
Interaction of Cerium Oxide Nanoparticles and Ionic Cerium with Duckweed (Lemna minor L.): Uptake, Distribution, and Phytotoxicity

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Material Preparation and Characterization
2.2. Plant Cultivation and Exposure
2.3. Growth Related Parameters
2.4. Cell Death and SEM/TEM Observation
2.5. Ce and Mineral Elements Analysis
2.6. Elemental Mapping by μ-XRF
2.7. Speciation of Ce in Duckweed
2.8. Statistical Analysis
3. Results
3.1. Characterization of CeO2 NPs
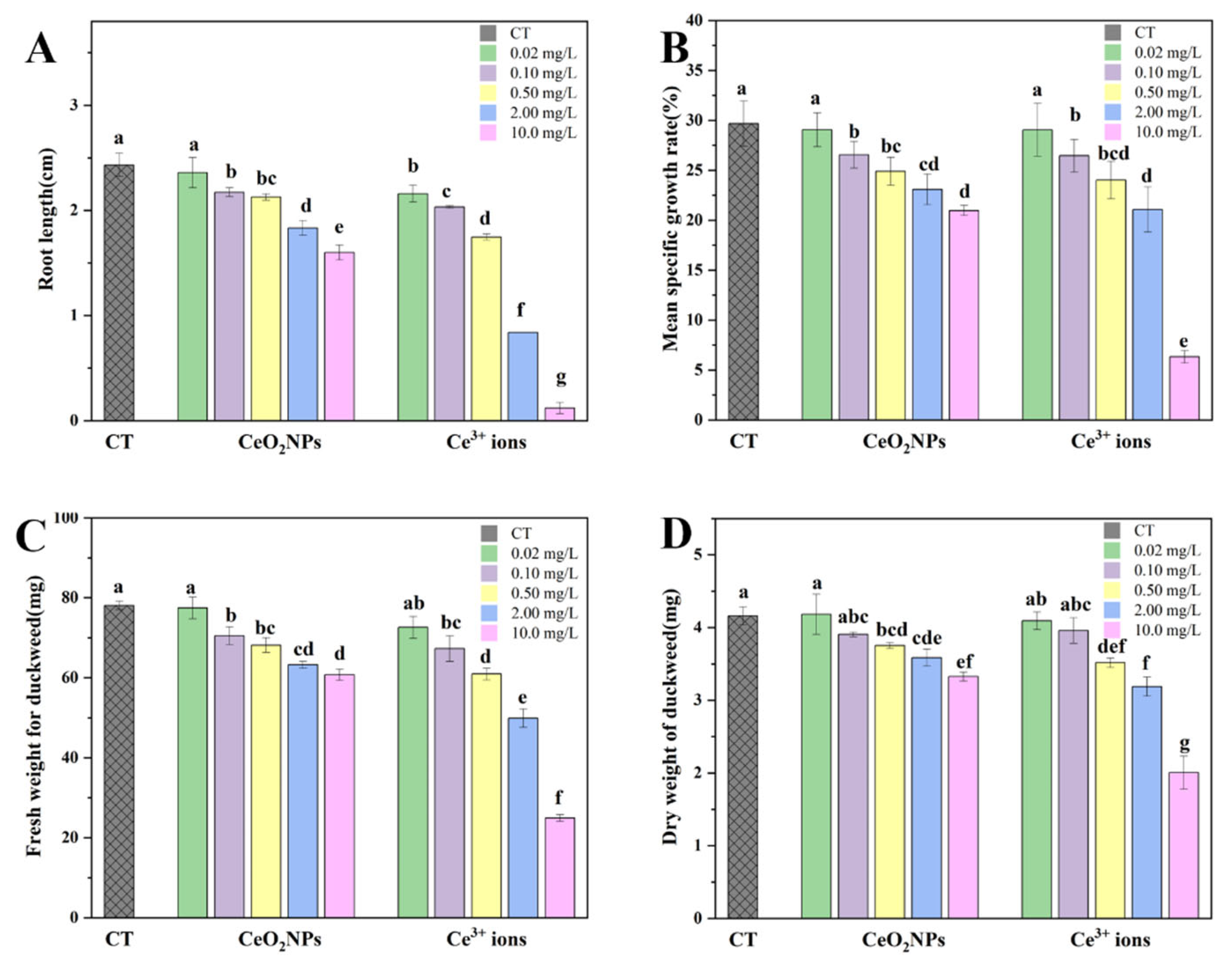
3.2. Effect of CeO2 NPs and Ce3+ Ions on the Growth of Duckweed
3.3. Pigments and Mineral Elements Analyses
3.4. Visualization and Quantification of Plant Cell Activity
3.5. SEM and TEM Observations
3.6. Elemental Mapping by μ-XRF
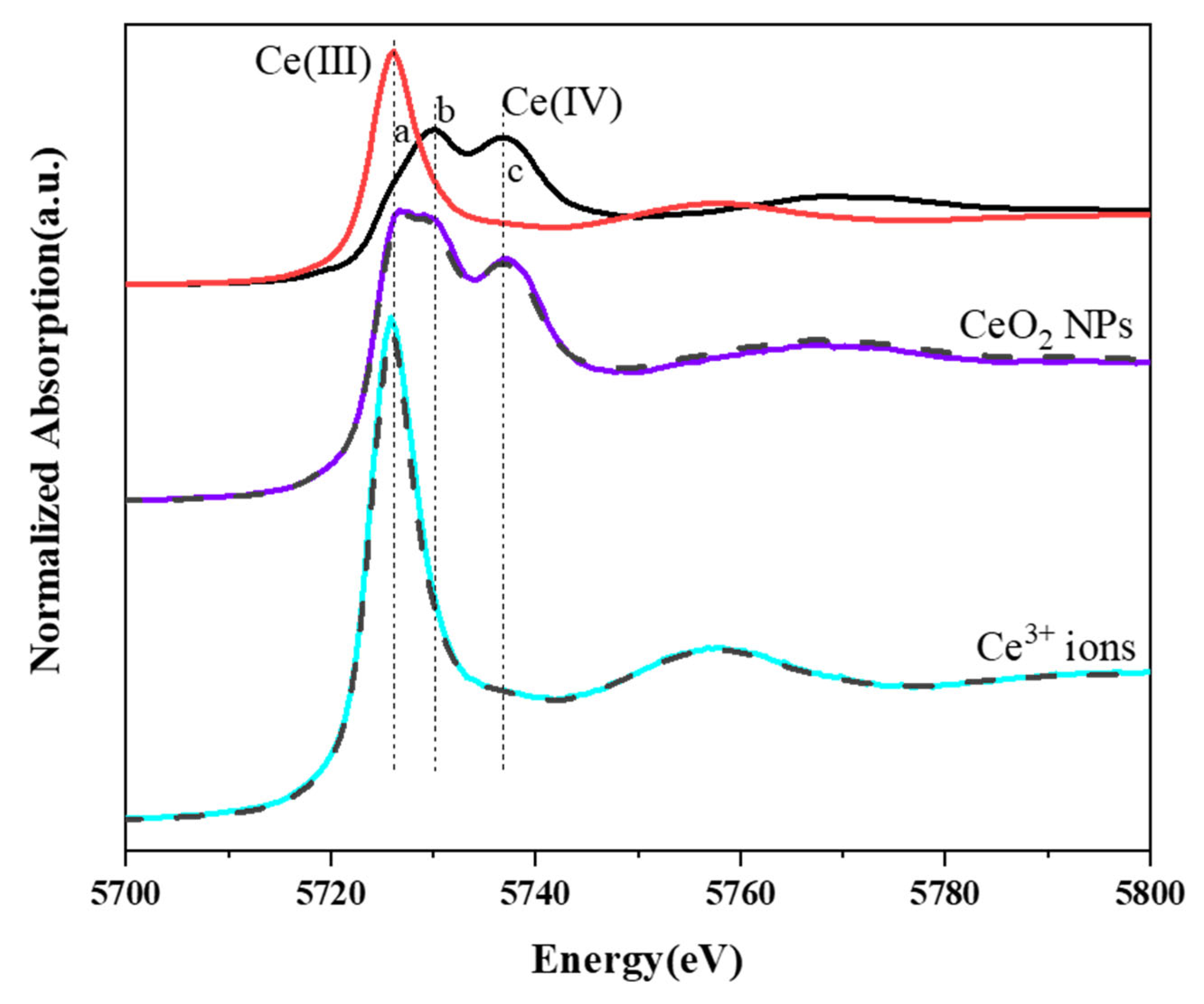
3.7. Contents and Speciation of Ce in Duckweed Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, Y.; Hou, F.; Tan, Y. CeO2 nanoplates with a hexagonal structure and their catalytic applications in highly selective hydrogenation of substituted nitroaromatics. Chem. Commun. 2012, 48, 2391–2393. [Google Scholar] [CrossRef]
- Johnson, A.C.; Park, B. Predicting contamination by the fuel additive cerium oxide engineered nanoparticles within the United Kingdom and the associated risks. Environ. Toxicol. Chem. 2012, 31, 2582–2587. [Google Scholar] [CrossRef] [PubMed]
- Celardo, I.; Pedersen, J.Z.; Traversa, E.; Ghibelli, L. Pharmacological potential of cerium oxide nanoparticles. Nanoscale 2011, 3, 1411–1420. [Google Scholar] [CrossRef] [PubMed]
- Keller, A.A.; McFerran, S.; Lazareva, A.; Suh, S. Global life cycle releases of engineered nanomaterials. J. Nanopart. Res. 2013, 15, 1–17. [Google Scholar]
- Xu, C.; Qu, X.G. Cerium oxide nanoparticle: A remarkably versatile rare earth nanomaterial for biological applications. NPG Asia Mater. 2014, 6, e90. [Google Scholar] [CrossRef]
- Zhao, X.C.; Yu, M.; Xu, D.; Liu, A.F.; Hou, X.W.; Hao, F.; Long, Y.M.; Zhou, Q.F.; Jiang, G.B. Distribution, Bioaccumulation, Trophic Transfer, and Influences of CeO2 Nanoparticles in a Constructed Aquatic Food Web. Environ. Sci. Technol. 2017, 51, 5205–5214. [Google Scholar] [CrossRef]
- Servin, A.D.; White, J.C. Nanotechnology in agriculture: Next steps for understanding engineered nanoparticle exposure and risk. Nanoimpact 2016, 1, 9–12. [Google Scholar] [CrossRef]
- Wojcieszek, J.; Jimenez-Lamana, J.; Bierla, K.; Ruzik, L.; Asztemborska, M.; Jarosz, M.; Szpunar, J. Uptake, translocation, size characterization and localization of cerium oxide nanoparticles in radish (Raphanus sativus L.). Sci. Total Environ. 2019, 683, 284–292. [Google Scholar] [CrossRef]
- Giese, B.; Klaessig, F.; Park, B.; Kaegi, R.; Steinfeldt, M.; Wigger, H.; von Gleich, A.; Gottschalk, F. Risks, Release and Concentrations of Engineered Nanomaterial in the Environment. Sci. Rep. 2018, 8, 1565. [Google Scholar] [CrossRef]
- Manier, N.; Bado-Nilles, A.; Delalain, P.; Aguerre-Chariol, O.; Pandard, P. Ecotoxicity of non-aged and aged CeO2 nanomaterials towards freshwater microalgae. Environ. Pollut. 2013, 180, 63–70. [Google Scholar] [CrossRef]
- Xie, C.; Ma, Y.; Zhang, P.; Zhang, J.; Li, X.; Zheng, K.; Li, A.; Wu, W.; Pang, Q.; He, X. Elucidating the origin of the toxicity of nano-CeO2 to Chlorella pyrenoidosa: The role of specific surface area and chemical composition. Environ. Sci. Nano 2021, 8, 1701. [Google Scholar]
- Radić, S.; Domijan, A.M.; Glavaš Ljubimir, K.; Maldini, K.; Ivešić, M.; Peharec Štefanić, P.; Krivohlavek, A. Toxicity of nanosilver and fumonisin B(1) and their interactions on duckweed (Lemna minor L.). Chemosphere 2019, 229, 86–93. [Google Scholar] [CrossRef]
- Rippner, D.A.; Lien, J.; Balla, H.; Guo, T.; Green, P.G.; Young, T.M.; Parikh, S.J. Surface modification induced cuprous oxide nanoparticle toxicity to duckweed at sub-toxic metal concentrations. Sci. Total Environ. 2020, 722, 137607. [Google Scholar] [CrossRef]
- Modlitbová, P.; Hlaváček, A.; Švestková, T.; Pořízka, P.; Šimoníková, L.; Novotný, K.; Kaiser, J. The effects of photon-upconversion nanoparticles on the growth of radish and duckweed: Bioaccumulation, imaging, and spectroscopic studies. Chemosphere 2019, 225, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Tarrahi, R.; Khataee, A.; Movafeghi, A.; Rezanejad, F.; Gohari, G. Toxicological implications of selenium nanoparticles with different coatings along with Se(4+) on Lemna minor. Chemosphere 2017, 181, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Lalau, C.M.; Simioni, C.; Vicentini, D.S.; Ouriques, L.C.; Mohedano, R.A.; Puerari, R.C.; Matias, W.G. Toxicological effects of AgNPs on duckweed (Landoltia punctata). Sci. Total Environ. 2020, 710, 136318. [Google Scholar] [CrossRef] [PubMed]
- Stegemeier, J.P.; Colman, B.P.; Schwab, F.; Wiesner, M.R.; Lowry, G.V. Uptake and Distribution of Silver in the Aquatic Plant Landoltia punctata (Duckweed) Exposed to Silver and Silver Sulfide Nanoparticles. Environ. Sci. Technol. 2017, 51, 4936–4943. [Google Scholar] [CrossRef]
- Lalau, C.M.; Mohedano, R.d.A.; Schmidt, É.C.; Bouzon, Z.L.; Ouriques, L.C.; dos Santos, R.W.; da Costa, C.H.; Vicentini, D.S.; Matias, W.G. Toxicological effects of copper oxide nanoparticles on the growth rate, photosynthetic pigment content, and cell morphology of the duckweed Landoltia punctata. Protoplasma 2015, 252, 221–229. [Google Scholar] [CrossRef]
- Yue, L.; Zhao, J.; Yu, X.; Lv, K.; Wang, Z.; Xing, B. Interaction of CuO nanoparticles with duckweed (Lemna minor L..): Uptake, distribution and ROS production sites. Environ. Pollut. 2018, 243, 543–552. [Google Scholar] [CrossRef]
- Zhang, Z.; He, X.; Zhang, H.; Ma, Y.; Zhang, P.; Ding, Y.; Zhao, Y. Uptake and distribution of ceria nanoparticles in cucumber plants. Metallomics 2011, 3, 816–822. [Google Scholar] [CrossRef]
- No, O.T. 221: Lemna sp. Growth Inhibition Test; OECD Guidelines for the Testing of Chemicals, Section; OECD: Paris, France, 2006; Volume 2, pp. 1–22. [Google Scholar]
- Baker, C.J.; Mock, N.M. An Improved Method for Monitoring Cell-Death in Cell-Suspension and Leaf Disc Assays Using Evans Blue. Plant Cell Tissue Organ Cult. 1994, 39, 7–12. [Google Scholar] [CrossRef]
- Dong, G.; Nkoh, J.N.; Hong, Z.N.; Dong, Y.; Lu, H.L.; Yang, J.; Pan, X.Y.; Xu, R.K. Phytotoxicity of Cu2+ and Cd2+ to the roots of four different wheat cultivars as related to charge properties and chemical forms of the metals on whole plant roots. Ecotoxicol. Environ. Saf. 2020, 196, 110545. [Google Scholar] [CrossRef] [PubMed]
- You, Y.; Liu, L.J.; Wang, Y.; Li, J.X.; Ying, Z.N.; Hou, Z.L.; Liu, H.J.; Du, S.T. Graphene oxide decreases Cd concentration in rice seedlings but intensifies growth restriction. J. Hazard. Mater. 2021, 417, 125958. [Google Scholar] [CrossRef]
- Liu, M.; Feng, S.; Ma, Y.; Xie, C.; He, X.; Ding, Y.; Zhang, J.; Luo, W.; Zheng, L.; Chen, D.; et al. Influence of Surface Charge on the Phytotoxicity, Transformation, and Translocation of CeO2 Nanoparticles in Cucumber Plants. ACS Appl. Mater. Interfaces 2019, 11, 16905–16913. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Ma, Y.; Zhang, Z.; He, X.; Li, Y.; Zhang, J.; Zheng, L.; Zhao, Y. Species-specific toxicity of ceria nanoparticles to Lactuca plants. Nanotoxicology 2015, 9, 1–8. [Google Scholar] [CrossRef]
- Cui, D.; Zhang, P.; Ma, Y.; He, X.; Li, Y.; Zhang, J.; Zhao, Y.; Zhang, Z. Effect of cerium oxide nanoparticles on asparagus lettuce cultured in an agar medium. Environ. Sci. Nano 2014, 1, 459–465. [Google Scholar] [CrossRef]
- Zhang, W.; Ebbs, S.D.; Musante, C.; White, J.C.; Gao, C.; Ma, X. Uptake and Accumulation of Bulk and Nanosized Cerium Oxide Particles and Ionic Cerium by Radish (Raphanus sativus L.). J. Agric. Food Chem. 2015, 63, 382–390. [Google Scholar] [CrossRef]
- Chaudhary, E.; Sharma, P. Effect of cadmium on nutrients concentration in duckweed: A case of Lemna minor and Lemna gibba. Environ. Sustain. 2021, 4, 183–190. [Google Scholar] [CrossRef]
- Morkunas, I.; Woźniak, A.; Mai, V.C.; Rucińska-Sobkowiak, R.; Jeandet, P. The role of heavy metals in plant response to biotic stress. Molecules 2018, 23, 2320. [Google Scholar] [CrossRef]
- Shtangeeva, I.; Ayrault, S.J.E.; Botany, E. Effects of Eu and Ca on yield and mineral nutrition of wheat (Triticum aestivum) seedlings. Environ. Exp. Bot. 2007, 59, 49–58. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B.; Mittra, B. Effects of salt on growth, ion accumulation, photosynthesis and leaf anatomy of the mangrove, Bruguiera parviflora. Trees-Struct. Funct. 2004, 18, 167–174. [Google Scholar] [CrossRef]
- Shen, H.; Ren, Q.G.; Mi, Y.; Shi, X.F.; Yao, H.Y.; Jin, C.Z.; Huang, Y.Y.; He, W.; Zhang, J.; Liu, B. Investigation of metal ion accumulation in Euglena gracilis by fluorescence methods. Nucl. Instrum. Methods Phys. Res. Sect. B-Beam Interact. Mater. At. 2002, 189, 506–510. [Google Scholar] [CrossRef]
- Miller, G.W.; Pushnik, J.C.; Welkie, G.W. Iron chlorosis, a world wide problem, the relation of chlorophyll biosynthesis to iron. J. Plant Nutr. 1984, 7, 1–22. [Google Scholar] [CrossRef]
- Terry, N.; Low, G. Leaf chlorophyll content and its relation to the intracellular localization of iron. J. Plant Nutr. 1982, 5, 301–310. [Google Scholar] [CrossRef]
- Hu, Z.; Richter, H.; Sparovek, G.; Schnug, E. Physiological and biochemical effects of rare earth elements on plants and their agricultural significance: A review. J. Plant Nutr. 2004, 27, 183–220. [Google Scholar] [CrossRef]
- Diatloff, E.; Smith, F.; Asher, C. Effects of Lanthanum and Cerium on the Growth and Mineral Nutrition of Corn and Mungbean. Ann. Bot. 2008, 101, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, P.; DeVay, J.; Meredith, C. Roles of water stress and phytotoxins in the development of Pierce’s disease of the grapevine. Physiol. Mol. Plant Pathol. 1988, 32, 1–15. [Google Scholar] [CrossRef]
- Zhao, J.; Ren, W.; Dai, Y.; Liu, L.; Wang, Z.; Yu, X.; Zhang, J.; Wang, X.; Xing, B. Uptake, Distribution, and Transformation of CuO NPs in a Floating Plant Eichhornia crassipes and Related Stomatal Responses. Environ. Sci. Technol. 2017, 51, 7686–7695. [Google Scholar] [CrossRef]
- Noori, A.; Ngo, A.; Gutierrez, P.; Theberge, S.; White, J.C. Silver nanoparticle detection and accumulation in tomato (Lycopersicon esculentum). J. Nanopart. Res. 2020, 22, 131. [Google Scholar] [CrossRef]
- Natarajan, L.; Jenifer, M.; Mukherjee, A. Eco-corona formation on the nanomaterials in the aquatic systems lessens their toxic impact: A comprehensive review. Environ. Res. 2021, 194, 110669. [Google Scholar] [CrossRef]
- Schwabe, F.; Schulin, R.; Rupper, P.; Rotzetter, A.; Stark, W.; Nowack, B. Dissolution and transformation of cerium oxide nanoparticles in plant growth media. J. Nanopart. Res. 2014, 16, 2668. [Google Scholar] [CrossRef]
- Zhang, P.; Ma, Y.; Zhang, Z.; He, X.; Zhang, J.; Guo, Z.; Tai, R.; Zhao, Y.; Chai, Z. Biotransformation of Ceria Nanoparticles in Cucumber Plants. ACS Nano 2012, 6, 9943–9950. [Google Scholar] [CrossRef] [PubMed]
- Schwabe, F.; Tanner, S.; Schulin, R.; Rotzetter, A.C.; Stark, W.J.; von Quadt, A.; Nowack, B. Dissolved Cerium contributes to uptake of Ce in presence of differently sized CeO2-nanoparticles by three crop plants. Metallomics 2015, 7, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Escolà Casas, M.; Matamoros, V. Linking plant-root exudate changes to micropollutant exposure in aquatic plants (Lemna minor and Salvinia natans). A prospective metabolomic study. Chemosphere 2022, 287, 132056. [Google Scholar] [CrossRef]
- Wan, D.; Yang, J.; Wang, X.; Xiang, W.; Selvinsimpson, S.; Chen, Y. Wavelength-Dependent Photoreactivity of Root Exudates from Aquatic Plants under UV-LED Irradiation. ACS ES&T Water 2022, 2, 2613–2622. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments (mg/L) | Macroelements (μg/g·Dw) | Microelements (μg/g·Dw) | |||
|---|---|---|---|---|---|
| Ca | Mg | Fe | Mn | Zn | |
| 0 (CT) | 6209.1 ± 72.7 a | 2249.91 ± 70.6 a | 168.3 ± 14.3 a | 98.8 ± 3.9 a | 30.4 ± 5.3 a |
| CeO2 NPs | |||||
| 0.02 | 5464.7 ± 70.2 b | 2312.4 ± 143.3 a | 135.48 ± 5.5 b | 92.4± 0.7 b | 24.1 ± 9.6 ab |
| 0.1 | 5191.4 ± 114.7 bc | 2227.3 ± 78.2 a | 143.0 ± 35.4 ab | 85.5 ± 1.5 c | 19.8 ± 5.9 abc |
| 0.5 | 5234.3 ± 295.2 bc | 2343.4 ± 93.1 a | 96.1 ± 1.444 c | 83.4 ± 1.7 c | 11.4 ± 0.7 bc |
| 2 | 4948.5 ± 89.6 c | 2401.45 ± 17.1 a | 83.3 ± 6.8 c | 84.6 ± 1.7 c | 15.4 ± 4.3 bc |
| 10 | 4970.2 ± 172.0 c | 2265.7 ± 47.8 a | 83.8 ± 1.4 c | 86.2 ± 2.0 c | 7.3 ± 0.7 c |
| Ce3+ ions | |||||
| 0.02 | 5606.9 ± 146.3 b | 2188.8 ± 93.7 a | 125.3 ± 16.6 b | 87.8 ± 1.6 a | 12.3 ± 5.0 b |
| 0.1 | 5128.4 ± 211.8 c | 2254.2 ± 196.3 a | 99.7 ± 0.5 b | 88.4 ± 3.8 a | 22.9 ± 3.0 ab |
| 0.5 | 5262.0 ± 221.0 bc | 2273.8 ± 69.5 a | 108.6 ± 2.3 b | 82.6 ± 2.0 a | 13.2 ± 4.3 b |
| 2 | 5037.6 ± 132.1 c | 2652.4 ± 542.3 a | 124.6 ± 29.4 b | 100.6 ± 25.4 a | 21.3 ± 4.8 ab |
| 10 | 3502.1 ± 153.6 d | 929.2 ± 24.4 b | 133.9 ± 11.7 b | 51.5 ± 1.3 b | 17.2 ± 4.2 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Zhao, X.; Ma, Y.; Dai, W.; Song, Z.; Wang, Y.; Shen, J.; He, X.; Yang, F.; Zhang, Z. Interaction of Cerium Oxide Nanoparticles and Ionic Cerium with Duckweed (Lemna minor L.): Uptake, Distribution, and Phytotoxicity. Nanomaterials 2023, 13, 2523. https://doi.org/10.3390/nano13182523
Liu Y, Zhao X, Ma Y, Dai W, Song Z, Wang Y, Shen J, He X, Yang F, Zhang Z. Interaction of Cerium Oxide Nanoparticles and Ionic Cerium with Duckweed (Lemna minor L.): Uptake, Distribution, and Phytotoxicity. Nanomaterials. 2023; 13(18):2523. https://doi.org/10.3390/nano13182523
Chicago/Turabian StyleLiu, Yang, Xuepeng Zhao, Yuhui Ma, Wanqin Dai, Zhuda Song, Yun Wang, Jiaqi Shen, Xiao He, Fang Yang, and Zhiyong Zhang. 2023. "Interaction of Cerium Oxide Nanoparticles and Ionic Cerium with Duckweed (Lemna minor L.): Uptake, Distribution, and Phytotoxicity" Nanomaterials 13, no. 18: 2523. https://doi.org/10.3390/nano13182523