
Carbon Sequestration Potential of Forest Invasive Species: A Case Study with Acacia dealbata Link

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
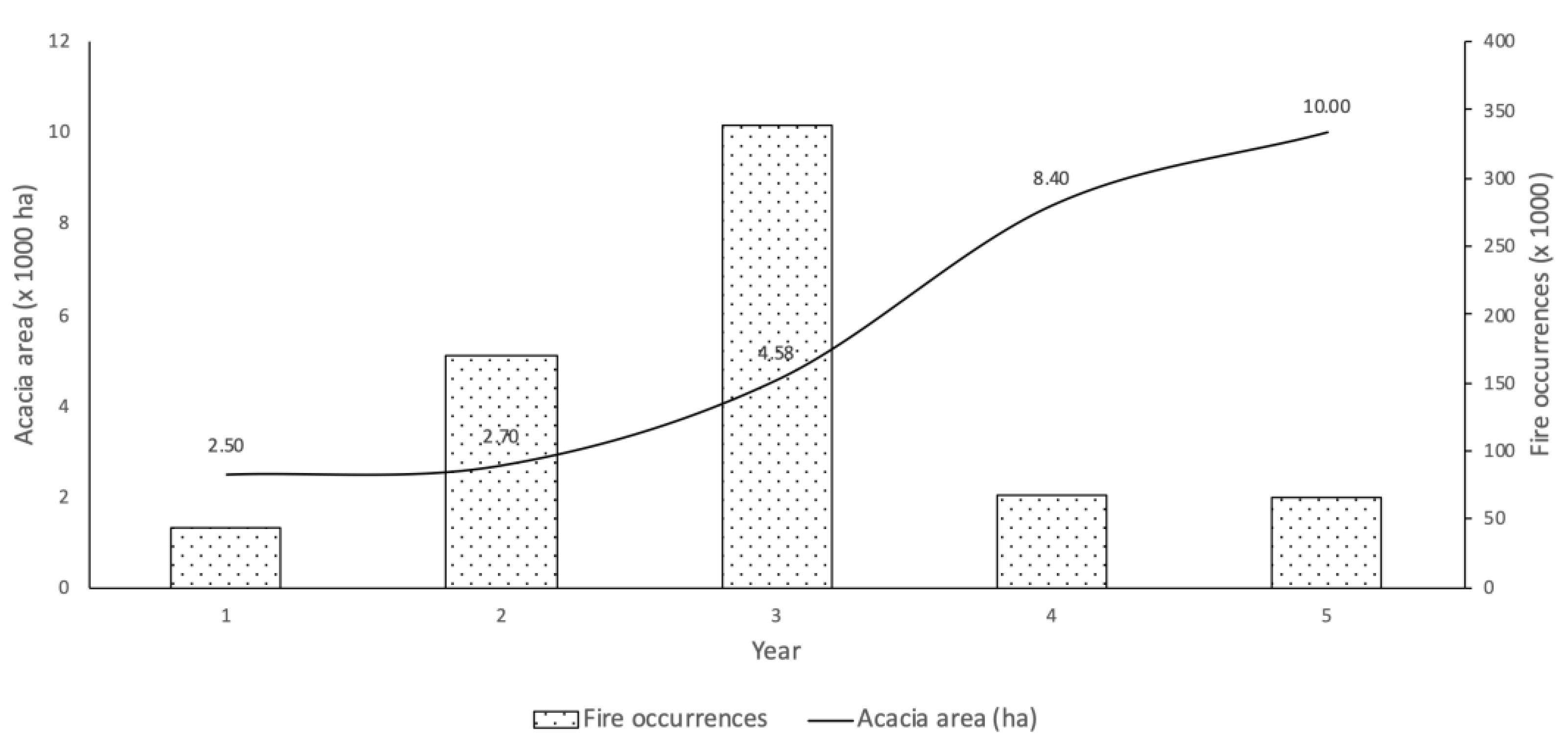
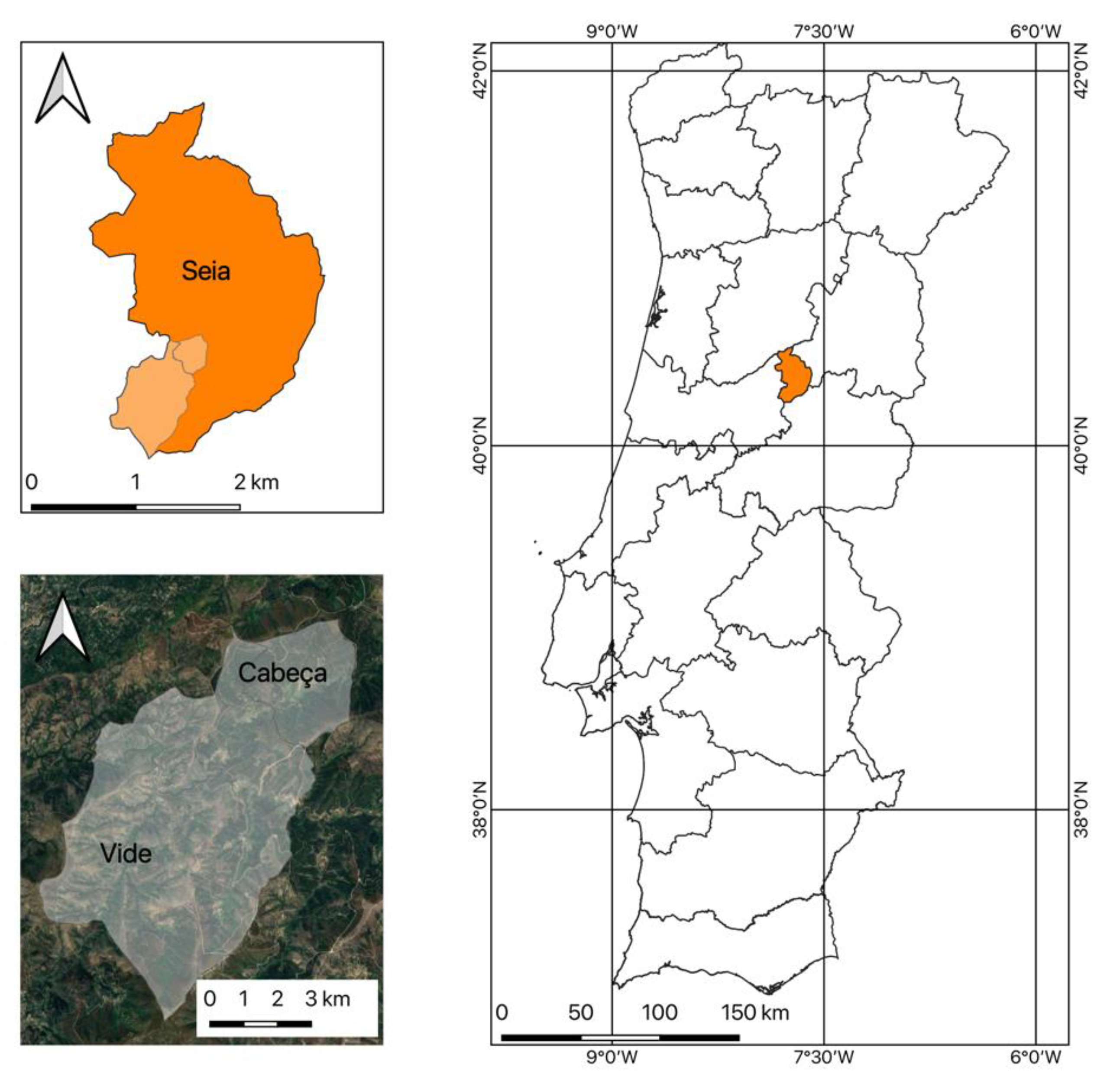
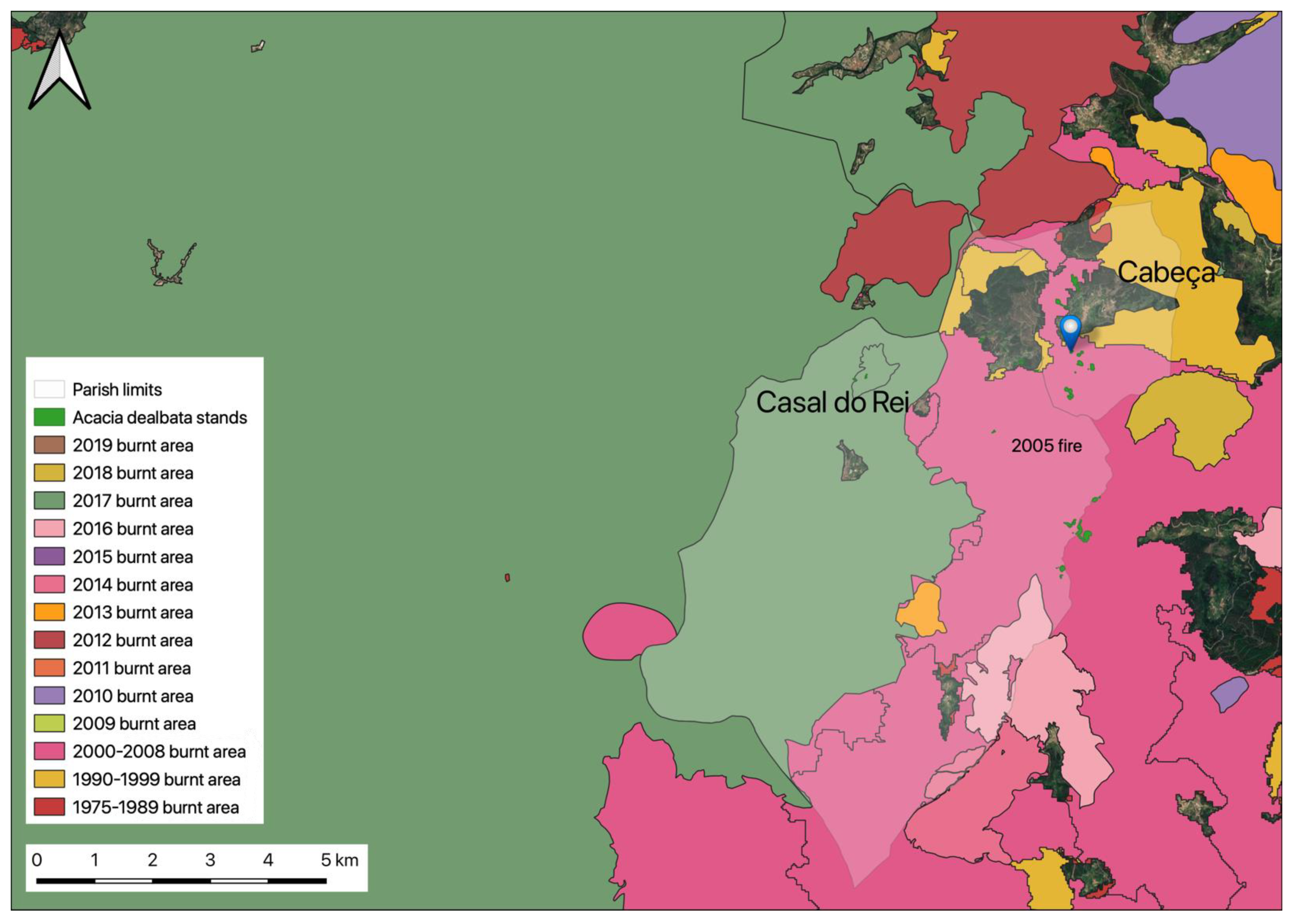
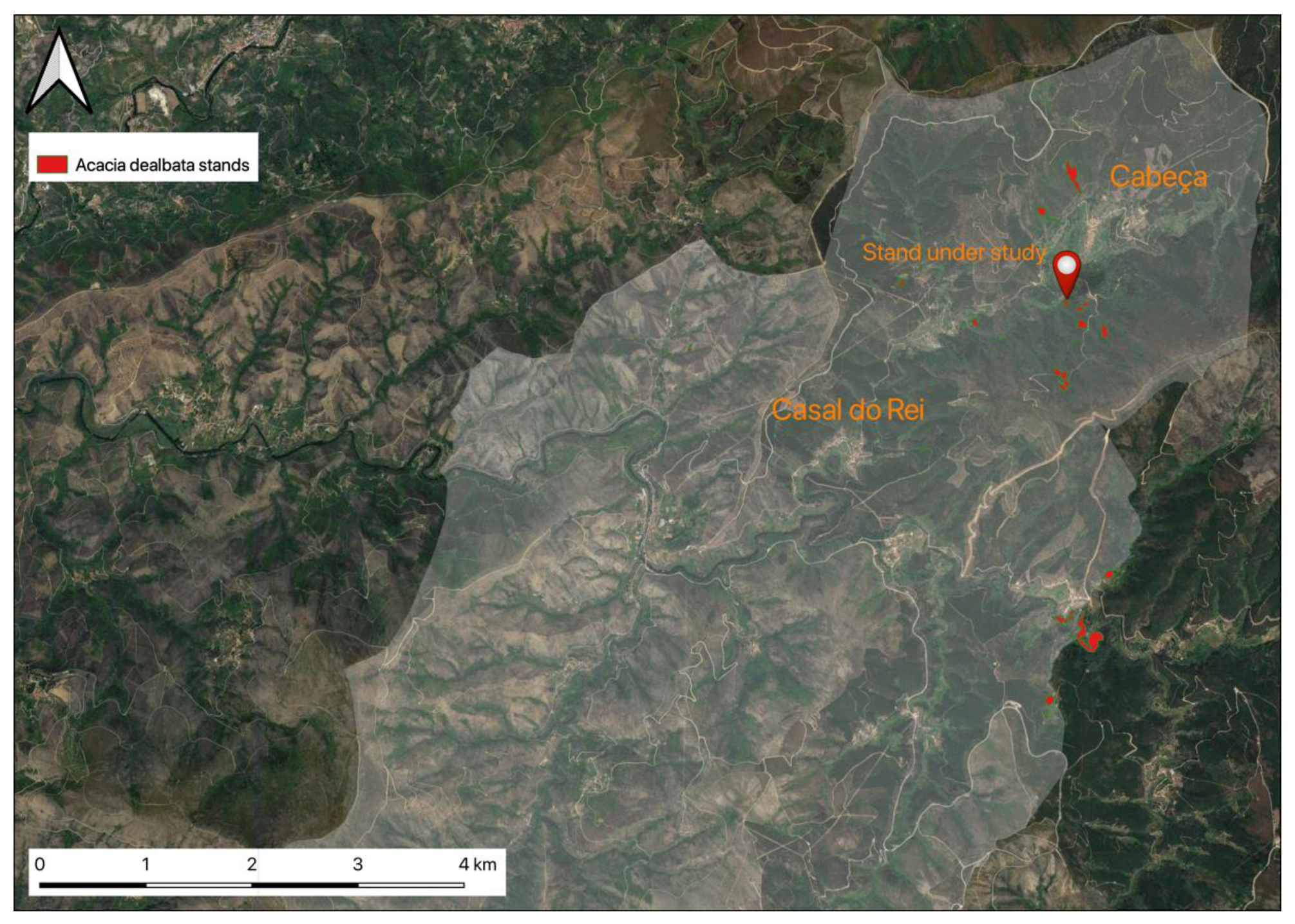
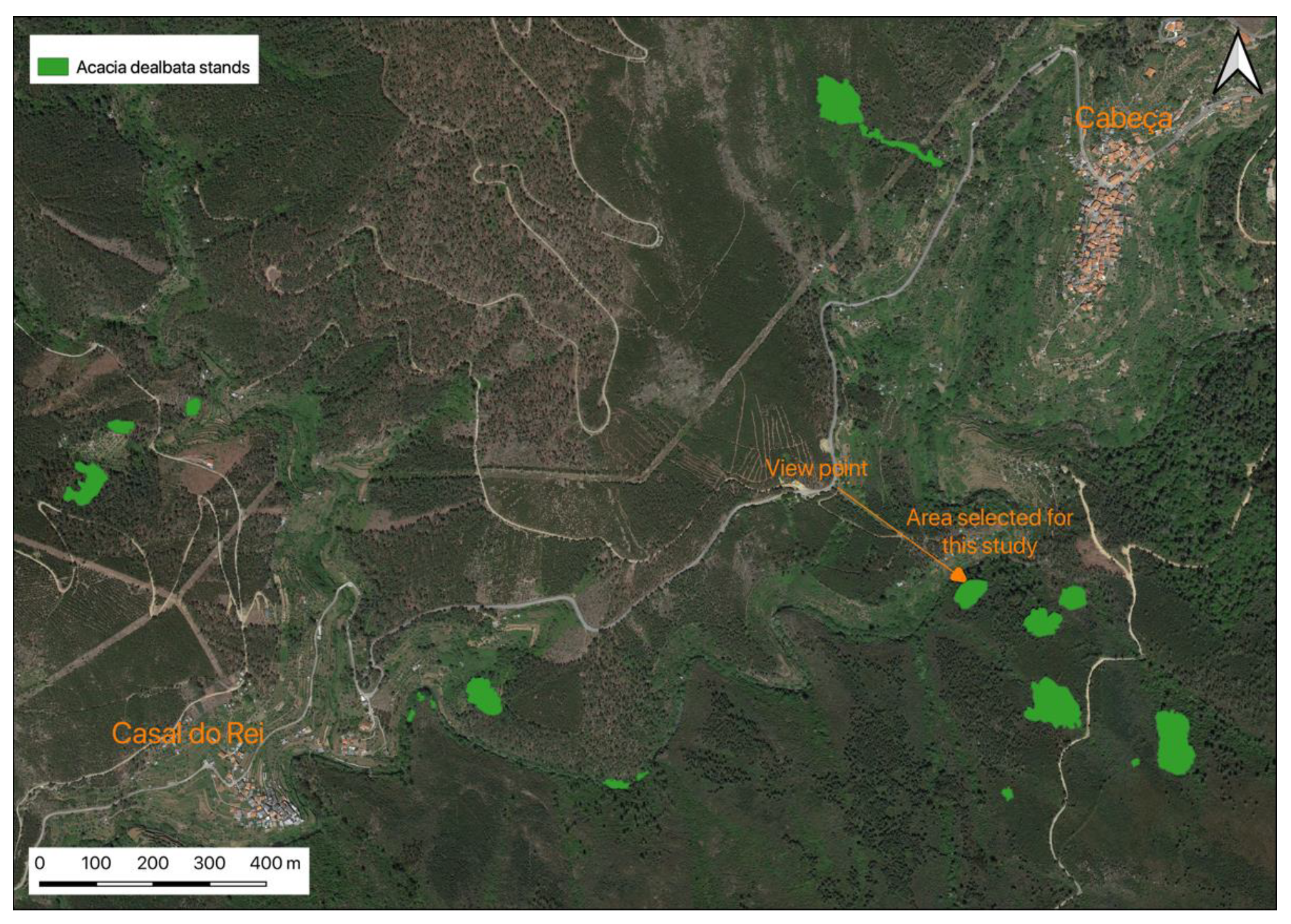
2.1. Location and A. dealbata Sampling
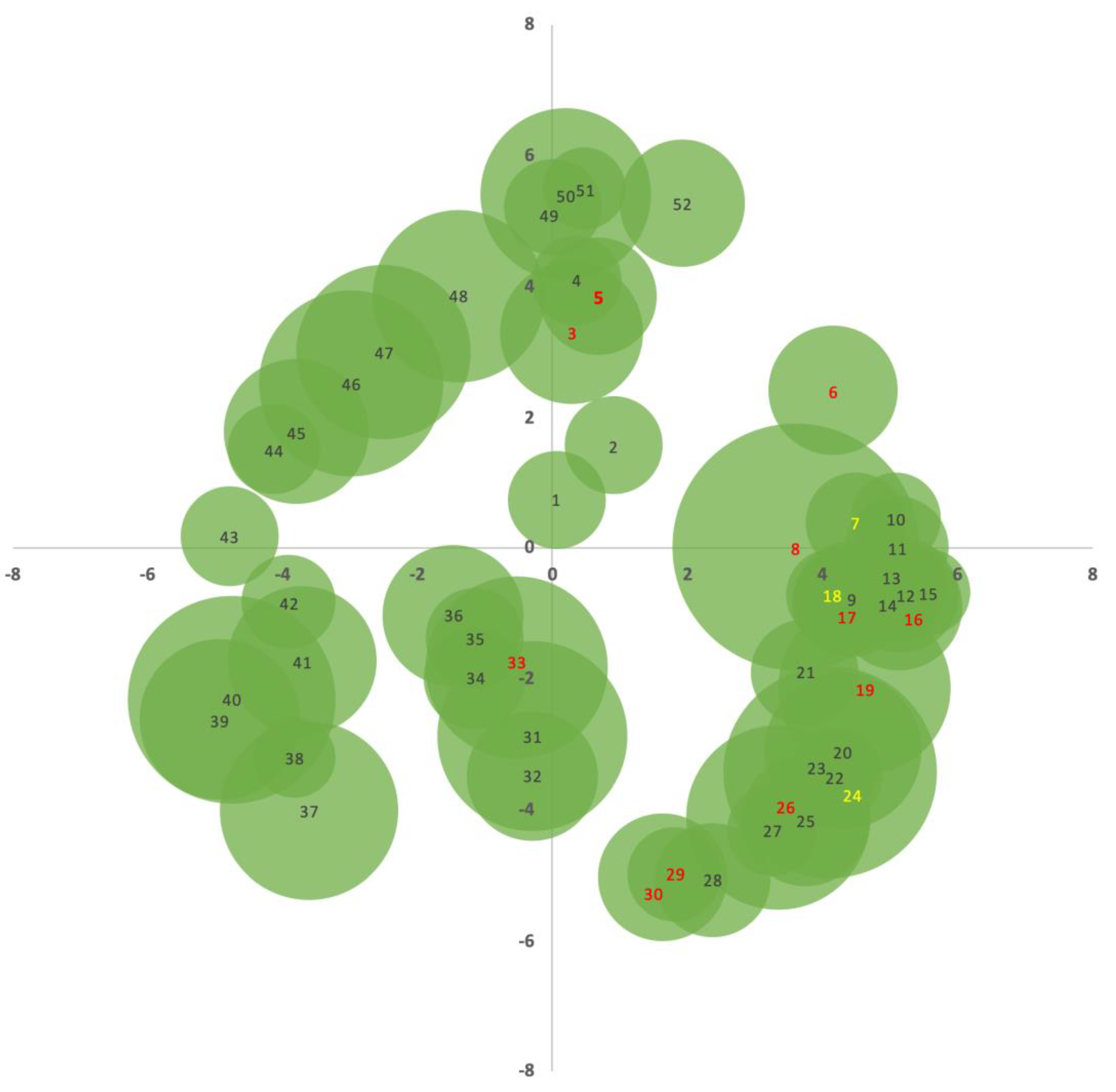
2.2. Dendrometric Analysis
2.3. Elemental Analysis
2.4. Determination of the Total Amount of Carbon Weight and Annual Rate of Carbon Storage
- Step 1: Determination of the wet weight of aerial biomass
- Step 2: Determination of the dry weight of the tree
- Step 3: Determination of the carbon weight of the tree
- Step 4: Determination of the amount of CO2 sequestered in the tree
- Step 5: Determination of the average amount of CO2 sequestration
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sanderson, L.A.; McLaughlin, J.A.; Antunes, P.M. The last great forest: A review of the status of invasive species in the North American boreal forest. Forestry 2012, 85, 329–340. [Google Scholar] [CrossRef]
- Allen, E.B.; Steers, R.J.; Dickens, S.J. Impacts of Fire and Invasive Species on Desert Soil Ecology. Rangel. Ecol. Manag. 2011, 64, 450–462. [Google Scholar] [CrossRef]
- Courchamp, F.; Caut, S.; Bonnaud, E.; Bourgeois, K.; Angulo, E.; Watari, Y. Eradication of alien invasive species: Surprise effects and conservation successes. Island Invasives 2011, 1, 285–289. [Google Scholar]
- Midgley, G.F.; Rutherford, M.C.; Davis, G.W.; Bosenberg, J.D.W. Photosynthetic Responses of Heliophilous Rhus Species to Environmental Modification by Invasive Shrubs. Funct. Ecol. 1992, 6, 334. [Google Scholar] [CrossRef]
- González-Muñoz, N.; Costa-Tenorio, M.; Espigares, T. Invasion of alien Acacia dealbata on Spanish Quercus robur forests: Impact on soils and vegetation. For. Ecol. Manag. 2012, 269, 214–221. [Google Scholar] [CrossRef]
- Barbero, M.; Bonin, G.; Loisel, R. Changes and disturbances of forest ecosystems caused by human activities in the western part of the mediterranean basin. Vegetatio 1990, 87, 151–173. [Google Scholar] [CrossRef]
- Brunel, S.; Schrader, G.; Brundu, G.; Fried, G. Emerging invasive alien plants for the Mediterranean Basin. EPPO Bull. 2010, 40, 219–238. [Google Scholar] [CrossRef]
- Hunt, M.A.; Battaglia, M.; Davidson, N.J.; Unwin, G.L. Competition between plantation Eucalyptus nitens and Acacia dealbata weeds in northeastern Tasmania. For. Ecol. Manag. 2006, 233, 260–274. [Google Scholar] [CrossRef]
- Bi, H.; Turvey, N. Inter-Specific Competition between Seedlings of Pinus radiata, Eucalyptus regnans and Acacia melanoxylon. Aust. J. Bot. 1994, 42, 61–70. [Google Scholar] [CrossRef]
- Forrester, D.I.; Bauhus, J.; Cowie, A.L. On the success and failure of mixed-species tree plantations: Lessons learned from a model system of Eucalyptus globulus and Acacia mearnsii. For. Ecol. Manag. 2005, 209, 147–155. [Google Scholar] [CrossRef]
- Bouillet, J.-P.; Laclau, J.-P.; Gonçalves, J.L.D.M.; Voigtlaender, M.; Gava, J.L.; Leite, F.P.; Hakamada, R.; Mareschal, L.; Mabiala, A.; Tardy, F.; et al. Eucalyptus and Acacia tree growth over entire rotation in single- and mixed-species plantations across five sites in Brazil and Congo. For. Ecol. Manag. 2013, 301, 89–101. [Google Scholar] [CrossRef]
- Correia, M.; Castro, S.; Ferrero, V.; Crisóstomo, J.A.; Rodríguez-Echeverría, S. Reproductive biology and success of invasive Australian acacias in Portugal. Bot. J. Linn. Soc. 2014, 174, 574–588. [Google Scholar] [CrossRef] [Green Version]
- Duarte, L.N.; Gomes, C.P.; Marchante, H.; Marchante, E. Integrating knowledge of ecological succession into invasive alien plant management: A case study from Portugal. Appl. Veg. Sci. 2020, 23, 328–339. [Google Scholar] [CrossRef]
- Le Maitre, D.C.; Kotzee, I.M.; O’Farrell, P.J. Impacts of land-cover change on the water flow regulation ecosystem service: Invasive alien plants, fire and their policy implications. Land Use Policy 2014, 36, 171–181. [Google Scholar] [CrossRef]
- Moreira, F.; Ferreira, A.; Abrantes, N.; Catry, F.; Fernandes, P.; Roxo, L.; Keizer, J.; Silva, J. Occurrence of native and exotic invasive trees in burned pine and eucalypt plantations: Implications for post-fire forest conversion. Ecol. Eng. 2013, 58, 296–302. [Google Scholar] [CrossRef]
- Aran, D.; García-Duro, J.; Reyes, O.; Casal, M. Fire and invasive species: Modifications in the germination potential of Acacia melanoxylon, Conyza canadensis and Eucalyptus globulus. For. Ecol. Manag. 2013, 302, 7–13. [Google Scholar] [CrossRef]
- Fernandes, M.M.; Devy-Vareta, N.; Rangan, H. Plantas exóticas invasoras e instrumentos de gestão territorial. O caso paradigmático do género Acacia em Portugal. GOT J. Geogr. Spat. Plan. 2013, 4, 83–107. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, S.; Monteiro, E.; Brito, P.; Vilarinho, C. Biomass resources in Portugal: Current status and prospects. Renew. Sustain. Energy Rev. 2017, 78, 1221–1235. [Google Scholar] [CrossRef]
- Briones-Hidrovo, A.; Copa, J.; Tarelho, L.A.; Gonçalves, C.; da Costa, T.P.; Dias, A.C. Environmental and energy performance of residual forest biomass for electricity generation: Gasification vs. combustion. J. Clean. Prod. 2021, 289, 125680. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pysek, P.; Rejmanek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and invasion of alien plants: Concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Pyšek, P.; Richardson, D.M. The biogeography of naturalization in alien plants. J. Biogeogr. 2006, 33, 2040–2050. [Google Scholar] [CrossRef]
- Águas, A.; Larcombe, M.J.; Matias, H.; Deus, E.; Potts, B.M.; Rego, F.C.; Silva, J.S. Understanding the naturalization of Eucalyptus globulus in Portugal: A comparison with Australian plantations. Eur. J. For. Res. 2017, 136, 433–446. [Google Scholar] [CrossRef]
- Lorenzo, P.; Gonzalez, L.O.; Reigosa, M.J. The genus Acacia as invader: The characteristic case of Acacia dealbata Link in Europe. Ann. For. Sci. 2010, 67, 101. [Google Scholar] [CrossRef] [Green Version]
- La Mantia, T.; Pasta, S.; Badalamenti, E. Acacia cyclops A. Cunn. ex G. Don (Leguminosae) in Italy: First cases of naturalization. An. Jardín Botánico Madr. 2012, 69, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Rouget, M.; Richardson, D.M. Inferring Process from Pattern in Plant Invasions: A Semimechanistic Model Incorporating Propagule Pressure and Environmental Factors. Am. Nat. 2003, 162, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Richardson, D.M.; Hui, C.; Nuñez, M.A.; Pauchard, A. Tree invasions: Patterns, processes, challenges and opportunities. Biol. Invasions 2014, 16, 473–481. [Google Scholar] [CrossRef]
- Peltzer, D.A.; Allen, R.B.; Lovett, G.; Whitehead, D.; Wardle, D.A. Effects of biological invasions on forest carbon sequestration. Glob. Chang. Biol. 2010, 16, 732–746. [Google Scholar] [CrossRef]
- Feng, Q.; Wang, B.; Chen, M.; Wu, P.; Lee, X.; Xing, Y. Invasive plants as potential sustainable feedstocks for biochar production and multiple applications: A review. Resour. Conserv. Recycl. 2021, 164, 105204. [Google Scholar] [CrossRef]
- Rogelj, J.; Shindell, D.; Jiang, K.; Fifita, S.; Forster, P.; Ginzburg, V.; Handa, C.; Kheshgi, H.; Kobayashi, S.; Kriegler, E. Mitigation pathways compatible with 1.5 °C in the context of sustainable development. In Global Warming of 1.5 °C; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2018; pp. 93–174. [Google Scholar]
- Toochi, E.C. Carbon sequestration: How much can forestry sequester CO2? For. Res. Eng. Int. J. 2018, 2, 148–150. [Google Scholar] [CrossRef] [Green Version]
- Clark, A.; Saucier, J.R.; McNab, W.H. Total-Tree Weight, Stem Weight and Volume Tables for Hardwood Species in the Southeast; Research Division; Georgia Forestry Commission: Milledgeville, GA, USA, 1986. [Google Scholar]
- Sá, L.C.R.; Loureiro, L.M.E.F.; Nunes, L.J.R.; Mendes, A.M. Torrefaction as a Pretreatment Technology for Chlorine Elimination from Biomass: A Case Study Using Eucalyptus globulus Labill. Resources 2020, 9, 54. [Google Scholar] [CrossRef]
- Nunes, L.J.; Raposo, M.A.; Meireles, C.I.; Gomes, C.J.P.; Ribeiro, N.M.A. Control of Invasive Forest Species through the Creation of a Value Chain: Acacia dealbata Biomass Recovery. Environments 2020, 7, 39. [Google Scholar] [CrossRef]
- Álvarez-Álvarez, P.; Pizarro, C.; Barrio-Anta, M.; Cámara-Obregón, A.; Bueno, J.L.M.; Álvarez, A.; Gutiérrez, I.; Burslem, D.F. Evaluation of Tree Species for Biomass Energy Production in Northwest Spain. Forests 2018, 9, 160. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, K.; Lal, R. Biochar application to soil for climate change mitigation by soil organic carbon sequestration. J. Plant Nutr. Soil Sci. 2014, 177, 651–670. [Google Scholar] [CrossRef] [Green Version]
- Stewart, C.E.; Zheng, J.; Botte, J.; Cotrufo, M.F. Co-generated fast pyrolysis biochar mitigates green-house gas emissions and increases carbon sequestration in temperate soils. GCB Bioenergy 2012, 5, 153–164. [Google Scholar] [CrossRef]
- Matovic, D. Biochar as a viable carbon sequestration option: Global and Canadian perspective. Energy 2011, 36, 2011–2016. [Google Scholar] [CrossRef]
- Gasol, C.M.; Brun, F.; Mosso, A.; Rieradevall, J.; Gabarrell, X.; Durany, X.G. Economic assessment and comparison of acacia energy crop with annual traditional crops in Southern Europe. Energy Policy 2010, 38, 592–597. [Google Scholar] [CrossRef]
- Lenis, Y.A.; Osorio, L.F.; Pérez, J.F. Fixed Bed Gasification of Wood Species with Potential as Energy Crops in Colombia: The Effect of the Physicochemical Properties. Energy Sources Part A Recover. Util. Environ. Eff. 2013, 35, 1608–1617. [Google Scholar] [CrossRef]
- Caldeira, M.; Saidelles, F.; Schumacher, M.; Godinho, T.D.O. Biomass in Acacia mearnsii De Wild stand, Rio Grande do Sul, Brasil. Sci. For. 2011, 39, 133–141. [Google Scholar]
- Acuña, E.; Cancino, J.; Rubilar, R.; Sandoval, S. Aboveground Biomass Growth and Yield of First Rotation Cutting Cycle of Acacia and Eucalyptus Short Rotation Dendroenergy Crops. Rev. Árvore 2017, 41, 41. [Google Scholar] [CrossRef]
- de Souza, J.B.; de Azevêdo, T.K.B.; Sousa, T.B.; da Silva, G.G.C.; Guimarães, J.B., Jr.; Pimenta, A.S. Colagem de compensados com adesivo à base de taninos de Acacia mangium Wild. cultivada no Nordeste do Brasil. Rev. Bras. Ciências Agrárias (Agrária) 2020, 15, 8659. [Google Scholar]
- Jing, Q.; Conijn, S.J.; Jongschaap, R.E.; Bindraban, P.S. Modeling the productivity of energy crops in different agro-ecological environments. Biomass Bioenergy 2012, 46, 618–633. [Google Scholar] [CrossRef]
- Richardson, D.M.; Kluge, R.L. Seed banks of invasive Australian Acacia species in South Africa: Role in invasiveness and options for management. Perspect. Plant Ecol. Evol. Syst. 2008, 10, 161–177. [Google Scholar] [CrossRef]
- Zhang, H.; Guan, D.; Song, M. Biomass and carbon storage of Eucalyptus and Acacia plantations in the Pearl River Delta, South China. For. Ecol. Manag. 2012, 277, 90–97. [Google Scholar] [CrossRef]
- Na, W.; Molong, S.; Lihai, W. Comparative Analysis on Calorific Value of Eucalyptus, Acacia mangium and Acacia crassicarpa. For. Eng. 2011, 4, 23–30. [Google Scholar]
- Feledyn-Szewczyk, B.; Matyka, M.; Staniak, M. Comparison of the Effect of Perennial Energy Crops and Agricultural Crops on Weed Flora Diversity. Agronomy 2019, 9, 695. [Google Scholar] [CrossRef] [Green Version]
- Sartori, F.; Lal, R.; Ebinger, M.H.; Parrish, D.J. Potential Soil Carbon Sequestration and CO2Offset by Dedicated Energy Crops in the USA. Crit. Rev. Plant Sci. 2006, 25, 441–472. [Google Scholar] [CrossRef]
- Chornesky, E.A.; Bartuska, A.M.; Aplet, G.H.; Britton, K.O.; Cummings-Carlson, J.; Davis, F.W.; Eskow, J.; Gordon, D.R.; Gottschalk, K.W.; Haack, R.A.; et al. Science Priorities for Reducing the Threat of Invasive Species to Sustainable Forestry. Bioscience 2005, 55, 335–348. [Google Scholar] [CrossRef] [Green Version]
- Yemshanov, D.; McKenney, D.W.; Pedlar, J.H.; Koch, F.H.; Cook, D. Towards an integrated approach to modelling the risks and impacts of invasive forest species. Environ. Rev. 2009, 17, 163–178. [Google Scholar] [CrossRef]
- Lubalega, T.K.; Gbawe, V.; KHASA, D.P.; RUEL, J.-C.; LEJOLY, J. Forest Regeneration of the Bateke Plateau Savannahs from Acacia Auriculformis Plantations in the Democratic Republic of the Congo. Forest 2017, 13, 21–30. [Google Scholar]
- Rodríguez-Echeverría, S.; Crisóstomo, J.A.; Nabais, C.; Freitas, H. Belowground mutualists and the invasive ability of Acacia longifolia in coastal dunes of Portugal. Biol. Invasions 2009, 11, 651–661. [Google Scholar] [CrossRef] [Green Version]
- Santos, A.; Simoes, R.; Tavares, M.L.C. Variation of some wood macroscopic properties along the stem of Acacia melanoxylon R. Br. adult trees in Portugal. For. Syst. 2013, 22, 463–470. [Google Scholar] [CrossRef]
- Martins, F.; Alegria, C.; Artur, G. Mapping invasive alien Acacia dealbata Link using ASTER multispectral imagery: A case study in central-eastern of Portugal. For. Syst. 2016, 25, e078. [Google Scholar] [CrossRef] [Green Version]
- Nunes, L.J.R.; Meireles, C.I.R.; Gomes, C.J.P.; Ribeiro, N.M.C.A. Historical Development of the Portuguese Forest: The Introduction of Invasive Species. Forests 2019, 10, 974. [Google Scholar] [CrossRef] [Green Version]
- Vaz, A.S.; Ribeiro, J.; Honrado, J.P.; Vicente, J.R. Stakeholders’ perceptions towards non-native acacias and implications for their management in Portugal. Forestry 2019, 93, 557–566. [Google Scholar] [CrossRef]
- Rodrigues, A.; Loureiro, L.; Nunes, L. Torrefaction of woody biomasses from poplar SRC and Portuguese roundwood: Properties of torrefied products. Biomass Bioenergy 2018, 108, 55–65. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Sample no. | Distance (m) | Direction (°) | Direction (rad) | DBH1 (cm) | DBH2 (cm) | DBHaverage (cm) | DBH Class |
|---|---|---|---|---|---|---|---|---|
| A.d. | 1 | 0.73 | 4 | 0.0698 | 4.7 | 4.8 | 4.75 | 5 |
| A.d. | 2 | 1.8 | 30 | 0.5236 | 5 | 4.6 | 4.8 | 5 |
| A.d. | 3 | 3.3 | 5 | 0.0873 | 10.6 | 9.9 | 10.25 | 15 |
| A.d. | 4 | 4.1 | 5 | 0.0873 | 4.4 | 3.7 | 4.05 | 5 |
| A.d. | 5 | 3.9 | 10 | 0.1745 | 7.1 | 6.7 | 6.9 | 10 |
| A.d. | 6 | 4.8 | 60 | 1.0472 | 9.7 | 7.2 | 8.45 | 10 |
| A.u. | 7 | 4.5 | 85 | 1.4835 | ||||
| A.d. | 8 | 3.6 | 90 | 1.5708 | 30.4 | 31 | 30.7 | 35 |
| A.d. | 9 | 4.5 | 100 | 1.7453 | 6.8 | 6.8 | 6.8 | 10 |
| A.d. | 10 | 5.1 | 85 | 1.4835 | 4.3 | 4.2 | 4.25 | 5 |
| A.d. | 11 | 5.1 | 90 | 1.5708 | 5 | 5.6 | 5.3 | 10 |
| A.d. | 12 | 5.2 | 97 | 1.693 | 4.2 | 4.1 | 4.15 | 5 |
| A.d. | 13 | 5.2 | 98 | 1.7104 | 6.5 | 6.8 | 6.65 | 10 |
| A.d. | 14 | 5.2 | 99 | 1.7279 | 3.5 | 3.5 | 3.5 | 5 |
| A.d. | 15 | 5.6 | 97 | 1.693 | 3.7 | 3.7 | 3.7 | 5 |
| A.d. | 16 | 5.2 | 100 | 1.7453 | 8.1 | 8 | 8.05 | 10 |
| A.d. | 17 | 4.4 | 101 | 1.7628 | 5.4 | 5.6 | 5.5 | 10 |
| A.u. | 18 | 4.2 | 100 | 1.7453 | 5 | |||
| A.d. | 19 | 5.1 | 115 | 2.0071 | 14.6 | 15.2 | 14.9 | 15 |
| A.d. | 20 | 5.3 | 126 | 2.1991 | 12.5 | 12.2 | 12.35 | 15 |
| A.d. | 21 | 4.2 | 117 | 2.042 | 5.9 | 5.7 | 5.8 | 10 |
| A.d. | 22 | 5.45 | 130 | 2.2689 | 4.6 | 4.5 | 4.55 | 5 |
| A.d. | 23 | 5.4 | 129 | 2.2515 | 3.4 | 3.6 | 3.5 | 5 |
| P.p. | 24 | 5.35 | 130 | 2.2689 | ||||
| A.d. | 25 | 5.6 | 138 | 2.4086 | 8.2 | 9 | 8.6 | 10 |
| A.d. | 26 | 5.3 | 141 | 2.4609 | 17.4 | 16.7 | 17.05 | 20 |
| A.d. | 27 | 5.4 | 143 | 2.4958 | 4.3 | 4.1 | 4.2 | 5 |
| A.d. | 28 | 5.6 | 155 | 2.7053 | 6.7 | 6.4 | 6.55 | 10 |
| A.d. | 29 | 5.3 | 160 | 2.7925 | 4.4 | 4.5 | 4.45 | 5 |
| A.d. | 30 | 5.3 | 162 | 2.8274 | 8.3 | 8.4 | 8.35 | 10 |
| A.d. | 31 | 2.9 | 186 | 3.2463 | 17.9 | 18.2 | 18.05 | 20 |
| A.d. | 32 | 3.5 | 185 | 3.2289 | 8.5 | 8.4 | 8.45 | 10 |
| A.d. | 33 | 1.9 | 196 | 3.4208 | 16.7 | 16.7 | 16.7 | 20 |
| A.d. | 34 | 2.3 | 210 | 3.6652 | 5.2 | 5.3 | 5.25 | 10 |
| A.d. | 35 | 1.8 | 220 | 3.8397 | 5.2 | 4.7 | 4.95 | 5 |
| A.d. | 36 | 1.8 | 235 | 4.1015 | 10.1 | 10 | 10.05 | 15 |
| A.d. | 37 | 5.4 | 222 | 3.8746 | 16.6 | 15.2 | 15.9 | 20 |
| A.d. | 38 | 5 | 230 | 4.0143 | 3.4 | 3.3 | 3.35 | 5 |
| A.d. | 39 | 5.6 | 242 | 4.2237 | 13 | 13 | 13 | 15 |
| A.d. | 40 | 5.3 | 244 | 4.2586 | 22.2 | 21.6 | 21.9 | 25 |
| A.d. | 41 | 4.1 | 245 | 4.2761 | 11.7 | 10.8 | 11.25 | 15 |
| A.d. | 42 | 4 | 258 | 4.5029 | 4.3 | 4.4 | 4.35 | 5 |
| A.d. | 43 | 4.8 | 272 | 4.7473 | 4.6 | 5.2 | 4.9 | 5 |
| A.d. | 44 | 4.4 | 290 | 5.0615 | 4.5 | 4 | 4.25 | 5 |
| A.d. | 45 | 4.2 | 295 | 5.1487 | 10.8 | 10.2 | 10.5 | 15 |
| A.d. | 46 | 3.9 | 310 | 5.4105 | 16.7 | 17.8 | 17.25 | 20 |
| A.d. | 47 | 3.9 | 320 | 5.5851 | 14.5 | 16.4 | 15.45 | 20 |
| A.d. | 48 | 4.1 | 340 | 5.9341 | 14.2 | 15.6 | 14.9 | 15 |
| A.d. | 49 | 5.2 | 0 | 0 | 4.6 | 4.8 | 4.7 | 5 |
| A.d. | 50 | 5.4 | 2 | 0.0349 | 14.3 | 15.4 | 14.85 | 15 |
| A.d. | 51 | 5.5 | 5 | 0.0873 | 3.6 | 3.2 | 3.4 | 5 |
| A.d. | 52 | 5.6 | 20 | 0.3491 | 7.4 | 8.7 | 8.05 | 10 |
| DBH Class | Quantity of Trees | Identification of Selected Trees |
|---|---|---|
| Class 5 | 19 | 5, 17, 29 |
| Class 10 | 14 | 3, 6, 16, 30 |
| Class 15 | 9 | 19, 26, 33 |
| Class 20 | 5 | |
| Class 25 | 1 | |
| Class 35 | 1 | 8 |
| Sample no. | Growth Starting Year | Age (years) | Ctotal (%) | ww (kg) | wbrl (kg) | wa (kg) | Productivity (kg∙year−1) |
|---|---|---|---|---|---|---|---|
| 3 | 2008 | 10 | 49.30 | 52.5 | 2.1 | 54.6 | 5 |
| 5 | 2013 | 5 | 48.70 | 23.6 | 5.2 | 28.8 | 6 |
| 6 | 2004 | 14 | 48.30 | 29.7 | 3.8 | 33.5 | 2 |
| 8 | 1998 | 20 | 49.10 | 578.9 | 15.9 | 594.8 | 30 |
| 16 | 2008 | 10 | 48.70 | 45 | 2.8 | 47.8 | 5 |
| 17 | 2009 | 9 | 48.40 | 14.2 | 4.3 | 18.5 | 2 |
| 19 | 2006 | 12 | 48.90 | 145.8 | 21 | 166.8 | 14 |
| 26 | 2004 | 14 | 50.40 | 223.6 | 25.5 | 249.1 | 18 |
| 29 | 2012 | 6 | 48.70 | 3.6 | 0.5 | 4.1 | 1 |
| 30 | 2009 | 9 | 48.80 | 43.5 | 2 | 45.5 | 5 |
| 33 | 2004 | 14 | 48.50 | 205 | 18.9 | 223.9 | 16 |
| Sample no. | Ctotal (%) | Moisture on Cut (%) | wtotal (kg) | dw (kg) | wcarbon (kg) | wCO2 (kg) | ARCS (kg∙year−1) |
|---|---|---|---|---|---|---|---|
| 3 | 49.30 | 48.30 | 54.60 | 28.23 | 13.92 | 51.07 | 5.11 |
| 5 | 48.70 | 48.65 | 28.80 | 14.79 | 7.20 | 26.43 | 5.29 |
| 6 | 48.30 | 42.10 | 33.50 | 19.40 | 9.37 | 34.38 | 2.46 |
| 8 | 49.10 | 38.30 | 594.80 | 366.99 | 180.19 | 661.31 | 33.07 |
| 16 | 48.70 | 44.12 | 47.80 | 26.71 | 13.01 | 47.74 | 4.77 |
| 17 | 48.40 | 39.74 | 18.50 | 11.15 | 5.40 | 19.80 | 2.20 |
| 19 | 48.90 | 40.53 | 166.80 | 99.20 | 48.51 | 178.02 | 14.84 |
| 26 | 50.40 | 38.56 | 249.10 | 153.05 | 77.14 | 283.09 | 20.22 |
| 29 | 48.70 | 36.18 | 4.10 | 2.62 | 1.27 | 4.68 | 0.78 |
| 30 | 48.80 | 45.71 | 45.50 | 24.70 | 12.05 | 44.24 | 4.92 |
| 33 | 48.50 | 42.78 | 223.90 | 128.12 | 62.14 | 228.04 | 16.29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nunes, L.J.R.; Raposo, M.A.M.; Meireles, C.I.R.; Pinto Gomes, C.J.; Almeida Ribeiro, N.M.C. Carbon Sequestration Potential of Forest Invasive Species: A Case Study with Acacia dealbata Link. Resources 2021, 10, 51. https://doi.org/10.3390/resources10050051
Nunes LJR, Raposo MAM, Meireles CIR, Pinto Gomes CJ, Almeida Ribeiro NMC. Carbon Sequestration Potential of Forest Invasive Species: A Case Study with Acacia dealbata Link. Resources. 2021; 10(5):51. https://doi.org/10.3390/resources10050051
Chicago/Turabian StyleNunes, Leonel J. R., Mauro A. M. Raposo, Catarina I. R. Meireles, Carlos J. Pinto Gomes, and Nuno M. C. Almeida Ribeiro. 2021. "Carbon Sequestration Potential of Forest Invasive Species: A Case Study with Acacia dealbata Link" Resources 10, no. 5: 51. https://doi.org/10.3390/resources10050051