1. Introduction
Skin is the largest organ that can protect the body from the outer environment, keep the body hydrated, and play an essential role as an immunologic barrier [
1,
2]. The epidermis is the outermost layer of the skin containing melanin which determines skin color. Melanin plays a crucial role in photoprotection through absorbing UV and scavenging free radicals. Melanogenesis, a process of melanin formation in melanocytes, is a multistage process of tyrosine oxidation and polymerization melanin [
3,
4]. Melanogenesis involves the regulation of various signal molecules and transcription factors that influence the microphthalmia transcription factor (MITF) and its downstream proteins, including tyrosinase-related protein-1 (TRP-1), tyrosinase-related protein-2 (TRP-2), and tyrosinase [
5].
Hyperpigmentation causes dark spots and freckles on some areas of the skin. On the contrary, a lack of melanin can cause skin diseases such as skin cancer and vitiligo [
6]. The difference in the extent of skin photoprotection in accordance with the content and composition of melanin contributes to the incidence of melanoma. It is well reported that the prevalence of skin cancer is higher in light-skinned people than dark skin [
7]. Hair greying is caused by a loss of melanin in hair follicles [
8]. Therefore, it is important not only to reduce melanin pigment, but also to induce melanogenesis. Hair greying is the most representative hair aging symptom caused by a decrease in the number of follicular melanocytes or a decrease in melanin production [
9]. In this study,
Sargassum fusiforme was noted as a candidate that could increase melanin related to the cAMP/PKA/CREB signaling pathway.
The cAMP/PKA/CREB signaling pathway plays a crucial role in regulating melanogenesis, starting with activation of the G protein-coupled receptor, melanocortin 1 receptor (MC1R), via the melanocyte-stimulating hormone (MSH) [
10]. This leads to activation of the G protein, which in turn activates adenylyl cyclase, an enzyme that converts ATP to cAMP. Increased cAMP levels activate protein kinase A (PKA), an enzyme that phosphorylates various target proteins, including the cAMP-response element binding protein (CREB), a transcription factor that regulates the expression of genes involved in melanogenesis [
11]. Phosphorylated CREB binds to cAMP response elements (CREs) on specific DNA sequences and upregulates the expression of enzymes, such as tyrosinase, TRP-1, and TRP-2 involved in melanogenesis, which lead to an increase in melanin pigmentation and a darker skin color. This mechanism is responsible for the tanning response to UV radiation [
12]. This has also been applied to hypopigmentation disorders such as vitiligo and grey hair.
Sargassum fusiforme is a species of brown seaweed that belongs to the genus
Sargassum. It grows abundantly along the rocky coastline of Asian countries such as Korea, Japan, and China. Along with being used as a food source, seaweed is also a source of structurally diverse active compounds that have biomedical potential to develop functional substances for foods and cosmetics [
13]. Among compounds isolated from
S. fusiforme, polysaccharides have received considerable attention in the scientific community due to their potential health promotion effects [
14]. Polysaccharides isolated from
S. fusiforme, which include lamiginine, alginate, fucoidan, and dietary fiber, have been of particular interest due to their potential health promotion effects [
15]. These polysaccharides have various biological activities and are the subject of much scientific research.
Although
Sargassum fusiforme extract (SFE) is commonly used in cosmetic products due to its potential skin benefits such as moisturizing, anti-inflammatory, and antioxidant effects, its hyper-pigmenting effect has not been reported yet [
16,
17].
Thus, this study aimed to investigate the effect of Sargassum fusiforme extract (SFE) on melanin synthesis in human epidermal melanocytes. It was found that SFE could induce melanogenesis by increasing the expression of MITF gene via activation of the cAMP/PKA/CREB signaling pathway.
2. Materials and Methods
2.1. Sargassum Fusiforme Extract
The aerial part of
Sargassum fusiforme was obtained from the Bomyeongsanghoe Co. (Jeju, Republic of Korea). They were thoroughly washed to completely remove impurities, dried at 35 °C to 40 °C, and extracted with water at 100 °C for 2 h and filtered. After cooling the filtrate, 2% of calcium chloride compared to the raw material was added, and a precipitate was formed via the reaction between calcium chloride and alginate, and the precipitate was removed via filtering. The extract obtained after removing the alginate was precipitated by adding 3 folds of ethanol. After precipitation, only the supernatant was collected by passing through a 5 µm filter and concentrated in a vacuum concentrator (Rotavapor
® R-215, Bu-chi, Flawil, Switzerland). Then, it was freeze-dried (TEFIC TF-10D, Tefic Biotech CO, Xi’an, China) to prepare
Sargassum fusiforme extract (SFE) [
18].
2.2. Cell Culture
Normal Human Epidermal Melanocytes, neonatal, and moderately pigmented donors (HeMn-MP) were purchased from Thermo Fisher Scientific (Waltham, MA, USA). These cells were cultured in a M254 medium supplemented with a human melanocyte growth solution (HMGS) at 37 °C in 5% CO2. All experiments were performed using the cells in passages 4–9.
2.3. Chemical Reagents and Antibodies
Tyrosine, 3,4-dihydroxyphenilalanine (L-DOPA), and dimethyl sulfoxide (DMSO) were purchased from Sigma-Aldrich (St Louis, MO, USA). Antibodies against MITF, TRP-1, tyrosinase, and β-actin were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Antibodies against CREB, p-CREB, H89 dihydrochloride, and the protein kinase A (PKA) inhibitor were obtained from Cell Signaling Technology (Beverly, MA, USA). Culture medium M254, the human melanocyte growth solution (HMGS), and the hydrogen peroxide solution were purchased from Thermo Fisher Scientific (Waltham, MA, USA).
2.4. Cell Viability Measurement and Melanin Contents Assay
HeMn-MP were initially seeded into 6-well plates at a density of 1.5 × 105 cells/well and incubated at 37 °C with 5% CO2 overnight. The next day, the cells were treated with a medium containing various concentrations of SFE for 5 days. Prior to cell collection, cell viability was measured using a WST-1 solution (DoGenBio, Seoul, Republic of Korea) after 4 h of incubation. Afterward, the supernatant was transferred to a new 96-well plate for absorbance measurement at 450 nm using a spectrophotometer (BioTek, Winooski, VT, USA).
The remaining cells were used for determining melanin content. The cells were washed with PBS and lysed with 1 N of NaOH for 1 h at 60 °C. Following this, the lysate was centrifuged, and the supernatant was collected to determine the amount of melanin. The absorbance of supernatant was measured at 450 nm.
2.5. Oxidative Stress Induction with Hydrogen Peroxide
HeMn-MP were seeded in the same way as described in the melanin contents assay section. After seeding, they were exposed to 10 to 150 µg/mL of SFE for 6 h before H2O2 treatment. After 200 µM of H2O2 was added to each well, the cells were incubated with H2O2 for 5 days. Cell viability and melanin contents were measured using the same method as described in the previous section.
2.6. Western Blot Analysis
The expression levels of melanogenesis-related proteins in HeMn-MP were detected using Western blot analysis. HeMn-MP cells at a density of 2 × 105 cells per dish were cultured in 6 cm dishes. After adding various concentrations of SFE to the culture medium, the cells were incubated for 5 days. The cells were lysed with a cell lysis buffer (CytoBuster™; Merck, Darmstadt, Germany) and the protein concentration was determined using the BCA protein assay (Thermo Fisher Scientific, Waltham, MA, USA). Equal amounts of proteins (30 µg/lane) were loaded and separated via sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), followed by an electrotransfer to polyvinylidene difluoride (PVDF) membranes. Blots were blocked with 5% skim milk for 30 min at room temperature and probed with primary antibodies (MITF, tyrosinase, CREB, p-CREB, TRP-1, or β-actin) overnight on ice. These membranes were then incubated with a horseradish peroxidase (HRP)-conjugated anti-mouse or anti-rabbit secondary antibody at room temperature for one hour. Subsequently, these membranes were washed with Tris-buffered saline with 0.1% Tween 20 Detergent (TBS-T) for 10 min in three times, and the bands were visualized using an enhanced chemiluminescence system (PicoEPD Western reagent; LPIS-Biotech, Daejeon, Republic of Korea) with an LAS 500 image analyzer (GE Healthcare Life Sciences, Seongnam-Si, Republic of Korea). The expression of each protein was analyzed using the ImageJ program (National Institutes of Health, Bethesda, MD, USA) and visualized as a graph. Results were confirmed by three independent experiments.
2.7. Tyrosinase Zymography
HeMn-MP cells were incubated with SFE for 3 days and washed three times with Dulbecco’s phosphate-buffered saline (DPBS). The cells were subsequently lysed with a cell lysis buffer (CytoBuster™; Merck, Darmstadt, Germany), and the protein content was measured using a BCA protein assay (Thermo Fisher Scientific, Waltham, MA, USA). Equal amounts of proteins (20 µg) were mixed with a 4× sample buffer and resolved via 4–12% SDS-PAGE. After electrophoresis, the tyrosinase activity of lysates was determined by an additional process. SDS gel was incubated with a rinse buffer (0.1 M of sodium phosphate buffer, pH 6.8) for 20 min at room temperature. Then, the gel was rinsed and incubated with a rinse buffer containing 5 mM of L-DOPA, a substrate in melanin synthesis, at 37 °C in the dark for 15 min.
2.8. Measurement of Cellular Tyrosinase Activity
For cellular tyrosinase activity, HeMn-MP cells were seeded into 24-well plates at a density of 5 × 104 cells/well and stabilized in a culture medium overnight. The next day, the cells were treated with a medium containing various concentrations of SFE for 72 h. The medium was removed and replaced with 1% Triton X-100 containing 0.1 M of a phosphate buffer (pH 6.5) and 0.1 mM of phenyl methane sulfonyl fluoride (PMSF). The mixture was frozen at −80 °C, thawed at room temperature, and then centrifuged at 13,000 rpm for 30 min at 4 °C. Reaction mixtures consisting of 50 mM of the phosphate buffer (pH 6.5), 100 µg of the supernatant protein and 2 mg/mL of L-DOPA were assayed on a 96-well plate at 37 °C for 30 min and the absorbance was measured at 490 nm using a spectrophotometer (BioTek, Winooski, VT, USA).
2.9. Reconstituted Three-Dimensional Human Skin Model
To investigate the effect of SFE on melanogenesis in a 3D skin model, a commercially available reconstituted human skin model (KeraSkin-M, BioSolution, Seoul, Republic of Korea) was used. The skin model contained both human epidermal keratinocytes and melanocytes. The skin model was cultured with SFE for 13 days in a maintenance medium (BioSolution Science, Seoul, Republic of Korea). To measure the degree of melanin synthesis, L* values were measured with a CR-300 chromameter (Minolta, Tokyo, Japan). Following this, the skin model was treated with 1 N of NaOH and incubated at 70 °C for 4 h to dissolve melanin. The amount of dissolved melanin was measured using a spectrophotometer (BioTek, Winooski, VT, USA) at 450 nm. All image data were analyzed with Image-Pro Plus (Media Cybernetics, Rockville, MD, USA) and expressed with statistically significant L* values.
2.10. Ex Vivo Human Hair Follicles
Pigmented hair follicles were obtained from a female donor. On a scalp plasty for a 67-year-old woman, 56 entire hair follicles (from the bulb to the infundibulum) were isolated via microdissection. Hairs exhibiting a low pigmentation in the hair shaft and medium pigmentation in the bulb were selected for this study. Hair follicles were placed in 48-well plates and incubated at 37 °C in a 5% CO2 incubator. They were then treated with 50 or 100 µg/mL of SFE for 9 days in William’s medium (Sigma-Aldrich, St Louis, MO, USA) supplemented with L-glutamine, insulin–transferrin–selenium G, hydrocortisone, penicillin and streptomycin antibiotics, and antifungal amphotericin B (300 µL per well).
After incubation, hair follicles were fixed in a buffered formalin solution for 30 min. After that, the samples were dehydrated, embedded in paraffin, sectioned (5 μm thick serial sections) using a microtome, and mounted on histological glass slides. Microscopical observations were performed using a Leica DMLB, an Olympus BX43, or a BX63 microscope. Images were captured with a DP72 or DP74 Olympus camera with Cell^D or CellSens Dimension (version 1.6) software (Olympus Life Science, Tokyo, Japan).
To assess the cell viability of the hair bulb, Masson’s trichrome staining was performed on formalin-fixed paraffin-embedded (FFPE) skin sections and observed microscopically. Melanin was visualized after silver impregnation according to the Fontana–Masson staining method using FFPE skin sections. Staining was assessed via microscopical observation and image analysis. Melanin contents in hair follicles were quantitatively measured as the melanin surface normalized to the hair follicle area.
2.11. Statistical Analysis
All experiments were conducted independently three times, and the data are presented as mean ± standard deviations. To assess the statistical significance of the results, unpaired two-tailed Student’s t-test was used to compare the treated cells with the control (untreated) cells. The significance level was set at * p < 0.05, ** p < 0.01, and *** p < 0.001, indicating a significant difference.
4. Discussion
Sargassum is a genus of brown seaweeds sourced from a wide and diverse range, holding promise in various domains, including both food and alternative medical applications [
19]. It is renowned not only for its exceptional nutritional value but also for being a rich repository of various natural antioxidant compounds such as polyphenols, carotenoids, meroterpenoids, and plant sterols.
Sargassum possesses qualities that position it as a valuable resource with significant potential for the development of commercially viable products across a range of fields, including food, biofuel, agriculture, therapeutic, and cosmetic products [
20].
Brown algae within the
Sargassum genus have been shown to exhibit diverse physiological benefits, including antioxidative effects [
21]. However, there has been no evidence of their ability to enhance melanogenesis. Additionally, these large
Sargassum seaweeds can be cultivated through aquaculture and can also be produced in various forms, including powders, tablets, or capsules, all of which are now widely involved in various biological applications, which may be an advantage of
Sargassum over other natural materials [
22].
Previous studies have demonstrated that cAMP is involved in the regulation of melanogenesis, the process by which skin cells produce the pigment melanin [
12]. The elevated levels of cAMP can activate protein kinase A (PKA), leading to the phosphorylation and activation of enzymes involved in melanin synthesis and ultimately resulting in increased melanogenesis. This process helps darken the skin in response to sun exposure, providing protection against ultraviolet radiation.
A recent study about the regulation of melanin synthesis has determined the melanogenic and antioxidant activities of Argan leaves extract. Phenol profiling was also performed via UPC-ESI-HRMS analysis to identify phenol compounds present in the extract [
23]. Similarly, a study on
Vernonia anthelmintica (L.) Willd, a traditional Chinese herbal medicine, has revealed its potential in promoting pigmentation for the treatment of skin depigmentation disorders by reducing the accumulation of ROS and inflammatory response [
24,
25].
Regarding oxidative stress, previous research conducted by Wood (2009) examined the effects of H
2O
2-induced oxidative damage on the human hair follicle, including the hair shaft, which is a critical factor in age-related hair greying [
26]. Numerous studies have investigated the protective mechanisms of melanocytes against oxidative stress via the Nrf2 and MITF signaling pathways [
27,
28].
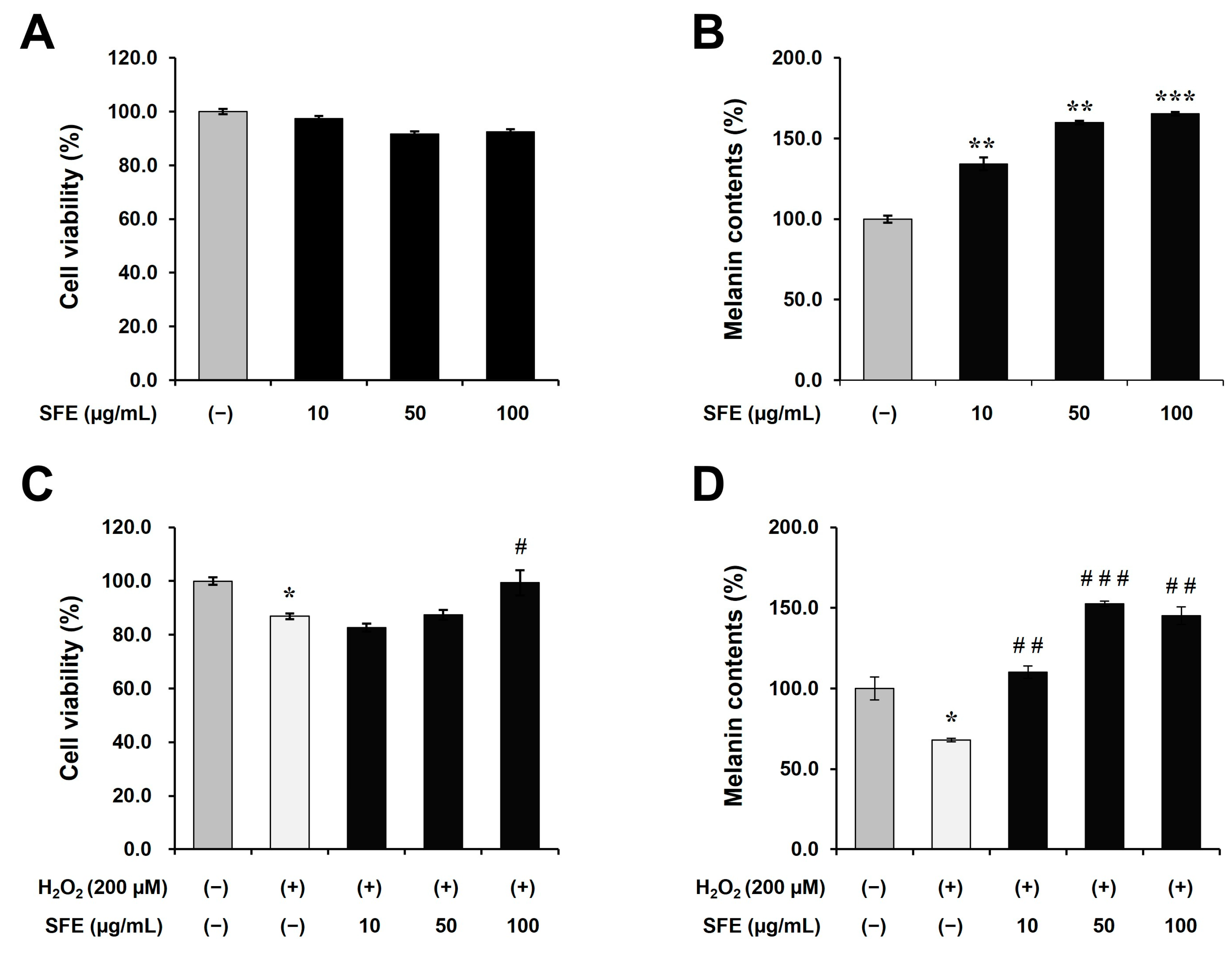
The present study aimed to investigate the effects of
Sargassum fusiforme extract (SFE) on melanin content and tyrosinase activity without causing cytotoxicity in moderately pigmented human epidermal melanocytes. (
Figure 1). This finding was also confirmed under oxidative stress conditions. Oxidative stress and melanogenesis have a complex relationship. Oxidative stress refers to an imbalance between production of reactive oxygen species and the cell’s ability to detoxify them, leading to cellular damage. Many studies have investigated inhibitory effects of reactive oxygen species (ROS) on melanin production [
29]. Reactive oxygen species (ROS) have been shown to inhibit melanin production by damaging melanocytes, which is a proposed mechanism for vitiligo and hair greying. Therefore, we investigated the potential of SFE to counteract this effect. As shown in
Figure 1, the reduction in melanin content and cell viability were restored via SFE treatment. Additionally, we found that SFE increased melanogenesis by promoting the phosphorylation of CREB via the cAMP/PKA/CREB signaling pathway and upregulating MITF in human epidermal melanocytes (
Figure 2). These results suggest that
Sargassum fusiforme extract may have a promoting effect on pigmentation.
Additionally, we conducted HPLC profiling for several compounds by performing LC-MS/MS to find out which components of
Sargassum fusiforme were responsible for this effect. However, we could not identify the candidates that caused pigmentation. In the results of the total equivalent analysis and total polyphenol analysis, the total sugar content was 58.8%. Contents of total polyphenols were analyzed to be 5.04 mg of GAE/g. However, based on a previous study, sugar is not expected to cause pigmentation. We also found that SFE had a high iron content. Iron is known to catalyze decomposition of hydrogen peroxide to hydroxyl radical and hydroxyl ions with melanin through the Fenton reaction [
30]. When melanin is exposed to the ferrous iron in the Fenton system, the rapid formation of melanin is observed [
31]. Those results indicate that iron can contribute to enhancing melanogenesis via the Fenton reaction. Moreover, previous studies have already reported that iron components can improve hair greying [
32]. Thus, it can be expected that melanin production is increased in iron-rich extracts. Additional research is needed on whether the ingredient that induces the pigmentation efficacy of SFE is iron.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




