. Reprod. Immunol. Microbiol. 1987, 15, 85–91. [Google Scholar] [CrossRef]
- Nelson, D.M. Apoptotic Changes Occur in Syncytiotrophoblast of Human Placental Villi Where Fibrin Type Fibrinoid Is Deposited at Discontinuities in the Villous Trophoblast. Placenta 1996, 17, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.H.; Chen, L.R.; Lee, Y.H. Exploring the Relationship between Preterm Placental Calcification and Adverse Maternal and Fetal Outcome. Ultrasound Obstet. Gynecol. 2011, 37, 328–334. [Google Scholar] [CrossRef]
- Poggi, S.H.; Bostrom, K.I.; Demer, L.L.; Skinner, H.C.; Koos, B.J. Placental Calcification: A Metastatic Process? Placenta 2001, 22, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Martinovic, S.; Latin, V.; Suchanek, E.; Stavljenic-Rukavina, A.; Sampath, K.I.; Vukicevic, S. Osteogenic Protein-1 Is Produced by Human Fetal Trophoblasts In Vivo and Regulates the Synthesis of Chorionic Gonadotropin and Progesterone by Trophoblasts In Vitro. Eur. J. Clin. Chem. Clin. Biochem. 1996, 34, 103–109. Available online: https://www.degruyter.com/document/doi/10.1515/cclm.1996.34.2.103/html (accessed on 4 January 2024). [CrossRef] [PubMed]
- Laurent, A.; Rouillac, C.; Delezoide, A.-L.; Giovangrandi, Y.; Vekemans, M.; Bellet, D.; Abitbol, M.; Vidaud, M. Insulin-like 4 (INSL4) Gene Expression in Human Embryonic and Trophoblastic Tissues. Mol. Reprod. Dev. 1998, 51, 123–129. [Google Scholar] [CrossRef]
- Paralkar, V.M.; Vail, A.L.; Grasser, W.A.; Brown, T.A.; Xu, H.; Vukicevic, S.; Ke, H.Z.; Qi, H.; Owen, T.A.; Thompson, D.D. Cloning and Characterization of a Novel Member of the Transforming Growth Factor-β/Bone Morphogenetic Protein Family. J. Biol. Chem. 1998, 273, 13760–13767. [Google Scholar] [CrossRef] [PubMed]
- Wallingford, M.C.; Benson, C.; Chavkin, N.W.; Chin, M.T.; Frasch, M.G. Placental Vascular Calcification and Cardiovascular Health: It Is Time to Determine How Much of Maternal and Offspring Health Is Written in Stone. Front. Physiol. 2018, 9, 1044. [Google Scholar] [CrossRef] [PubMed]
- Tzimas, G.N.; Afshar, M.; Emadali, A.; Chevet, E.; Vali, H.; Metrakos, P.P. Correlation of Cell Necrosis and Tissue Calcification with Ischemia/Reperfusion Injury after Liver Transplantation. Transpl. Proc. 2004, 36, 1766–1768. [Google Scholar] [CrossRef]
- Proudfoot, D. Calcium Signaling and Tissue Calcification. Cold Spring Harb. Perspect. Biol. 2019, 11, a035303. [Google Scholar] [CrossRef]
- Proudfoot, D.; Skepper, J.N.; Hegyi, L.; Bennett, M.R.; Shanahan, C.M.; Weissberg, P.L. Apoptosis Regulates Human Vascular Calcification In Vitro. Circ. Res. 2000, 87, 1055–1062. [Google Scholar] [CrossRef]
- Mastrolia, S.A.; Weintraub, A.Y.; Sciaky-Tamir, Y.; Tirosh, D.; Loverro, G.; Hershkovitz, R. Placental Calcifications: A Clue for the Identification of High-Risk Fetuses in the Low-Risk Pregnant Population? J. Matern.-Fetal Neonatal Med. 2016, 29, 921–927. [Google Scholar] [CrossRef]
- Chen, K.-H.; Seow, K.-M.; Chen, L.-R. The Role of Preterm Placental Calcification on Assessing Risks of Stillbirth. Placenta 2015, 36, 1039–1044. [Google Scholar] [CrossRef] [PubMed]
- Contro, E.; deSouza, R.; Bhide, A. Chronic Intervillositis of the Placenta: A Systematic Review. Placenta 2010, 31, 1106–1110. [Google Scholar] [CrossRef]
- Ferrer Montoya, R.; Robles Iglesias, A.; Pérez Dajaruch, M.d.l.A.; Crespo Bello, E.; González Vázquez, G. Corioamnionitis clínica e histológica: Revisión bibliográfica. MULTIMED 2014, 18. [Google Scholar]
- Mendoza, O.E.; Palomino, F.B.; Franco, N.R.; Guerra, D.R.; Niño, N.R. Corioamnionitis. Rev. Repert. Med. Y Cirugía 2013, 22, 248–256. [Google Scholar] [CrossRef]
- Jones, C.J.; Fox, H. Syncytial Knots and Intervillous Bridges in the Human Placenta: An Ultrastructural Study. J. Anat. 1977, 124, 275–286. [Google Scholar] [PubMed]
- Burton, G.J.; Jones, C.J.P. Syncytial Knots, Sprouts, Apoptosis, and Trophoblast Deportation from the Human Placenta. Taiwan. J. Obs. Gynecol. 2009, 48, 28–37. [Google Scholar] [CrossRef]
- Fogarty, N.M.E.; Ferguson-Smith, A.C.; Burton, G.J. Syncytial Knots (Tenney-Parker Changes) in the Human Placenta: Evidence of Loss of Transcriptional Activity and Oxidative Damage. Am. J. Pathol. 2013, 183, 144–152. [Google Scholar] [CrossRef]
- Natarajan, V.; Singh, P.; Vigneshwar, N.K.V.; Anthony, M.L.; Rao, S.; Chacham, S.; Chaturvedi, J.; Basu, S. Maternal and Placental Risk Factors for Small Gestational Age and Fetal Malnutrition. Curr. Pediatr. Rev. 2023, 19, 187–196. [Google Scholar] [CrossRef]
- Ramphal, S.; Govender, N.; Singh, S.; Khaliq, O.; Naicker, T. Histopathological Features in Advanced Abdominal Pregnancies Co-Infected with SARS-CoV-2 and HIV-1 Infections: A Case Evaluation. Eur. J. Obstet. Gynecol. Reprod. Biol. X 2022, 15, 100153. [Google Scholar] [CrossRef]
- Parks, W.T. Placental Hypoxia: The Lesions of Maternal Malperfusion. Semin. Perinatol. 2015, 39, 9–19. [Google Scholar] [CrossRef]
- Hladky, K.; Yankowitz, J.; Hansen, W.F. Placental Abruption. Obstet. Gynecol. Surv. 2002, 57, 299. [Google Scholar] [CrossRef]
- Tikkanen, M. Placental Abruption: Epidemiology, Risk Factors and Consequences. Acta Obstet. Gynecol. Scand. 2011, 90, 140–149. [Google Scholar] [CrossRef]
- Paessler, S.; Walker, D.H. Pathogenesis of the Viral Hemorrhagic Fevers. Annu. Rev. Pathol. Mech. Dis. 2013, 8, 411–440. [Google Scholar] [CrossRef]
- Scholz, C.; Hermann, C.; Kachler, A.; Kainer, F.; Friese, K.; Makrigiannakis, A.; Jeschke, U. Association of Placental Inflammation with Fetomaternal Hemorrhage and Loss of Placental Mucin-1. Arch. Gynecol. Obs. 2012, 285, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Beniwal, M.; Kar, P.; Sharma, J.B.; Murthy, N.S. Hepatitis E in Pregnancy. Int. J. Gynecol. Obstet. 2004, 85, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Brar, R.; Sikka, P.; Suri, V.; Singh, M.P.; Suri, V.; Mohindra, R.; Biswal, M. Maternal and Fetal Outcomes of Dengue Fever in Pregnancy: A Large Prospective and Descriptive Observational Study. Arch. Gynecol. Obs. 2021, 304, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, C.F.; Lopes, V.G.S.; Brasil, P.; Pires, A.R.C.; Rohloff, R.; Nogueira, R.M.R. Dengue Infection in Pregnancy and Its Impact on the Placenta. Int. J. Infect. Dis. 2017, 55, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, F.C.P.d.A.D.M.; da Silva, A.S.V.; Recht, J.; Guaraldo, L.; Moreira, M.E.L.; de Siqueira, A.M.; Gerardin, P.; Brasil, P. Vertical Transmission of Chikungunya Virus: A Systematic Review. PLoS ONE 2021, 16, e0249166. [Google Scholar] [CrossRef] [PubMed]
- Castejón Sandoval, O.C. The Placenta in a Case of Pregnant Woman Infected by Chikungunya Virus. J. Virol. Retrovirol. 2016, 2, 1–4. Available online: https://www.researchgate.net/publication/308516584_The_placenta_in_a_case_of_pregnant_woman_infected_by_Chikungunya_virus (accessed on 22 December 2023).
- Chaw, L.; Kamigaki, T.; Burmaa, A.; Urtnasan, C.; Od, I.; Nyamaa, G.; Nymadawa, P.; Oshitani, H. Burden of Influenza and Respiratory Syncytial Virus Infection in Pregnant Women and Infants Under 6 Months in Mongolia: A Prospective Cohort Study. PLoS ONE 2016, 11, e0148421. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.Y.; Katz, J.; Tielsch, J.; Khatry, S.K.; Shrestha, L.; LeClerq, S.C.; Magaret, A.; Kuypers, J.; Steinhoff, M.C.; Englund, J.A. Clinical Presentation and Birth Outcomes Associated with Respiratory Syncytial Virus Infection in Pregnancy. PLoS ONE 2016, 11, e0152015. [Google Scholar] [CrossRef]
- Bokun, V.; Moore, J.J.; Moore, R.; Smallcombe, C.C.; Harford, T.J.; Rezaee, F.; Esper, F.; Piedimonte, G. Respiratory Syncytial Virus Exhibits Differential Tropism for Distinct Human Placental Cell Types with Hofbauer Cells Acting as a Permissive Reservoir for Infection. PLoS ONE 2019, 14, e0225767. [Google Scholar] [CrossRef]
- Kantola, K.; Hedman, L.; Arthur, J.; Alibeto, A.; Delwart, E.; Jartti, T.; Ruuskanen, O.; Hedman, K.; Söderlund-Venermo, M. Seroepidemiology of Human Bocaviruses 1–4. J. Infect. Dis. 2011, 204, 1403–1412. [Google Scholar] [CrossRef]
- Schildgen, O.; Müller, A.; Allander, T.; Mackay, I.M.; Völz, S.; Kupfer, B.; Simon, A. Human Bocavirus: Passenger or Pathogen in Acute Respiratory Tract Infections? Clin. Microbiol. Rev. 2008, 21, 291–304. [Google Scholar] [CrossRef]
- Endo, R.; Ishiguro, N.; Kikuta, H.; Teramoto, S.; Shirkoohi, R.; Ma, X.; Ebihara, T.; Ishiko, H.; Ariga, T. Seroepidemiology of Human Bocavirus in Hokkaido Prefecture, Japan. J. Clin. Microbiol. 2007, 45, 3218–3223. [Google Scholar] [CrossRef] [PubMed]
- Lindner, J.; Karalar, L.; Zehentmeier, S.; Plentz, A.; Pfister, H.; Struff, W.; Kertai, M.; Segerer, H.; Modrow, S. Humoral Immune Response against Human Bocavirus VP2 Virus-like Particles. Viral Immunol. 2008, 21, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Riipinen, A.; Väisänen, E.; Lahtinen, A.; Karikoski, R.; Nuutila, M.; Surcel, H.-M.; Taskinen, H.; Hedman, K.; Söderlund-Venermo, M. Absence of Human Bocavirus from Deceased Fetuses and Their Mothers. J. Clin. Virol. 2010, 47, 186–188. [Google Scholar] [CrossRef]
- Dhama, K.; Malik, Y.S.; Malik, S.V.S.; Singh, R.K. Ebola from Emergence to Epidemic: The Virus and the Disease, Global Preparedness and Perspectives. J. Infect. Dev. Ctries. 2015, 9, 441–455. [Google Scholar] [CrossRef]
- Jones, R.M.; Brosseau, L.M.; Nov, S. COMMENTARY: Ebola Virus Transmission via Contact and Aerosol—A New Paradigm; CIDRAP: Minneapolis, MN, USA, 2014; Available online: https://www.cidrap.umn.edu/ebola/commentary-ebola-virus-transmission-contact-and-aerosol-new-paradigm (accessed on 29 December 2023).
- Schwartz, D.A. Maternal Filovirus Infection and Death from Marburg and Ravn Viruses: Highly Lethal to Pregnant Women and Their Fetuses Similar to Ebola Virus. In Emerging Challenges in Filovirus Infections; IntechOpen: London, UK, 2019; Available online: https://www.intechopen.com/chapters/68376 (accessed on 29 December 2023).
- Olgun, N.S. Viral Infections in Pregnancy: A Focus on Ebola Virus. Curr. Pharm. Des. 2018, 24, 993–998. [Google Scholar] [CrossRef] [PubMed]
- Coler, B.; Cervantes, O.; Li, M.; Coler, C.; Li, A.; Shivakumar, M.; Every, E.; Schwartz, D.; Adams Waldorf, K.M. Common Pathways Targeted by Viral Hemorrhagic Fever Viruses to Infect the Placenta and Increase the Risk of Stillbirth. Placenta 2023, 141, 2–9. [Google Scholar] [CrossRef]
- Richmond, J.K.; Baglole, D.J. Lassa Fever: Epidemiology, Clinical Features, and Social Consequences. BMJ 2003, 327, 1271–1275. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Aziz, A.A.; Meegan, J.M.; Laughlin, L.W. Rift Valley Fever as a Possible Cause of Human Abortions. Trans. R. Soc. Trop. Med. Hyg. 1980, 74, 685–686. [Google Scholar] [CrossRef]
- Adam, I.; Karsany, M.S. Case Report: Rift Valley Fever with Vertical Transmission in a Pregnant Sudanese Woman. J. Med. Virol. 2008, 80, 929. [Google Scholar] [CrossRef] [PubMed]
- Baudin, M.; Jumaa, A.M.; Jomma, H.J.E.; Karsany, M.S.; Bucht, G.; Näslund, J.; Ahlm, C.; Evander, M.; Mohamed, N. Association of Rift Valley Fever Virus Infection with Miscarriage in Sudanese Women: A Cross-Sectional Study. Lancet Glob. Health 2016, 4, e864–e871. [Google Scholar] [CrossRef] [PubMed]
- Assiri, A.; Abedi, G.R.; Al Masri, M.; Bin Saeed, A.; Gerber, S.I.; Watson, J.T. Middle East Respiratory Syndrome Coronavirus Infection During Pregnancy: A Report of 5 Cases from Saudi Arabia. Clin. Infect. Dis. 2016, 63, 951–953. [Google Scholar] [CrossRef]
- Malik, A.; El Masry, K.M.; Ravi, M.; Sayed, F. Middle East Respiratory Syndrome Coronavirus during Pregnancy, Abu Dhabi, United Arab Emirates, 2013. Emerg. Infect. Dis. 2016, 22, 515–517. [Google Scholar] [CrossRef]
- Jeong, S.Y.; Sung, S.I.; Sung, J.-H.; Ahn, S.Y.; Kang, E.-S.; Chang, Y.S.; Park, W.S.; Kim, J.-H. MERS-CoV Infection in a Pregnant Woman in Korea. J. Korean Med. Sci. 2017, 32, 1717–1720. [Google Scholar] [CrossRef]
- Schwartz, D.A.; Graham, A.L. Potential Maternal and Infant Outcomes from Coronavirus 2019-nCoV (SARS-CoV-2) Infecting Pregnant Women: Lessons from SARS, MERS, and Other Human Coronavirus Infections. Viruses 2020, 12, 194. [Google Scholar] [CrossRef] [PubMed]
- Ng, P.C.; So, K.W.; Leung, T.F.; Cheng, F.W.T.; Lyon, D.J.; Wong, W.; Cheung, K.L.; Fung, K.S.C.; Lee, C.H.; Li, A.M.; et al. Infection Control for SARS in a Tertiary Neonatal Centre. Arch. Dis. Child. Fetal Neonatal Ed. 2003, 88, F405–F409. [Google Scholar] [CrossRef]
- Wong, S.F.; Chow, K.M.; de Swiet, M. Severe Acute Respiratory Syndrome and Pregnancy. BJOG 2003, 110, 641–642. [Google Scholar] [CrossRef]
- Ng, W.F.; Wong, S.F.; Lam, A.; Mak, Y.F.; Yao, H.; Lee, K.C.; Chow, K.M.; Yu, W.C.; Ho, L.C. The Placentas of Patients with Severe Acute Respiratory Syndrome: A Pathophysiological Evaluation. Pathology 2006, 38, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Chamseddine, R.S.; Wahbeh, F.; Chervenak, F.; Salomon, L.J.; Ahmed, B.; Rafii, A. Pregnancy and Neonatal Outcomes in SARS-CoV-2 Infection: A Systematic Review. J. Pregnancy 2020, 2020, e4592450. [Google Scholar] [CrossRef]
- McClymont, E.; Albert, A.Y.; Alton, G.D.; Boucoiran, I.; Castillo, E.; Fell, D.B.; Kuret, V.; Poliquin, V.; Reeve, T.; Scott, H.; et al. Association of SARS-CoV-2 Infection During Pregnancy with Maternal and Perinatal Outcomes. JAMA 2022, 327, 1983–1991. [Google Scholar] [CrossRef] [PubMed]
- Timircan, M.; Bratosin, F.; Vidican, I.; Suciu, O.; Tirnea, L.; Avram, V.; Marincu, I. Exploring Pregnancy Outcomes Associated with SARS-CoV-2 Infection. Medicina 2021, 57, 796. [Google Scholar] [CrossRef]
- Baergen, R.N.; Heller, D.S. Placental Pathology in COVID-19 Positive Mothers: Preliminary Findings. Pediatr. Dev. Pathol. 2020, 23, 177–180. Available online: https://journals.sagepub.com/doi/full/10.1177/1093526620925569 (accessed on 22 December 2023). [CrossRef]
- Singh, N.; Buckley, T.; Shertz, W. Placental Pathology in COVID-19: Case Series in a Community Hospital Setting. Cureus 2021, 13, e12522. [Google Scholar] [CrossRef]
- Kouba, I.; Bracero, L.; Millington, K.; Blitz, M.J. Placental Calcifications after Coronavirus Disease 2019 in First Trimester of Pregnancy: Ultrasound and Pathology Findings. Med. Ultrason. 2023, 25, 233–235. [Google Scholar] [CrossRef]
- Knegendorf, L.; Drave, S.A.; Dao Thi, V.L.; Debing, Y.; Brown, R.J.P.; Vondran, F.W.R.; Resner, K.; Friesland, M.; Khera, T.; Engelmann, M.; et al. Hepatitis E Virus Replication and Interferon Responses in Human Placental Cells. Hepatol. Commun. 2018, 2, 173. [Google Scholar] [CrossRef]
- Zhao, J.; Huang, X.; Mcleod, P.; Jiang, J.; Liu, W.; Haig, A.; Jevnikar, A.M.; Jiang, Z.; Zhang, Z.-X. Toll-like Receptor 3 Is an Endogenous Sensor of Cell Death and a Potential Target for Induction of Long-Term Cardiac Transplant Survival. Am. J. Transplant. 2021, 21, 3268–3279. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Zhang, Y.; Lv, Q.; Liu, B.; Jin, M.; Zhang, W.; He, Q.; Deng, M.; Liu, X.; Li, G.; et al. Toll-like Receptor 3 (TLR3) Induces Apoptosis via Death Receptors and Mitochondria by Up-Regulating the Transactivating P63 Isoform α (TAP63α). J. Biol. Chem. 2011, 286, 15918–15928. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Vasudevan, S.G. Clinical and Experimental Evidence for Transplacental Vertical Transmission of Flaviviruses. Antivir. Res. 2023, 210, 105512. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Chan, K.W.K.; Tan, N.W.W.; Mahid, M.B.A.; Chowdhury, A.; Chang, K.T.E.; Vasudevan, S.G. Experimental Evidence for a High Rate of Maternal-Fetal Transmission of Dengue Virus in the Presence of Antibodies in Immunocompromised Mice. EBioMedicine 2022, 77, 103930. [Google Scholar] [CrossRef] [PubMed]
- Castillo Ramirez, J.A.; Urcuqui-Inchima, S. Dengue Virus Control of Type I IFN Responses: A History of Manipulation and Control. J. Interferon Cytokine Res. 2015, 35, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Morrison, J.; Aguirre, S.; Fernandez-Sesma, A. Innate Immunity Evasion by Dengue Virus. Viruses 2012, 4, 397–413. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, S.; Fernandez-Sesma, A. Collateral Damage during Dengue Virus Infection: Making Sense of DNA by cGAS. J. Virol. 2017, 91, e01081-16. Available online: https://journals.asm.org/doi/full/10.1128/jvi.01081-16 (accessed on 3 January 2024). [CrossRef]
- Viettri, M.; Caraballo, G.; Sanchez, M.E.; Espejel-Nuñez, A.; Betanzos, A.; Ortiz-Navarrete, V.; Estrada-Gutierrez, G.; Nava, P.; Ludert, J.E. Comparative Infections of Zika, Dengue, and Yellow Fever Viruses in Human Cytotrophoblast-Derived Cells Suggest a Gating Role for the Cytotrophoblast in Zika Virus Placental Invasion. Microbiol. Spectr. 2023, 11, e00630-23. [Google Scholar] [CrossRef]
- Gérardin, P.; Couderc, T.; Bintner, M.; Tournebize, P.; Renouil, M.; Lémant, J.; Boisson, V.; Borgherini, G.; Staikowsky, F.; Schramm, F.; et al. Chikungunya Virus–Associated Encephalitis. Neurology 2016, 86, 94–102. [Google Scholar] [CrossRef]
- Ramful, D.; Carbonnier, M.; Pasquet, M.; Bouhmani, B.; Ghazouani, J.; Noormahomed, T.; Beullier, G.; Attali, T.; Samperiz, S.; Fourmaintraux, A.; et al. Mother-to-Child Transmission of Chikungunya Virus Infection. Pediatr. Infect. Dis. J. 2007, 26, 811–815. [Google Scholar] [CrossRef]
- Salomão, N.; Brendolin, M.; Rabelo, K.; Wakimoto, M.; de Filippis, A.M.; Dos Santos, F.; Moreira, M.E.; Basílio-de-Oliveira, C.A.; Avvad-Portari, E.; Paes, M.; et al. Spontaneous Abortion and Chikungunya Infection: Pathological Findings. Viruses 2021, 13, 554. [Google Scholar] [CrossRef]
- Bidzhekov, K.; Zernecke, A.; Weber, C. MCP-1 Induces a Novel Transcription Factor with Proapoptotic Activity. Circ. Res. 2006, 98, 1107–1109. [Google Scholar] [CrossRef] [PubMed]
- Velázquez-Cervantes, M.A.; Martínez-Castillo, M.; González-García, L.D.; Vargas-Pavía, T.A.; Martínez-Salazar, M.G.; Mancilla-Herrera, I.; León-Reyes, G.; García-Cordero, J.; Helguera-Repetto, A.C.; León-Juárez, M. The BeWo Cell Line Derived from a Human Placental Choriocarcinoma Is Permissive for Respiratory Syncytial Virus Infection. Virus Genes 2019, 55, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Arnold, R.; König, W. Respiratory Syncytial Virus Infection of Human Lung Endothelial Cells Enhances Selectively Intercellular Adhesion Molecule-1 Expression. J. Immunol. 2005, 174, 7359–7367. [Google Scholar] [CrossRef] [PubMed]
- Carette, J.E.; Raaben, M.; Wong, A.C.; Herbert, A.S.; Obernosterer, G.; Mulherkar, N.; Kuehne, A.I.; Kranzusch, P.J.; Griffin, A.M.; Ruthel, G.; et al. Ebola Virus Entry Requires the Cholesterol Transporter Niemann-Pick C1. Nature 2011, 477, 340–343. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Chen, L.; Zhang, J.; Xiong, C.; Li, X. The SARS-CoV-2 Receptor ACE2 Expression of Maternal-Fetal Interface and Fetal Organs by Single-Cell Transcriptome Study. PLoS ONE 2020, 15, e0230295. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Pontnou, M.; Navarro, A.; Camacho, J.; Crispi, F.; Alguacil-Guillén, M.; Moreno-Baró, A.; Hernandez-Losa, J.; Sesé, M.; Ramón y Cajal, S.; Garcia Ruíz, I.; et al. Diffuse Trophoblast Damage Is the Hallmark of SARS-CoV-2-Associated Fetal Demise. Mod. Pathol. 2021, 34, 1704–1709. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Hua, D.; Li, J.-P.; Zhang, X.-N.; Bai, L.; Cao, L.-B.; Guo, Y.; Zhang, M.; Dong, J.-Z.; Liang, X.-W.; et al. Modulation of Innate Immune Response to Viruses Including SARS-CoV-2 by Progesterone. Sig Transduct. Target. Ther. 2022, 7, 137. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.-C.; Uhl, S.; Hoagland, D.; Møller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 2020, 181, 1036–1045.e9. [Google Scholar] [CrossRef]
- Lockwood, C.J.; Murk, W.K.; Kayisli, U.A.; Buchwalder, L.F.; Huang, S.J.; Arcuri, F.; Li, M.; Gopinath, A.; Schatz, F. Regulation of Interleukin-6 Expression in Human Decidual Cells and Its Potential Role in Chorioamnionitis. Am. J. Pathol. 2010, 177, 1755–1764. [Google Scholar] [CrossRef]
- Guzeloglu-Kayisli, O.; Kayisli, U.A.; Semerci, N.; Basar, M.; Buchwalder, L.F.; Buhimschi, C.S.; Buhimschi, I.A.; Arcuri, F.; Larsen, K.; Huang, J.S.; et al. Mechanisms of Chorioamnionitis-Associated Preterm Birth: Interleukin-1β Inhibits Progesterone Receptor Expression in Decidual Cells. J. Pathol. 2015, 237, 423–434. [Google Scholar] [CrossRef]
- Holland, O.; Dekker Nitert, M.; Gallo, L.A.; Vejzovic, M.; Fisher, J.J.; Perkins, A.V. Review: Placental Mitochondrial Function and Structure in Gestational Disorders. Placenta 2017, 54, 2–9. [Google Scholar] [CrossRef]
- Hastie, R.; Lappas, M. The Effect of Pre-Existing Maternal Obesity and Diabetes on Placental Mitochondrial Content and Electron Transport Chain Activity. Placenta 2014, 35, 673–683. [Google Scholar] [CrossRef]
- Sies, H. Oxidative Stress: A Concept in Redox Biology and Medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef]
- Lee, H.-C.; Wei, Y.-H. Mitochondrial Biogenesis and Mitochondrial DNA Maintenance of Mammalian Cells under Oxidative Stress. Int. J. Biochem. Cell Biol. 2005, 37, 822–834. [Google Scholar] [CrossRef]
- Mandò, C.; Savasi, V.M.; Anelli, G.M.; Corti, S.; Serati, A.; Lisso, F.; Tasca, C.; Novielli, C.; Cetin, I. Mitochondrial and Oxidative Unbalance in Placentas from Mothers with SARS-CoV-2 Infection. Antioxidants 2021, 10, 1517. [Google Scholar] [CrossRef] [PubMed]
- South, K.; Lane, D.A. ADAMTS-13 and von Willebrand Factor: A Dynamic Duo. J. Thromb. Haemost. 2018, 16, 6–18. [Google Scholar] [CrossRef] [PubMed]
- Gory-Fauré, S.; Prandini, M.H.; Pointu, H.; Roullot, V.; Pignot-Paintrand, I.; Vernet, M.; Huber, P. Role of Vascular Endothelial-Cadherin in Vascular Morphogenesis. Development 1999, 126, 2093–2102. [Google Scholar] [CrossRef] [PubMed]
- Hromatka, B.S.; Drake, P.M.; Kapidzic, M.; Stolp, H.; Goldfien, G.A.; Shih, I.-M.; Fisher, S.J. Polysialic Acid Enhances the Migration and Invasion of Human Cytotrophoblasts. Glycobiology 2013, 23, 593–602. [Google Scholar] [CrossRef]
- Sukhikh, G.T.; Ziganshina, M.M.; Nizyaeva, N.V.; Kulikova, G.V.; Volkova, J.S.; Yarotskaya, E.L.; Kan, N.E.; Shchyogolev, A.I.; Tyutyunnik, V.L. Differences of Glycocalyx Composition in the Structural Elements of Placenta in Preeclampsia. Placenta 2016, 43, 69–76. [Google Scholar] [CrossRef]
- Perna, A.; Tani, A.; Sellitto, C.; Marini, M.; La Verde, M.; De Luca, A.; Guerra, G.; Lucariello, A.; Manetti, M.; Sgambati, E. Sialylation Status in Placentas from Pregnancies with SARS-CoV-2 Infection. Tissue Cell 2023, 82, 102074. [Google Scholar] [CrossRef]
- Pillay, P.; Moodley, K.; Vatish, M.; Moodley, J.; Duarte, R.; Mackraj, I. Exosomal Th1/Th2 Cytokines in Preeclampsia and HIV-Positive Preeclamptic Women on Highly Active Anti-Retroviral Therapy. Cytokine 2020, 125, 154795. [Google Scholar] [CrossRef]
- Sheller-Miller, S.; Urrabaz-Garza, R.; Saade, G.; Menon, R. Damage-Associated Molecular Pattern Markers HMGB1 and Cell-Free Fetal Telomere Fragments in Oxidative-Stressed Amnion Epithelial Cell-Derived Exosomes. J. Reprod. Immunol. 2017, 123, 3–11. [Google Scholar] [CrossRef]
- Kammala, A.K.; Mosebarger, A.; Radnaa, E.; Rowlinson, E.; Vora, N.; Fortunato, S.J.; Sharma, S.; Safarzadeh, M.; Menon, R. Extracellular Vesicles-Mediated Recombinant IL-10 Protects against Ascending Infection-Associated Preterm Birth by Reducing Fetal Inflammatory Response. Front. Immunol. 2023, 14, 1196453. [Google Scholar] [CrossRef]
- Zhao, Y.; Zheng, Y.; Liu, X.; Luo, Q.; Wu, D.; Liu, X.; Zou, L. Inhibiting Trophoblast PAR-1 Overexpression Suppresses sFlt-1-Induced Anti-Angiogenesis and Abnormal Vascular Remodeling: A Possible Therapeutic Approach for Preeclampsia. Mol. Hum. Reprod. 2018, 24, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Galán, A.; Fernández-Messina, L.; Sánchez-Madrid, F. Control of Immunoregulatory Molecules by miRNAs in T Cell Activation. Front. Immunol. 2018, 9, 2148. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Xue, M.; Xu, P.; Hu, F.; Sun, B.; Xiao, Z. MicroRNA Profiling Analysis Revealed Different Cellular Senescence Mechanisms in Human Mesenchymal Stem Cells Derived from Different Origin. Genomics 2017, 109, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, C.; Zhang, C.; Shang, R.; Zhang, Y.; Wu, S.; Long, Y. LncRNA Small Nucleolar RNA Host Gene 5 Inhibits Trophoblast Autophagy in Preeclampsia by Targeting microRNA-31-5p and Promoting the Transcription of Secreted Protein Acidic and Rich in Cysteine. Bioengineered 2022, 13, 7221–7237. [Google Scholar] [CrossRef] [PubMed]
- Motta, J.-P.; Magne, L.; Descamps, D.; Rolland, C.; Squarzoni-Dale, C.; Rousset, P.; Martin, L.; Cenac, N.; Balloy, V.; Huerre, M.; et al. Modifying the Protease, Antiprotease Pattern by Elafin Overexpression Protects Mice from Colitis. Gastroenterology 2011, 140, 1272–1282. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, S.H.; Hui, C.C.; Cheah, A.Y.; You, X.M.; Husain, M.; Rabinovitch, M. Targeted Overexpression of Elafin Protects Mice against Cardiac Dysfunction and Mortality Following Viral Myocarditis. J. Clin. Investig. 1999, 103, 1211–1219. [Google Scholar] [CrossRef] [PubMed]
- Drannik, A.G.; Nag, K.; Sallenave, J.-M.; Rosenthal, K.L. Antiviral Activity of Trappin-2 and Elafin in Vitro and in Vivo against Genital Herpes. J. Virol. 2013, 87, 7526–7538. [Google Scholar] [CrossRef]
- Taggart, C.C.; Cryan, S.-A.; Weldon, S.; Gibbons, A.; Greene, C.M.; Kelly, E.; Low, T.B.; O’neill, S.J.; McElvaney, N.G. Secretory Leucoprotease Inhibitor Binds to NF-kappaB Binding Sites in Monocytes and Inhibits P65 Binding. J. Exp. Med. 2005, 202, 1659–1668. [Google Scholar] [CrossRef]
- Drannik, A.G.; Nag, K.; Yao, X.-D.; Henrick, B.M.; Sallenave, J.-M.; Rosenthal, K.L. Trappin-2/Elafin Modulate Innate Immune Responses of Human Endometrial Epithelial Cells to PolyI:C. PLoS ONE 2012, 7, e35866. [Google Scholar] [CrossRef]
- Butler, M.W.; Robertson, I.; Greene, C.M.; O’Neill, S.J.; Taggart, C.C.; McElvaney, N.G. Elafin Prevents Lipopolysaccharide-Induced AP-1 and NF-kappaB Activation via an Effect on the Ubiquitin-Proteasome Pathway. J. Biol. Chem. 2006, 281, 34730–34735. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Trimester | Effects on Placenta | References |
|---|---|---|---|
| CHIKV | 3 | Induces the release of proinflammatory molecules and alteration in the placental structure, absence of the syncytiotrophoblast layer, interruption in the placental surface, fibrosis in the stromal region, calcification, mitochondrial and endoplasmic reticulum alterations in trophoblasts, subtrophoblastic edema and cell death. | [16] |
| HCMV | 1,2,3 | In trophoblasts, modifies the proliferative, migratory, and invasive properties necessary for this implantation. | [21,22,23] |
| HSV | 1,2,3 | Disturbs the transport of antigens via HLA-G, affecting the tolerance of immune cells and thereby generating mechanisms of damage to the maternal–fetal interface. | [27,28] |
| HPV | 1 | Promotes an apoptotic effect, implantation, early embryonic development and miscarriage. | [31,32,33,34] |
| LASV | 3 | Regulation of interferon response. | [40] |
| SARS-CoV-2 | 1 | Poor maternal vascular malperfusion, increased fibrin deposition, villous agglutination, intervillous thrombi, atherosis and increased syncytial knots, calcification, manifest placental damage associated with inflammation, manifesting intervillitis, with high infiltration of neutrophils and Hofbauer cells in the chorionic villi, as well as cell death. | [50,51,52,53] |
| HEV | 3 | Calcification, fibrosis, and great tissue inflammation, membrane ruptures, hemorrhages, expression of proinflammatory cytokines and high infiltration of immune system cells. | [54,55,56,57] |
| DENV | 1 | Induces the release of proinflammatory molecules, inhibit the activation of the cGAS/STING pathway, decrease the activation of the mTOR pathway related to cellular processes such as growth and differentiation. | [58,59] |
| RSV | 3 | Alterations in the membranes of syncytiotrophoblasts and placental damage due to inflammation. | [60,61,62] |
| HBoV | 1,2,3 | Spontaneous abortions. | [63] |
| EBOV | 1,2,3 | Expression of proinflammatory cytokines, permeability of endothelial cells and hemorrhages in the placenta. | [64,65] |
| Changes in the Placenta | Virus | References |
|---|---|---|
| 1. Fibrin deposits | Adenovirus, Coxsackievirus–B, SARS-CoV-2 | [51,52,66] |
| 2. Placental calcification | HEV, CHIKV, SARS-CoV-2 | [58,67,68] |
| 3. Placental inflammation (Intervillositis and Chorioamnionitis) | HEV, DENV, CHIKV, SARS-CoV-1, SARS-CoV-2, RSV, EBOV | [16,52,59,66,69,70,71,72] |
| 4. Syncytial knots | RSV, SARS-CoV-2 | [53,73,74] |
| 5. Maternal vascular malperfusion | SARS-CoV1, SARS-CoV-2 | [72,75,76] |
| 6. Placental abruption | HBV, SARS-CoV-2 | [77,78] |
| 7. Hemorrhages | HEV, DENV, EBOV, MARV, LASV, RVF | [58,70,79,80] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruz-Holguín, V.J.; González-García, L.D.; Velázquez-Cervantes, M.A.; Arévalo-Romero, H.; De Jesús-González, L.A.; Helguera-Repetto, A.C.; León-Reyes, G.; Salazar, M.I.; Cedillo-Barrón, L.; León-Juárez, M. Collateral Damage in the Placenta during Viral Infection in Pregnancy: A Possible Mechanism for Vertical Transmission and an Adverse Pregnancy Outcome. Diseases 2024, 12, 59. https://doi.org/10.3390/diseases12030059
Cruz-Holguín VJ, González-García LD, Velázquez-Cervantes MA, Arévalo-Romero H, De Jesús-González LA, Helguera-Repetto AC, León-Reyes G, Salazar MI, Cedillo-Barrón L, León-Juárez M. Collateral Damage in the Placenta during Viral Infection in Pregnancy: A Possible Mechanism for Vertical Transmission and an Adverse Pregnancy Outcome. Diseases. 2024; 12(3):59. https://doi.org/10.3390/diseases12030059
Chicago/Turabian StyleCruz-Holguín, Victor Javier, Luis Didier González-García, Manuel Adrián Velázquez-Cervantes, Haruki Arévalo-Romero, Luis Adrián De Jesús-González, Addy Cecilia Helguera-Repetto, Guadalupe León-Reyes, Ma. Isabel Salazar, Leticia Cedillo-Barrón, and Moisés León-Juárez. 2024. "Collateral Damage in the Placenta during Viral Infection in Pregnancy: A Possible Mechanism for Vertical Transmission and an Adverse Pregnancy Outcome" Diseases 12, no. 3: 59. https://doi.org/10.3390/diseases12030059