
The Mechanism of Orientation Detection Based on Artificial Visual System



Abstract
:1. Introduction
2. Mechanism
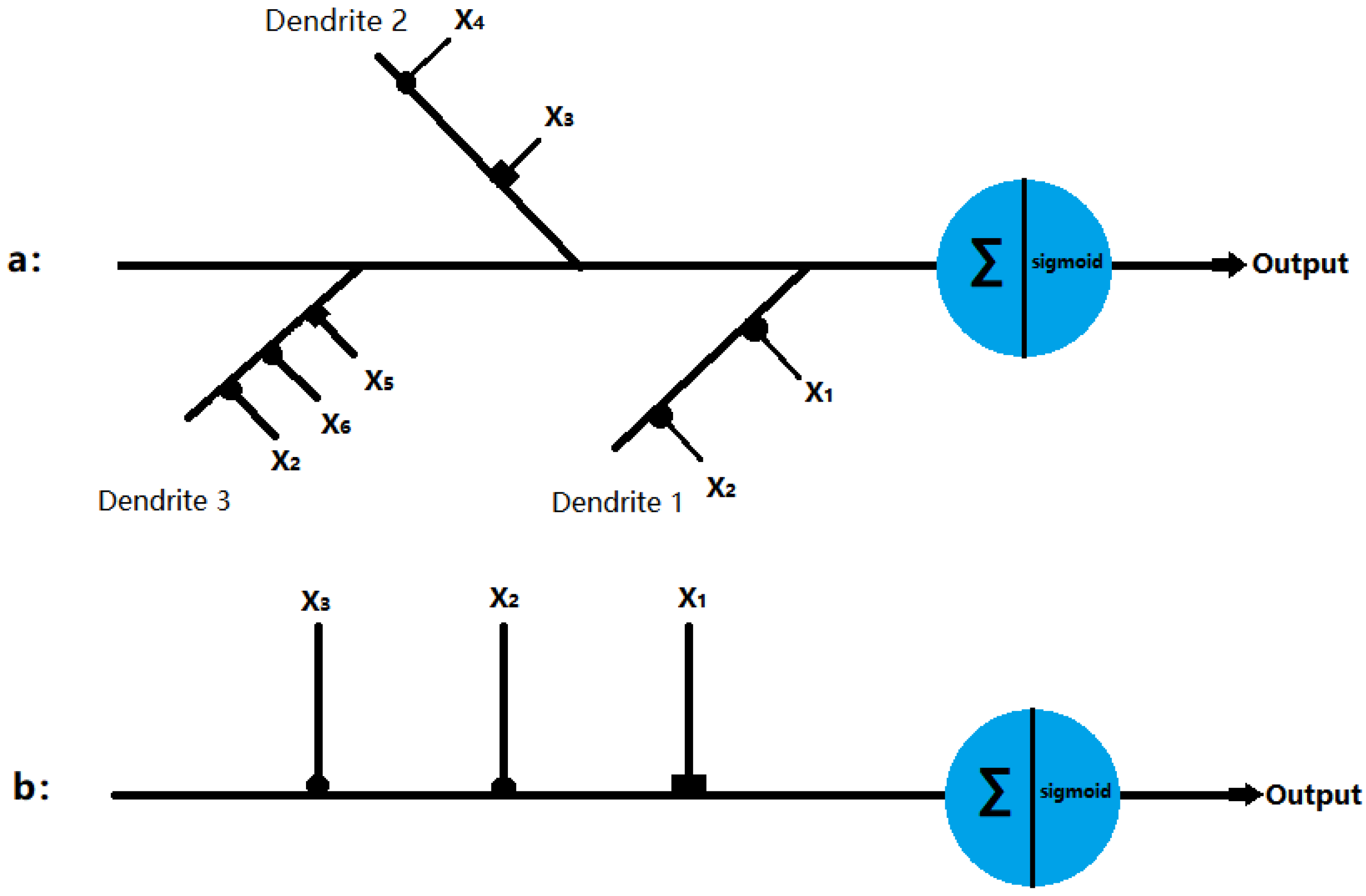
2.1. Dendritic Neuron Model
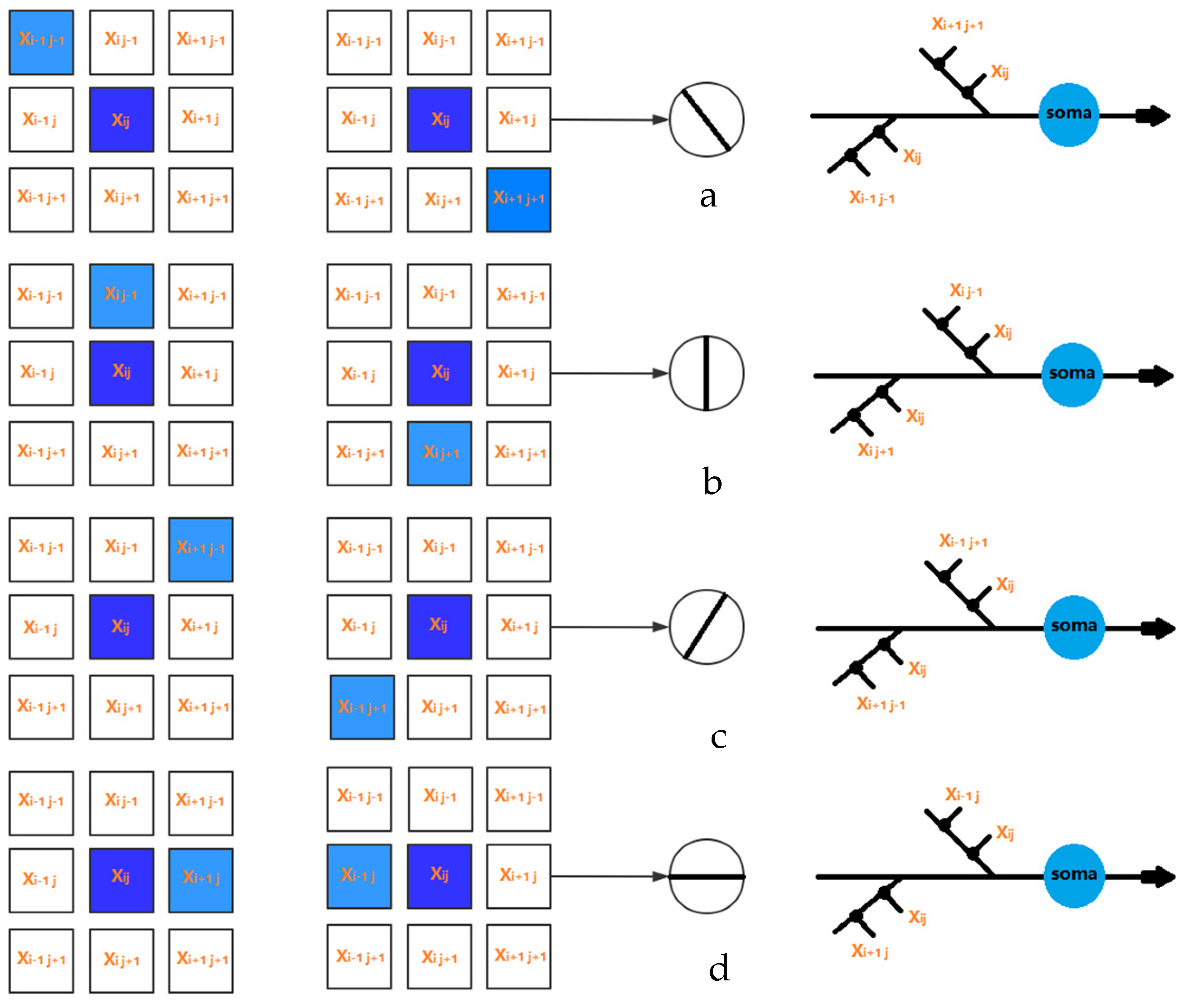
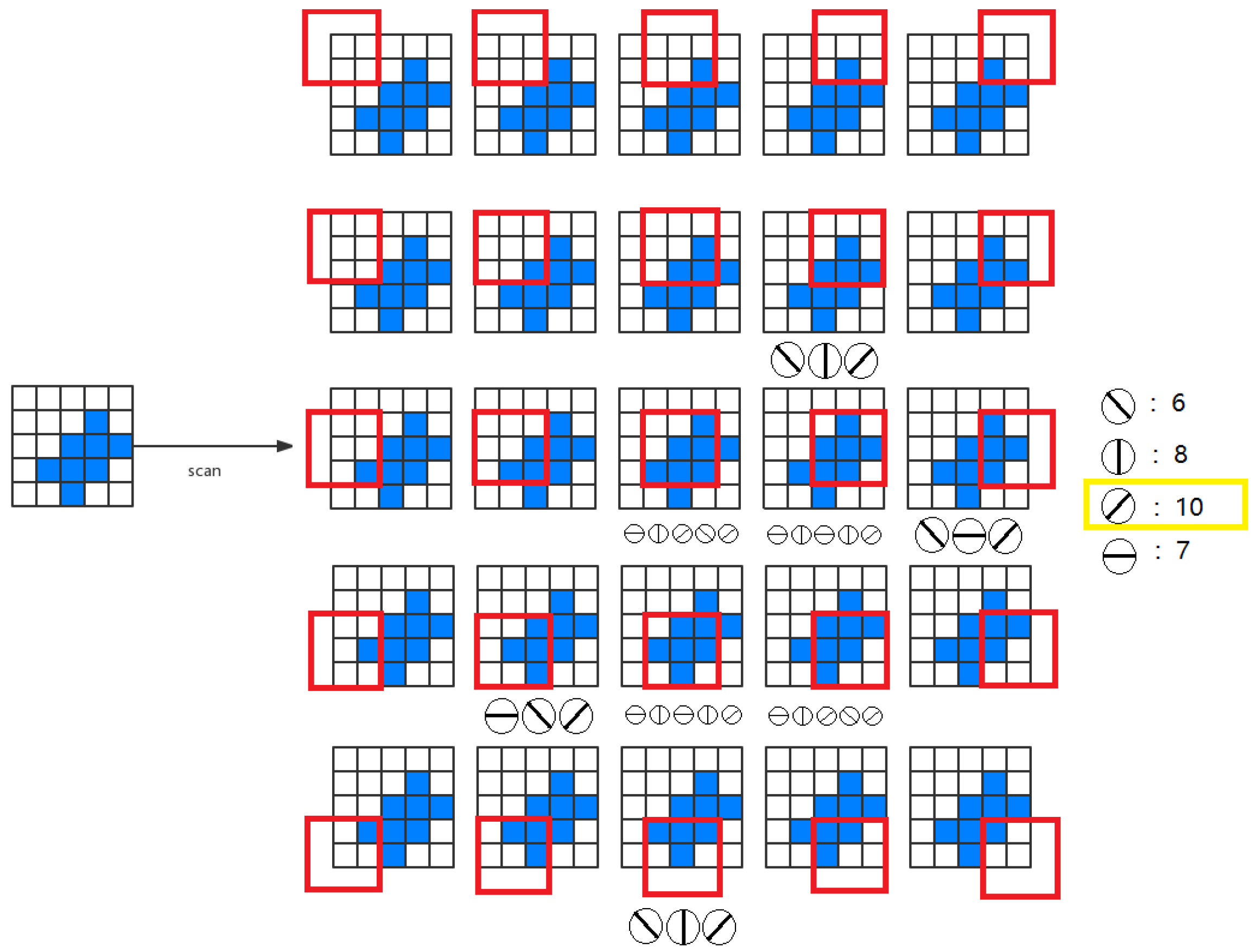
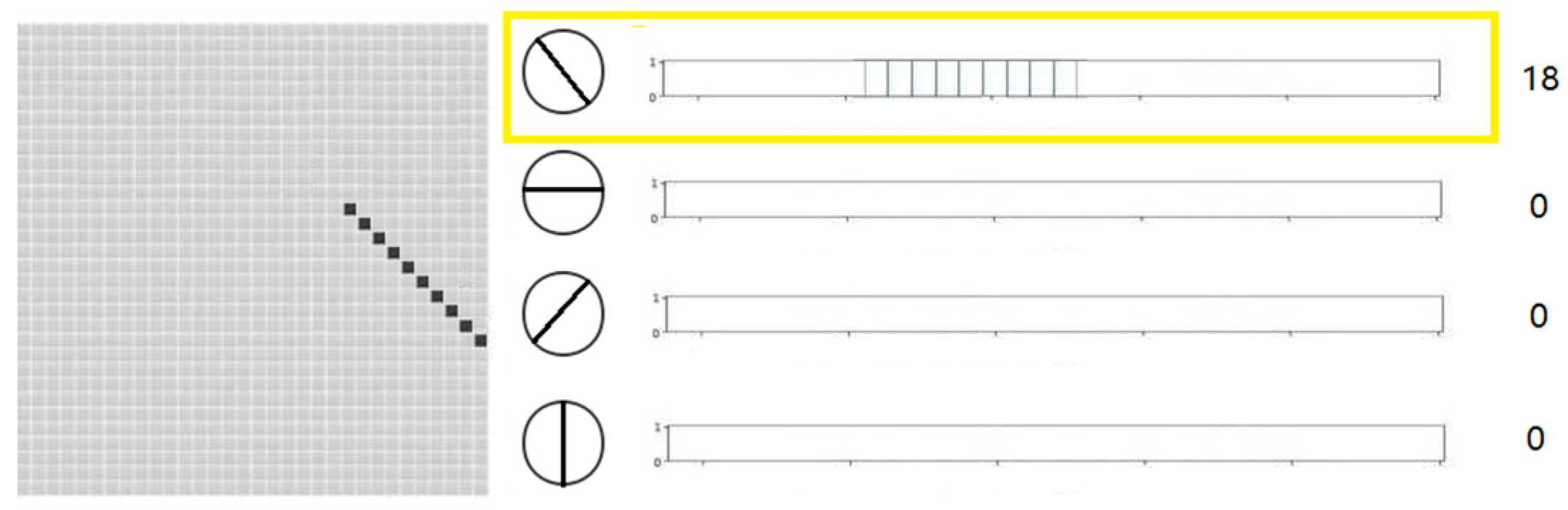
2.2. Local Orientation Detection Neuron
2.3. Global Orientation Detection
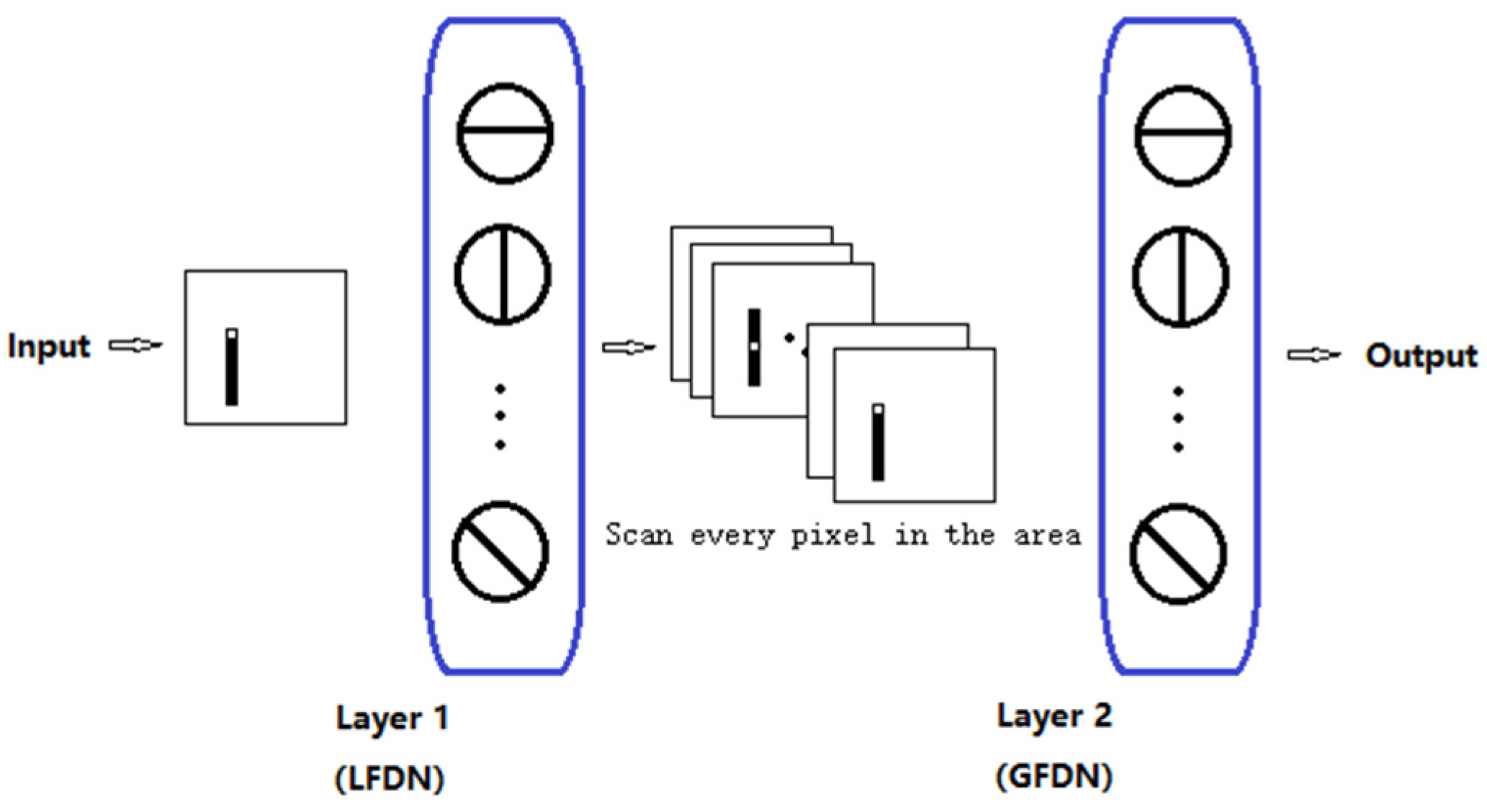
2.4. Artificial Visual System (AVS)
3. Experiment
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hubel, D.H.; Wiesel, T.N. Receptive fields, binocular interaction and functional architecture in the cat’s visual cortex. J. Physiol. 1962, 160, 106–154. [Google Scholar] [CrossRef] [PubMed]
- Hubel, D.H.; Wiesel, T.N. Receptive fields of single neurones in the cat’s striate cortex. J. Physiol. 1959, 148, 574–591. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, C.D.; Li, W. Adult visual cortical plasticity. Neuron 2012, 75, 250–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubel, D.H.; Wiesel, T.N. Exploration of the primary visual cortex, 1955–1978. Nature 1982, 299, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Hubel, D.H.; Wiesel, T.N. Receptive fields and functional architecture of monkey striate cortex. J. Physiol. 1968, 195, 215–243. [Google Scholar] [CrossRef]
- Jin, J.; Wang, Y.; Swadlow, H.A.; Alonso, J.M. Population receptive fields of ON and OFF thalamic inputs to an orientation column in visual cortex. Nat. Neurosci. 2011, 14, 232–238. [Google Scholar] [CrossRef]
- Priebe, N.J.; Ferster, D. Mechanisms of neuronal computation in mammalian visual cortex. Neuron 2012, 75, 194–208. [Google Scholar] [CrossRef] [Green Version]
- Wilson, D.E.; Whitney, D.E.; Scholl, B.; Fitzpatrick, D. Orientation selectivity and the functional clustering of synaptic inputs in primary visual cortex. Nat. Neurosci. 2016, 19, 1003–1009. [Google Scholar] [CrossRef]
- McLaughlin, D.; Shapley, R.; Shelley, M.; Wielaard, D.J. A neuronal network model of macaque primary visual cortex (v1): Orientation selectivity and dynamics in the input layer 4cα. Proc. Natl. Acad. Sci. USA 2000, 97, 8087–8092. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Tonegawa, S. Molecular genetic analysis of synaptic plasticity, activity-dependent neural development, learning, and memory in the mammalian brain. Annu. Rev. Neurosci. 1997, 20, 157–184. [Google Scholar] [CrossRef] [Green Version]
- Golding, N.L.; Spruston, N. Dendritic sodium spikes are variable triggers of axonal action potentials in hippocampal CA1 pyramidal neurons. Neuron 1998, 21, 1189–1200. [Google Scholar] [CrossRef] [Green Version]
- Häusser, M.; Spruston, N.; Stuart, G.J. Diversity and dynamics of dendritic signaling. Science 2000, 290, 739–744. [Google Scholar] [CrossRef] [Green Version]
- Martina, M.; Vida, I.; Jonas, P. Distal initiation and active propagation of action potentials in interneuron dendrites. Science 2000, 287, 295–300. [Google Scholar] [CrossRef] [Green Version]
- Schwindt, P.C.; Crill, W.E. Local and propagated dendritic action potentials evoked by glutamate iontophoresis on rat neocortical pyramidal neurons. J. Neurophysiol. 1997, 77, 2466–2483. [Google Scholar] [CrossRef] [PubMed]
- Stuart, G.; Spruston, N.; Sakmann, B.; Häusser, M. Action potential initiation and backpropagation in neurons of the mammalian CNS. Trends Neurosci. 1997, 20, 125–131. [Google Scholar] [CrossRef]
- Velte, T.J.; Masland, R.H. Action potentials in the dendrites of retinal ganglion cells. J. Neurophysiol. 1999, 81, 1412–1417. [Google Scholar] [CrossRef] [Green Version]
- Taylor, W.; Vaney, D.I. New directions in retinal research. Trends Neurosci. 2003, 26, 379–385. [Google Scholar] [CrossRef]
- Fried, S.I.; Münch, T.A.; Werblin, F.S. Mechanisms and circuitry underlying directional selectivity in the retina. Nature 2002, 420, 411–414. [Google Scholar] [CrossRef]
- Oesch, N.; Euler, T.; Taylor, W.R. Direction-selective dendritic action potentials in rabbit retina. Neuron 2005, 47, 739–750. [Google Scholar] [CrossRef] [Green Version]
- Koch, C. Biophysics of Computation: Information Processing in Single Neurons; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Silver, R.A. Neuronal arithmetic. Nat. Rev. Neurosci. 2010, 11, 474–489. [Google Scholar] [CrossRef] [Green Version]
- Todo, Y.; Tamura, H.; Yamashita, K.; Tang, Z. Unsupervised learnable neuron model with nonlinear interaction on dendrites. Neural Netw. 2014, 60, 96–103. [Google Scholar] [CrossRef]
- Hebb, D.O. The Organization of Behavior: A Neuropsychological Theory; John Wiley and Sons: Hoboken, NJ, USA, 1949. [Google Scholar]
- Gidon, A.; Zolnik, T.A.; Fidzinski, P.; Bolduan, F.; Papoutsi, A.; Poirazi, P.; Holtkamp, M.; Vida, I.; Larkum, M.E. Dendritic action potentials and computation in human layer 2/3 cortical neurons. Science 2020, 367, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Ramon y Cajal, S. Histologie du Système Nerveux de l’Homme et des Vertébrés; Maloine: Paris, France, 1911; Volume 2, pp. 153–173. [Google Scholar]
- Poirazi, P.; Brannon, T.; Mel, B.W. Pyramidal neuron as two-layer neural network. Neuron 2003, 37, 989–999. [Google Scholar] [CrossRef] [Green Version]
- Hubel, D.H.; Wiesel, T.N. Receptive fields of optic nerve fibres in the spider monkey. J. Physiol. 1960, 154, 572–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, K.-L.; Gupta, H.; Sorooshian, S. Artificial neural network modeling of the rainfall-runoff process. Water Resour. Res. 1995, 31, 2517–2530. [Google Scholar] [CrossRef]
- Hassoun, M.H.; Intrator, N.; McKay, S.; Christian, W. Fundamentals of artificial neural networks. Comput. Phys. 1996, 10, 137. [Google Scholar] [CrossRef] [Green Version]
- Al-Shayea, Q.K. Artificial neural networks in medical diagnosis. Int. J. Comput. Sci. Issues 2011, 8, 150–154. [Google Scholar]
- Khashei, M.; Bijari, M. An artificial neural network (p,d,q) model for timeseries forecasting. Expert Syst. Appl. 2010, 37, 479–489. [Google Scholar] [CrossRef]
- Guresen, E.; Kayakutlu, G.; Daim, T.U. Using artificial neural network models in stock market index prediction. Expert Syst. Appl. 2011, 38, 10389–10397. [Google Scholar] [CrossRef]
- McCulloch, W.S.; Pitts, W. A logical calculus of the ideas immanent in nervous activity. Bull. Math. Biophys. 1943, 5, 115–133. [Google Scholar] [CrossRef]
- London, M.; Häusser, M. Dendritic computation. Annu. Rev. Neurosci. 2005, 28, 503–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agmon-Snir, H.; Carr, C.E.; Rinzel, J. The role of dendrites in auditory coincidence detection. Nat. Cell Biol. 1998, 393, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.C.; Binzegger, T.; Kahana, O.; Martin, K.A.C.; Segev, I. Dendritic asymmetry cannot account for directional responses of neurons in visual cortex. Nat. Neurosci. 1999, 2, 820–824. [Google Scholar] [CrossRef]
- Artola, A.; Brocher, S.; Singer, W. Different voltage-dependent thresholds for inducing long-term depression and long-term potentiation in slices of rat visual cortex. Nat. Cell Biol. 1990, 347, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Euler, T.; Detwiler, P.; Denk, W. Directionally selective calcium signals in dendrites of starburst amacrine cells. Nat. Cell Biol. 2002, 418, 845–852. [Google Scholar] [CrossRef]
- Magee, J.C. Dendritic integration of excitatory synaptic input. Nat. Rev. Neurosci. 2000, 1, 181–190. [Google Scholar] [CrossRef]
- Single, S.; Borst, A. Dendritic integration and its role in computing image velocity. Science 1998, 281, 1848–1850. [Google Scholar] [CrossRef] [Green Version]
- Spruston, N.; Stuart, G.; Hausser, M. Dendritic integration. In Dendrites; Stuart, G., Spruston, N., Hausser, M., Eds.; Oxford University Press: Oxford, UK, 2008. [Google Scholar]
- Dringenberg, H.C.; Hamze, B.; Wilson, A.; Speechley, W.; Kuo, M.-C. Heterosynaptic facilitation of in vivo thalamocortical long-term potentiation in the adult rat visual cortex by acetylcholine. Cereb. Cortex 2006, 17, 839–848. [Google Scholar] [CrossRef]
- Koch, C.; Poggio, T.; Torre, V. Nonlinear interactions in a dendritic tree: Localization, timing, and role in information processing. Proc. Natl. Acad. Sci. USA 1983, 80, 2799–2802. [Google Scholar] [CrossRef] [Green Version]
- Koch, C.; Poggio, T.; Torre, V. Retinal ganglion cells: A functional interpretation of dendritic morphology. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1982, 298, 227–263. [Google Scholar]
- Kepecs, A.; Wang, X.-J.; Lisman, J. Bursting neurons signal input slope. J. Neurosci. 2002, 22, 9053–9062. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Image Pixel | Scan Window | Extracted Orientation Information | Output Orientation Information | Scanning Step |
|---|---|---|---|---|
| 32 × 32 | 3 × 3 | 8 | 4 | 1 |
| Object Type | Orientation Angle | ||||
|---|---|---|---|---|---|
| 0° | 45° | 90° | 135° | ||
| 2 pixels | No. of samples | 928 | 841 | 928 | 841 |
| Correct numbers | 928 | 841 | 928 | 841 | |
| Accuracy | 100% | 100% | 100% | 100% | |
| 3 pixels | No. of samples | 960 | 900 | 960 | 900 |
| Correct numbers | 960 | 900 | 960 | 900 | |
| Accuracy | 100% | 100% | 100% | 100% | |
| 4 pixels | No. of samples | 992 | 961 | 992 | 961 |
| Correct numbers | 992 | 961 | 992 | 961 | |
| Accuracy | 100% | 100% | 100% | 100% | |
| 8 pixels | No. of samples | 1699 | 2249 | 1699 | 2249 |
| Correct numbers | 1699 | 2249 | 1699 | 2249 | |
| Accuracy | 100% | 100% | 100% | 100% | |
| 12 pixels | No. of samples | 2379 | 3411 | 2379 | 3411 |
| Correct numbers | 2379 | 3411 | 2379 | 3411 | |
| Accuracy | 100% | 100% | 100% | 100% | |
| 16 pixels | No. of samples | 1319 | 1489 | 1319 | 1489 |
| Correct numbers | 1319 | 1489 | 1319 | 1489 | |
| Accuracy | 100% | 100% | 100% | 100% | |
| 32 pixels | No. of samples | 1284 | 1645 | 1284 | 1645 |
| Correct numbers | 1284 | 1645 | 1284 | 1645 | |
| Accuracy | 100% | 100% | 100% | 100% | |
| ≥48 pixels | No. of samples | 2515 | 1275 | 2515 | 1275 |
| Correct numbers | 2515 | 1275 | 2515 | 1275 | |
| Accuracy | 100% | 100% | 100% | 100% | |
| Layers | Parameters | Learning Cost | Reasoning | Bio-Soundness | Noise Resistance | |
|---|---|---|---|---|---|---|
| CNN | >7 | 820,004 | High | Black Box | Low | Low |
| AVS | 2 | 12 | No | Reasonable | High | High |
| NOISES | 0% | 5% | 10% | 15% | 20% | 25% | 30% |
|---|---|---|---|---|---|---|---|
| CNN | 99.887% | 90.783% | 74.441% | 59.108% | 47.547% | 39.866% | 35.343% |
| AVS | 100% | 96.571% | 85.562% | 71.490% | 59.716% | 49.924% | 43.452% |
 | The output is 0 degree |
 | The output is 45 degree |
 | The output is 90 degree |
 | The output is 135 degree |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Zheng, T.; Todo, Y. The Mechanism of Orientation Detection Based on Artificial Visual System. Electronics 2022, 11, 54. https://doi.org/10.3390/electronics11010054
Zhang X, Zheng T, Todo Y. The Mechanism of Orientation Detection Based on Artificial Visual System. Electronics. 2022; 11(1):54. https://doi.org/10.3390/electronics11010054
Chicago/Turabian StyleZhang, Xiliang, Tang Zheng, and Yuki Todo. 2022. "The Mechanism of Orientation Detection Based on Artificial Visual System" Electronics 11, no. 1: 54. https://doi.org/10.3390/electronics11010054