Understanding the Unique Role of Phospholipids in the Lubrication of Natural Joints: An Interfacial Tension Study

 ,
,
Abstract
:1. Introduction
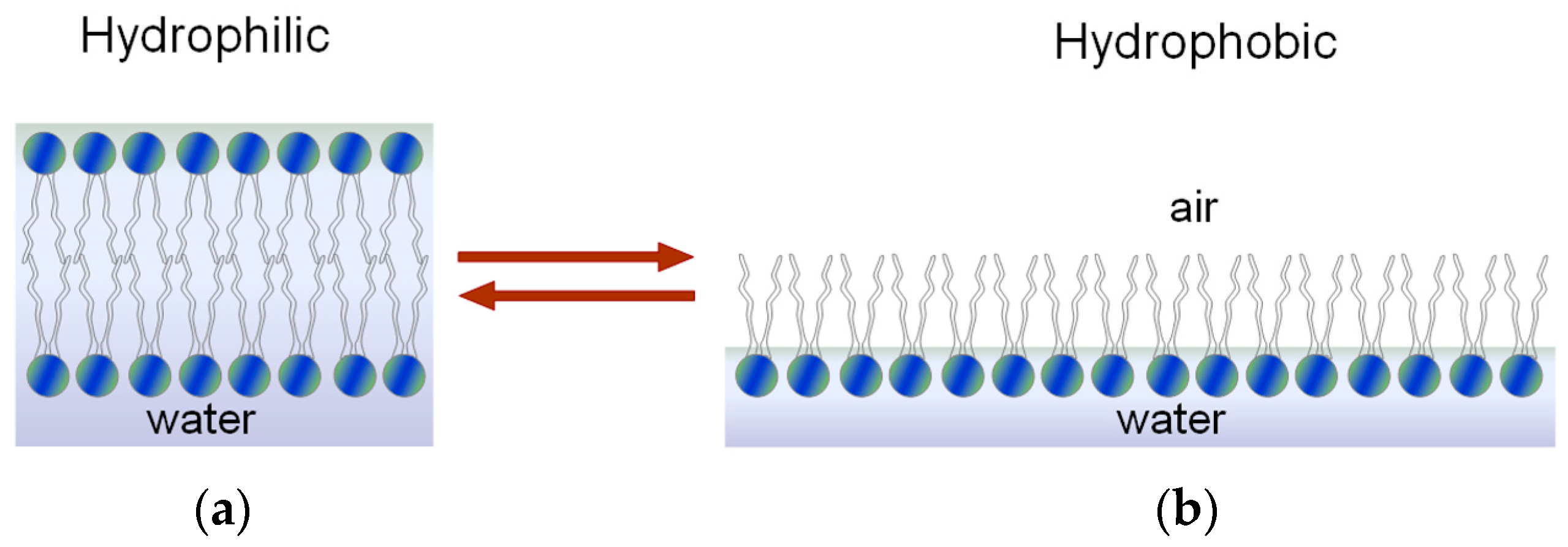
Hydrophilic and Hydrophobic Character of the Phospholipid Membrane
2. Materials and Methods
2.1. The Interfacial Tension Experiments
2.2. Friction Coefficient versus Charge Density of the Cartilage Surface
2.3. Friction Test in Universal Buffer Solutions (pH 2.0–9.5)
3. Results and Discussion
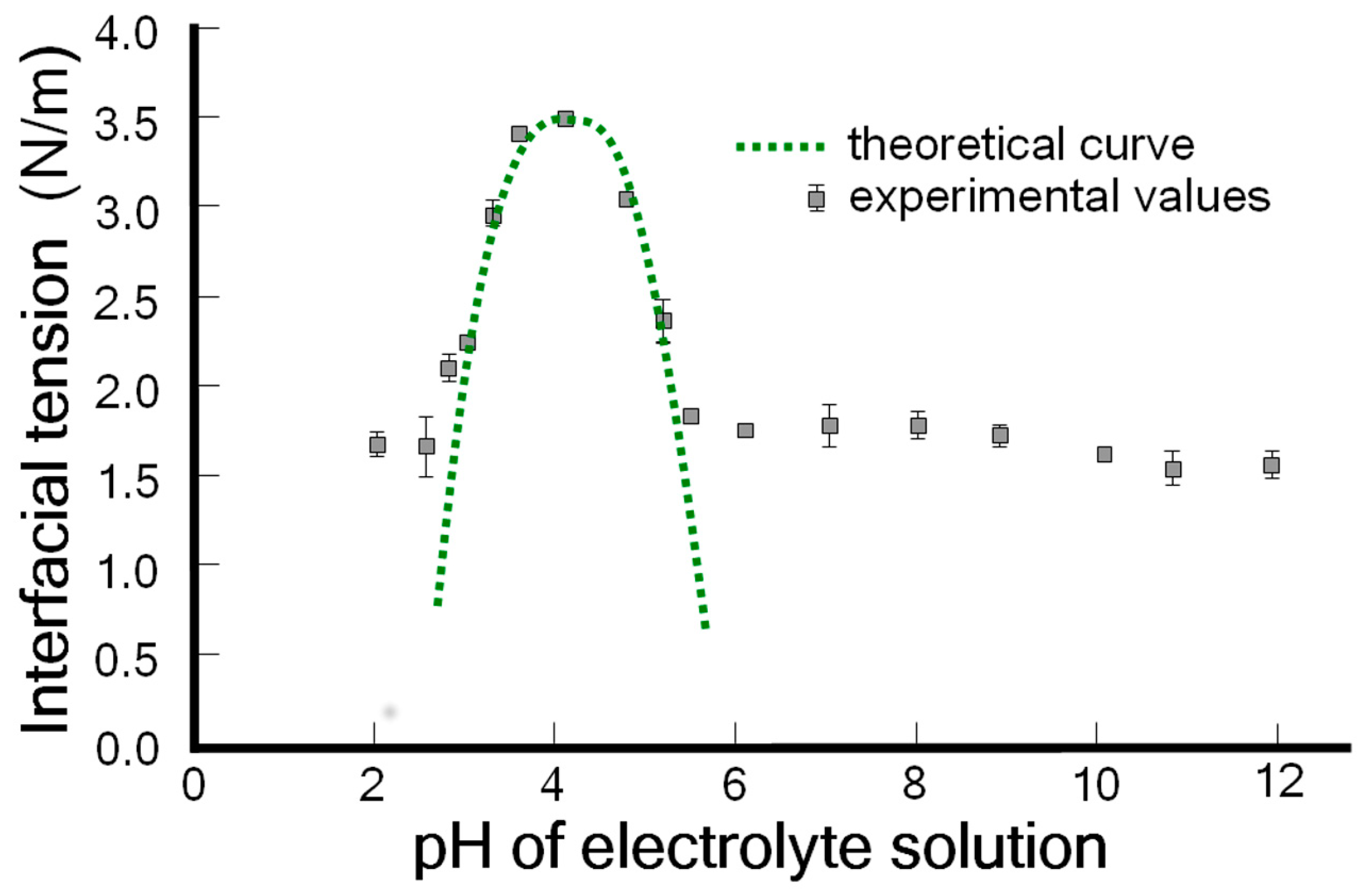
3.1. The Amphoteric Character of the Phospholipid Bilayer Surface of Spherical Lipid Bilayers Formed from Phosphatidylcholine
3.2. Hyaluronic Acid/Phosphatidylcholine Bilayer Interactions
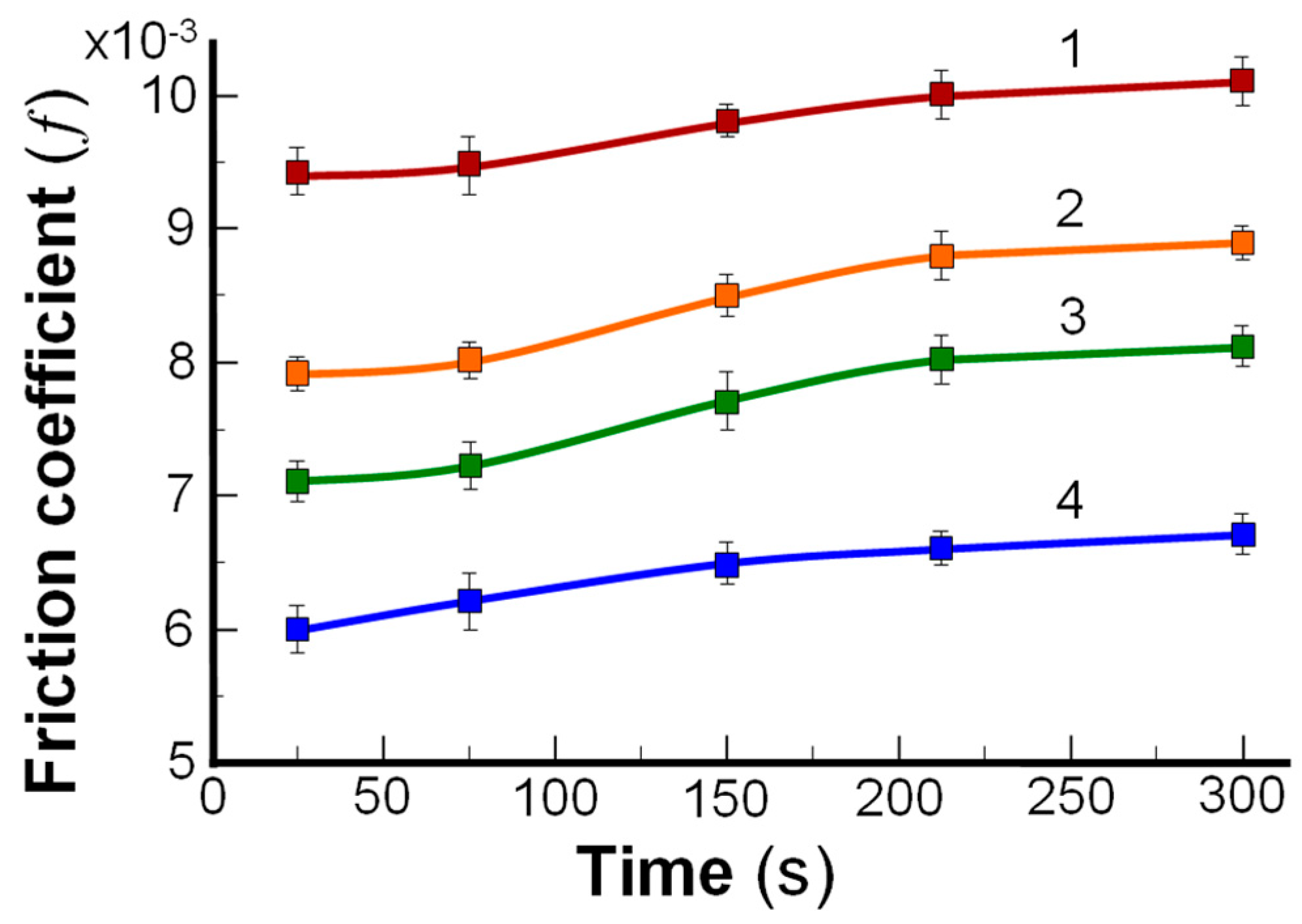
3.3. Lamellar-Repulsive Lubrication Mechanism between Cartilage/Cartilage Surface and Friction on Amphoteric Charged Surfaces
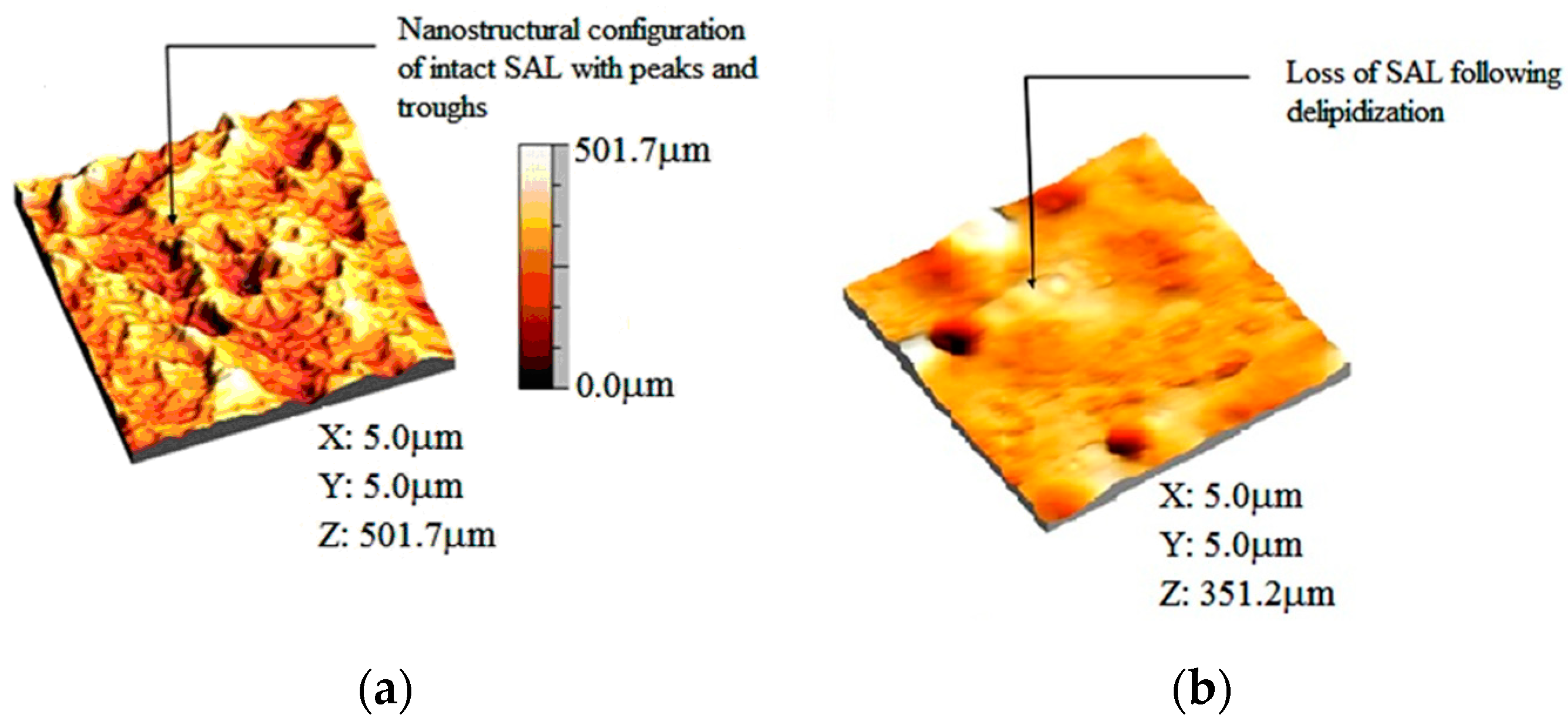
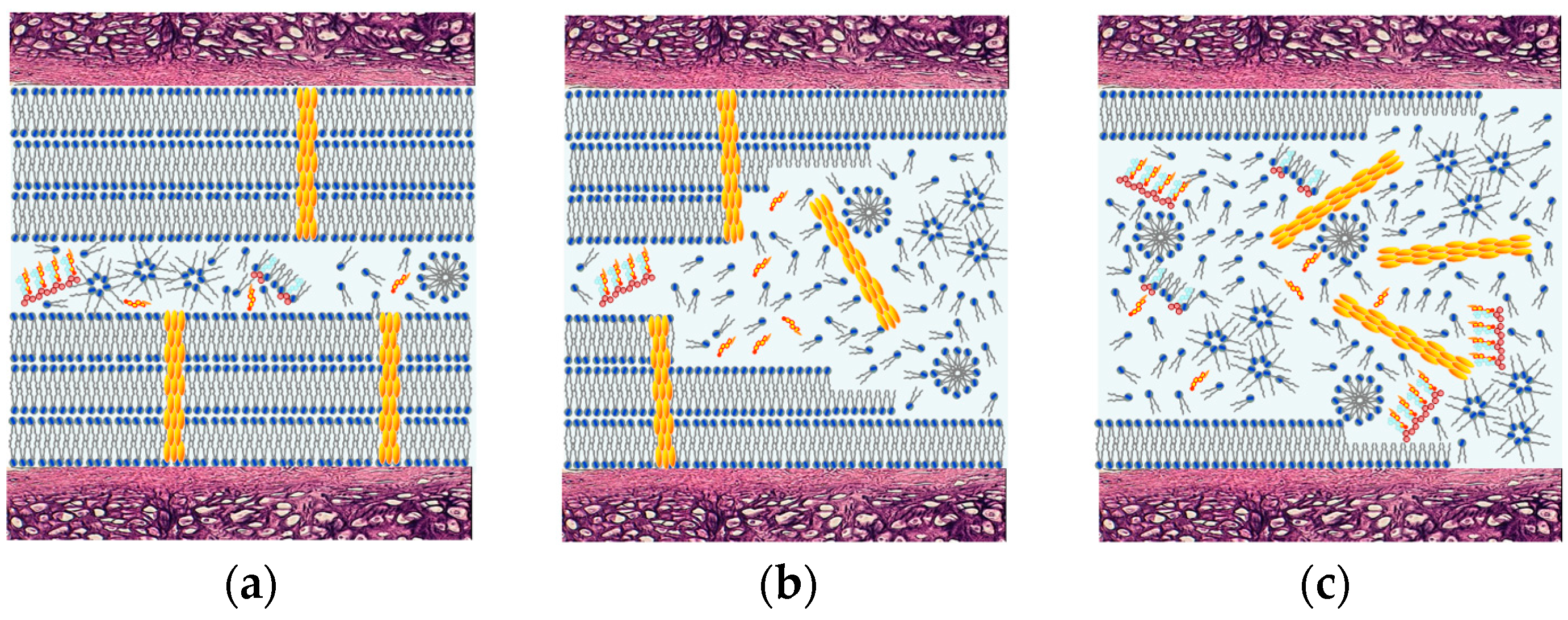
3.4. Deactivation of a Surface-Active Phospholipid Bilayer
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Schmitz, G.; Muller, G. Structure and function of lamellar bodies, lipid-protein complexes involved in storage and secretion of cellular lipids. J. Lipid Res. 1991, 32, 1539–1570. [Google Scholar] [PubMed]
- Hills, B.A. Surface-active phospholipid: A Pandora’s box of clinical applications, Part II Barrier and lubricating properties. J. Intern. Med. 2002, 32, 242–251. [Google Scholar] [CrossRef]
- Pawlak, Z.; Petelska, A.D.; Urbaniak, W.; Yusuf, K.Q.; Oloyede, A. Relationship between wettability and lubrication characteristics of the surfaces of contacting phospholipid-based membranes. Cell Biochem. Biophys. 2013, 65, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Mocny, P.; Klok, H.A. Tribology of surface-grafted polymer brushes. Mol. Syst. Des. Eng. 2016, 1, 141–154. [Google Scholar] [CrossRef]
- Yang, W.; Zhou, F. Polymer brushes for antibiofouling and lubrication. Biosurf. Biotribol. 2017, 3, 97–114. [Google Scholar] [CrossRef]
- Krishnamoorthy, M.; Hakobyan, S.; Ramstedt, M.; Gautrot, J.E. Surface-initiated polymer brushes in the biomedical field: Applications in membrane science, biosensing, cell culture, regenerative medicine and antibacterial coatings. Chem. Rev. 2014, 114, 10976–11026. [Google Scholar] [CrossRef]
- Samaroo, K.J.; Tan, M.; Eguiluz, R.C.A.; Gourdon, D.; Putnam, D.; Bonassar, L.J. Tunable lubricin-mimetics for boundary lubrication of cartilage. Biotribology 2017, 9, 18–23. [Google Scholar] [CrossRef]
- Singh, A.; Corvelli, M.; Unterman, S.A.; Wepasnick, K.A.; McDonnell, P.; Elisseeff, J.H. Enhanced lubrication on tissue and biomaterial surfaces through peptide-mediated binding of hyaluronic acid. Nat. Mater. 2014, 13, 988–995. [Google Scholar] [CrossRef]
- Chen, M.; Briscoe, W.H.; Armes, S.P.; Klein, J. Lubrication at physiological pressures by polyzwitterionic brushes. Science 2009, 323, 1698–1701. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Liu, Z.; Li, N.; Wang, X.; Zhou, F.; Liu, W. Hairy polyelectrolyte brushes-grafted thermosensitive microgels as artificial synovial fluid for simultaneous biomimetic lubrication and arthritis treatment. Appl. Mater. Interfaces 2014, 6, 20452–20463. [Google Scholar] [CrossRef]
- Ishihara, K. Highly lubricated polymer interfaces for advanced artificial hip joints through biomimetic design. Polym. J. 2015, 47, 585–597. [Google Scholar] [CrossRef]
- Nie, G.; Li, G.; Wang, L.; Zhang, X. Nanocomposites of polymer brush and inorganic nanoparticles: Preparation, characterization and application. Polym. Chem. 2016, 7, 753–769. [Google Scholar] [CrossRef]
- Bayer, I.S. Advances in tribology of lubricin and lubricin-like synthetic polymer nanostructures. Lubricants 2018, 6, 30. [Google Scholar] [CrossRef]
- Chappuis, J.; Sherman, A.; Neumann, A.W. Surface tension of animal cartilage as it relates to friction in joints. Ann. Biomed. Eng. 1983, 11, 435–449. [Google Scholar] [CrossRef]
- Hills, B.A. Boundary lubrication in vivo. Proc. Inst. Mech. Eng. H 2000, 214, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Hills, B.A. Oligolamellar lubrication of joints by surface-active phospholipid. J. Rheumatol. 1989, 16, 82–91. [Google Scholar]
- Britton, H.T.K.; Robinson, R.A. Universal buffer solutions and the dissociation constant of veronal. J. Chem. Soc. 1931, 1456–1462. [Google Scholar] [CrossRef]
- Petelska, A.D.; Figaszewski, Z. Effect of pH on the interfacial tension of lipid bilayer membrane. Biophys. J. 2000, 78, 812–817. [Google Scholar] [CrossRef]
- Adamson, A. Physical Chemistry of Surfaces; A Wiley-Interscience Publication, John Wiley & Sons, Inc.: New York, NY, USA, 1960; ISBN 0471610194. [Google Scholar]
- Petelska, A.D.; Figaszewski, Z.A. Interfacial tension of the two-component bilayer lipid membrane modelling of cell membrane. Bioelectrochem. Bioenerget. 1998, 46, 199–204. [Google Scholar] [CrossRef]
- Petelska, A.D.; Naumowicz, M.; Figaszewski, Z. Interfacial tension of the lipid membrane formed from lipid-fatty acid and lipid-amine systems. Bioeletrochemistry 2007, 70, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Mueller, P.; Rudin, D.O.; Tien, H.T.; Wescott, W.C. Methods for the formation of single bimolecular lipid membranes in aqueous solution. J. Phys. Chem. 1963, 67, 534–535. [Google Scholar] [CrossRef]
- Pawlak, Z.; Urbaniak, W.; Gadomski, A.; Yusuf, K.Q.; Afara, I.O.; Oloyede, A. The role of lamellate phospholipid bilayers in lubrication of joints. Acta Bioeng. Biomech. 2012, 14, 101–106. [Google Scholar] [PubMed]
- Sarma, A.V.; Powell, G.L.; LaBerge, M. Phospholipid composition of articular cartilage boundary lubricant. J. Orthop. Res. 2001, 19, 671–676. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, I.M.; Hills, B.A. Synovial surfactant: Lamellar bodies in type B synoviocytes and proteolipid in synovial fluid and the articular lining. Br. J. Rheumatol. 1996, 35, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, Z.; Urbaniak, W.; Oloyede, A. The relationship between friction and wettability in aqueous environment. Wear 2011, 271, 1745–1749. [Google Scholar] [CrossRef]
- Petelska, A.D.; Figaszewski, Z. Acid-base equilibria at interface separating electrolyte solution and lipid bilayer formed from phosphatidylcholine. Biophys. Chem. 2003, 104, 13–19. [Google Scholar] [CrossRef]
- Petelska, A.D.; Figaszewski, Z.A. Effect of pH on the interfacial tension of bilayer lipid membrane formed from phosphatidylcholine or phosphatidylserine. Biochim. Biophys. Acta 2002, 1561, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Petelska, A.D.; Figaszewski, Z.A. Interfacial tension of bilayer lipid membrane. Cent. Eur. J. Chem. 2012, 10, 16–26. [Google Scholar] [CrossRef]
- Linn, F.C.; Radin, E.I. Lubrication of animal joints III—The effect of certain chemical alterations of the cartilage and lubricant. Arthritis Rheumatol. 1968, 11, 674–682. [Google Scholar] [CrossRef]
- Kosińska, M.K.; Liebisch, G.A. A lipidomic study of phospholipid classes and species in human synovial fluid. Arthritis Rheumatol. 2013, 65, 2323–2333. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, Z.; Gadomski, A.; Sojka, M.; Urbaniak, W.; Bełdowski, P. The amphoteric effect on friction between the bovine cartilage/cartilage surfaces under slightlysheared hydration lubrication mode. Colloids Surf. B 2016, 146, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Petelska, A.D.; Beldowski, P.; Augé, W.K.; Casey, T.; Walczak, D.; Lemke, K.; Gadomski, A. Hyaluronic acid and phospholipid interactions useful for repaired articular cartilage surfaces—A mini review toward tribological surgical adjuvants. Colloid Polym. Sci. 2017, 295, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Siódmiak, J.; Bełdowski, P.; Augé, W.K., II; Ledziński, D.; Śmigiel, S.; Gadomski, A. Molecular dynamic analysis of hyaluronic acid and phospholipid interaction in tribological surgical adjuvant design for osteoarthritis. Molecules 2017, 22, 1436. [Google Scholar] [CrossRef] [PubMed]
- Sorkin, R.; Kampf, N.; Zhu, L.; Klein, J. Hydration lubrication and shear-induced self-healing of lipid bilayer boundary lubricants in phosphatidylcholine dispersions. Soft Matter 2016, 12, 2773–2784. [Google Scholar] [CrossRef]
- Wang, M.; Liu, C.; Thormann, E.; Dédinaité, A. Hyaluronan and phospholipid association in biolubrication. Biomacromolecules 2013, 14, 4198–4206. [Google Scholar] [CrossRef] [PubMed]
- Gadomski, A.; Pawlak, Z.; Oloyede, A. Directed ion transport asvirtual cause of some facilitated friction-lubrication mechanism pre-vailing in articular cartilage. Tribol. Lett. 2008, 30, 83–90. [Google Scholar] [CrossRef]
- Pasquali-Ronchetti, I.; Quaglino, D.; Mori, G.; Bacchelli, B. Hyaluronan-phospholipid interactions. J. Struct. Biol. 1997, 120, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, Z.; Urbaniak, W.; Hagner-Derengowska, M.; Hagner, W. The probable explanation for the low friction of natural joints. Cell Biochem. Biophys. 2015, 71, 1615–1621. [Google Scholar] [CrossRef] [PubMed]
- Augé, W.K. Translamellar dynamics of the surface activephospholipid layer: Protonwave propagation throughsphingomyelin raft channels. Cartilage 2013, 4, 131S. [Google Scholar]
- Gadomski, A.; Rubí, J.M.; Łuczka, J.; Ausloos, M. On temperature- and space-dimension dependent matter agglomeration in a mature growing stage. Chem. Phys. 2005, 310, 153–161. [Google Scholar] [CrossRef]
- Verberne, G.; Schroeder, A.; Halperin, G.; Barenholz, Y.; Etsion, I. Liposomes as potential biolubricant additives for wear reduction in human synovial joints. Wear 2010, 268, 1037–1042. [Google Scholar] [CrossRef]
- Yu, C.-H.; Groves, J.T. Engineering supported membranes for cell biology. Med. Biol. Eng. Comput. 2010, 48, 955–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Laat, B.; de Groot, P.G. Autoantibodies directed against domain I of Beta2-Glycoprotein I. Curr. Rheumatol. Rep. 2011, 13, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Sodin-Šemrl, S.; Frank, M.; Ambrožič, A.; Pavlič, J.; Šuštar, V.; Čučnik, S.; Božič, B.; Kveder, T.; Rozman, B. Advances in Planar Lipid Bilayers and Liposomes; Elsevier: Amsterdam, The Netherlands, 2008; Volume 8, pp. 243–273. [Google Scholar]
- Kondo, A.; Miyamoto, T.; Yonekawa, O.; Giessing, A.M.; Østerlund, E.C.; Jensen, N. Glycopeptide profiling of beta-2-glycoprotein I by mass spectrometry reveals attenuated sialylation in patients with antiphospholipid syndrome. J. Proteomics 2009, 73, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Tripodi, A.; de Groot, P.G.; Pengo, V. Antiphospholipid syndrome: Laboratory detection, mechanisms of action and treatment. J. Intern. Med. 2011, 270, 110–122. [Google Scholar] [CrossRef]
- Pawlak, Z.; Urbaniak, W.; Hagner-Derengowska, M.; Hagner, W. Lamellar slippage of bilayers—A hypothesis on low friction of natural joints. Biointerphases 2014, 9, 041004. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, Z.; Mrela, A.; Kaczmarek, M.; Cieszko, M.; Urbaniak, W. Natural joints: Boundary lubrication and antiphospholipid syndrome (APS). Biosystems 2019, 177, 44–47. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | Natural Articular Cartilage | Artificial Articular Cartilage |
|---|---|---|
| Wettability | Surfaces are hydrophobic, Δθ ≈ 0° | Surfaces with different hydrophobicity, Δθ = 22°–26° |
| Surface Property | Surface amorphous layer has a multi-bilayer structure and the surface is hydrophilic | A layer of protein adheres to the surface and this surface becomes hydrophilic |
| Friction | Ultra-low friction (f = 0.005) between tribopair with synovial fluid of 0.9% NaCl, lubricin and hyaluron, protein macromolecules, and phospholipid micelles, pH of ~7.4 | Low friction (f = 0.05–0.15) between tribopair with synovial fluid in the presence of 0.9% NaCl, lubricin and hyaluron, protein macromolecules, and phospholipid micelles, pH of ~7.2–7.4 |
| Degenerate Condition | Content of phospholipids is decreasing, phospholipids bilayers are uncertain or vanishing, wettability is lower (~40°) | Proteins more than phospholipids tend to adsorb strongly on soft and hard hydrophobic surfaces |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petelska, A.D.; Kazimierska-Drobny, K.; Janicka, K.; Majewski, T.; Urbaniak, W. Understanding the Unique Role of Phospholipids in the Lubrication of Natural Joints: An Interfacial Tension Study. Coatings 2019, 9, 264. https://doi.org/10.3390/coatings9040264
Petelska AD, Kazimierska-Drobny K, Janicka K, Majewski T, Urbaniak W. Understanding the Unique Role of Phospholipids in the Lubrication of Natural Joints: An Interfacial Tension Study. Coatings. 2019; 9(4):264. https://doi.org/10.3390/coatings9040264
Chicago/Turabian StylePetelska, Aneta D., Katarzyna Kazimierska-Drobny, Katarzyna Janicka, Tomasz Majewski, and Wiesław Urbaniak. 2019. "Understanding the Unique Role of Phospholipids in the Lubrication of Natural Joints: An Interfacial Tension Study" Coatings 9, no. 4: 264. https://doi.org/10.3390/coatings9040264