
Probing Polarity and pH Sensitivity of Carbon Dots in Escherichia coli through Time-Resolved Fluorescence Analyses
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.1.1. Preparation of CDs
2.1.2. Bacterial Sample Preparation
2.1.3. Preparation of Fluorescein Solution
2.2. Experimental Procedure
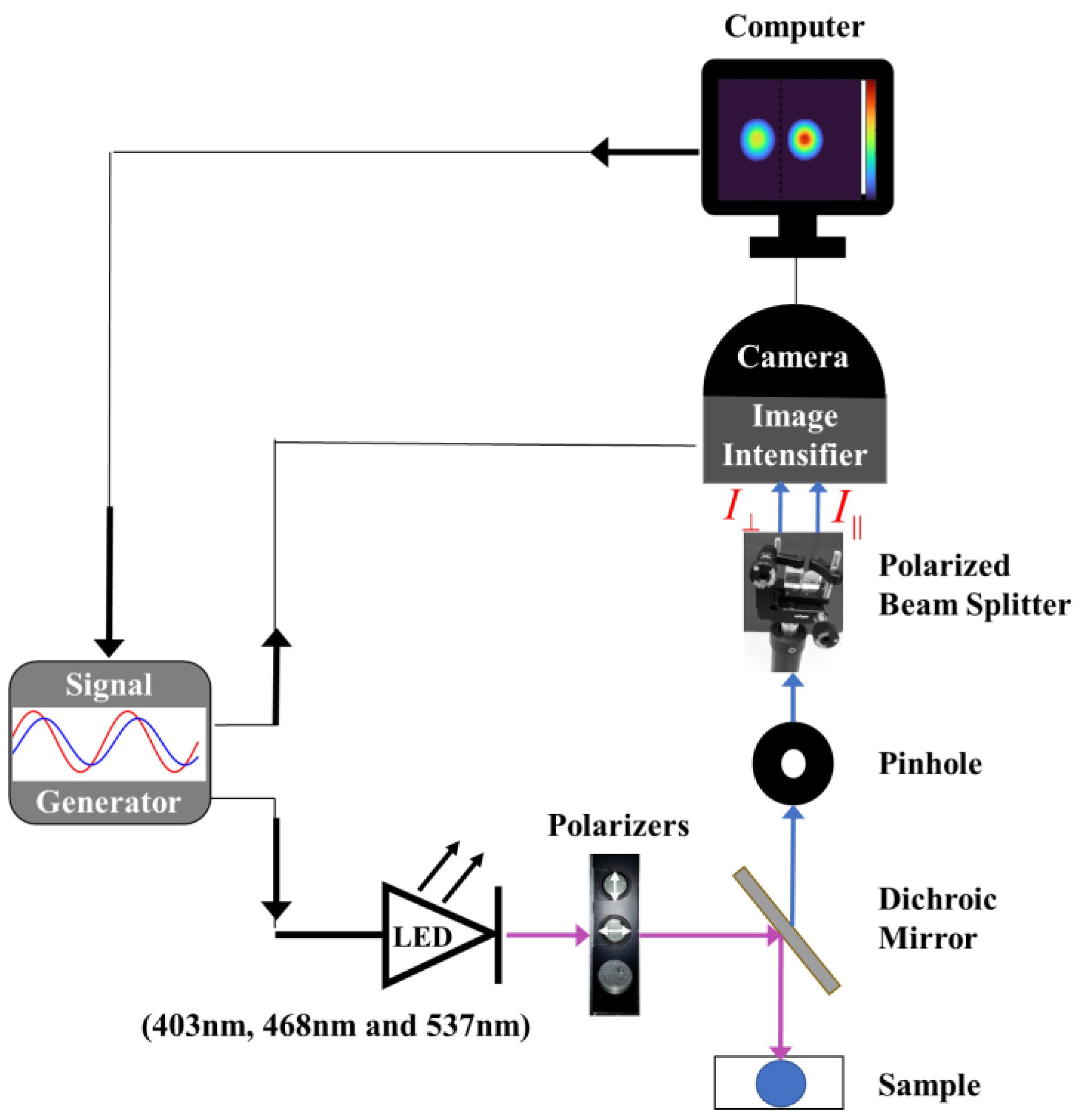
2.2.1. Experimental Setup
2.2.2. Calibration
2.2.3. Data Analysis
3. Results and Discussion
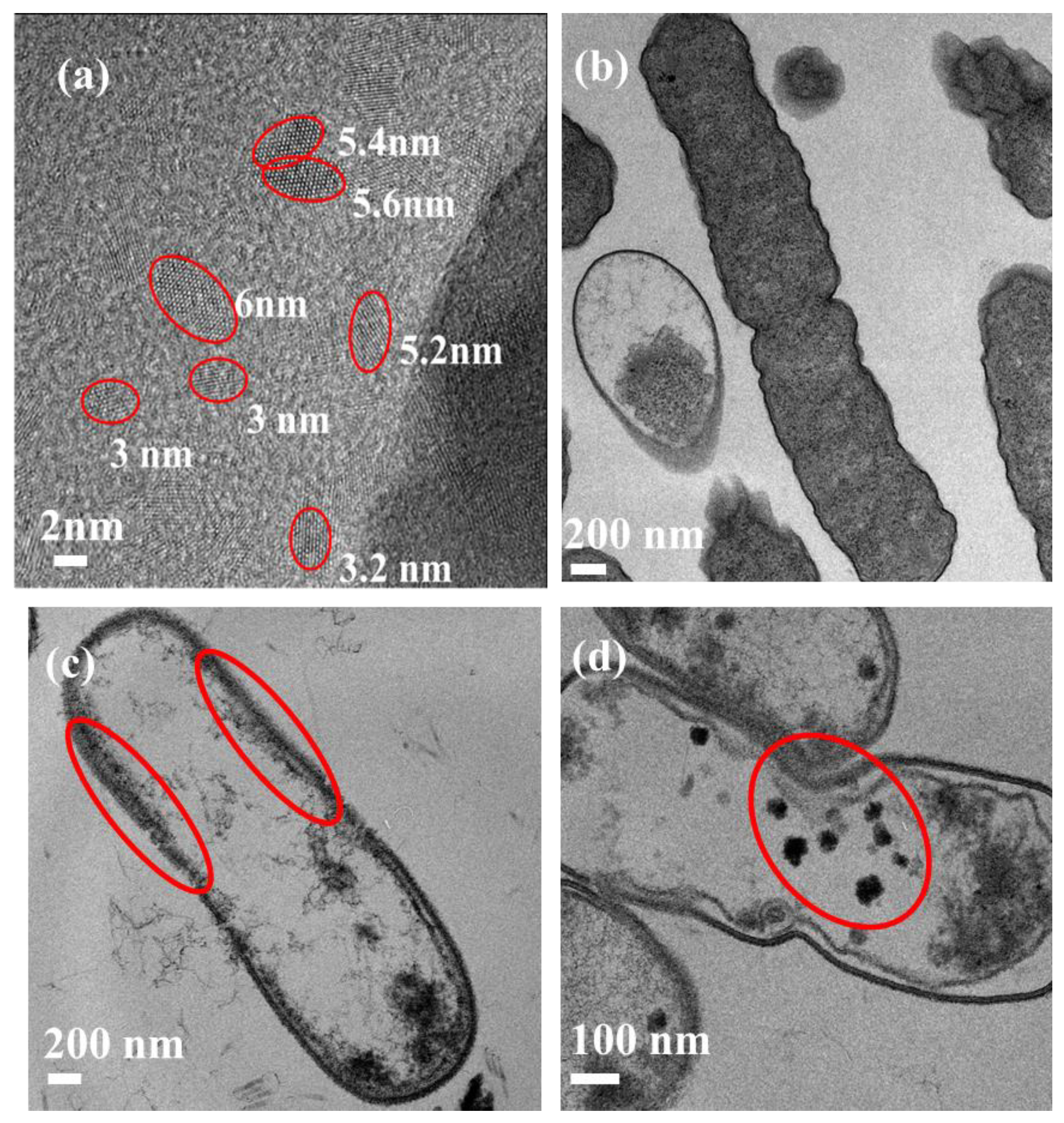
3.1. Investigating the Interaction between CDs and E. coli Bacteria
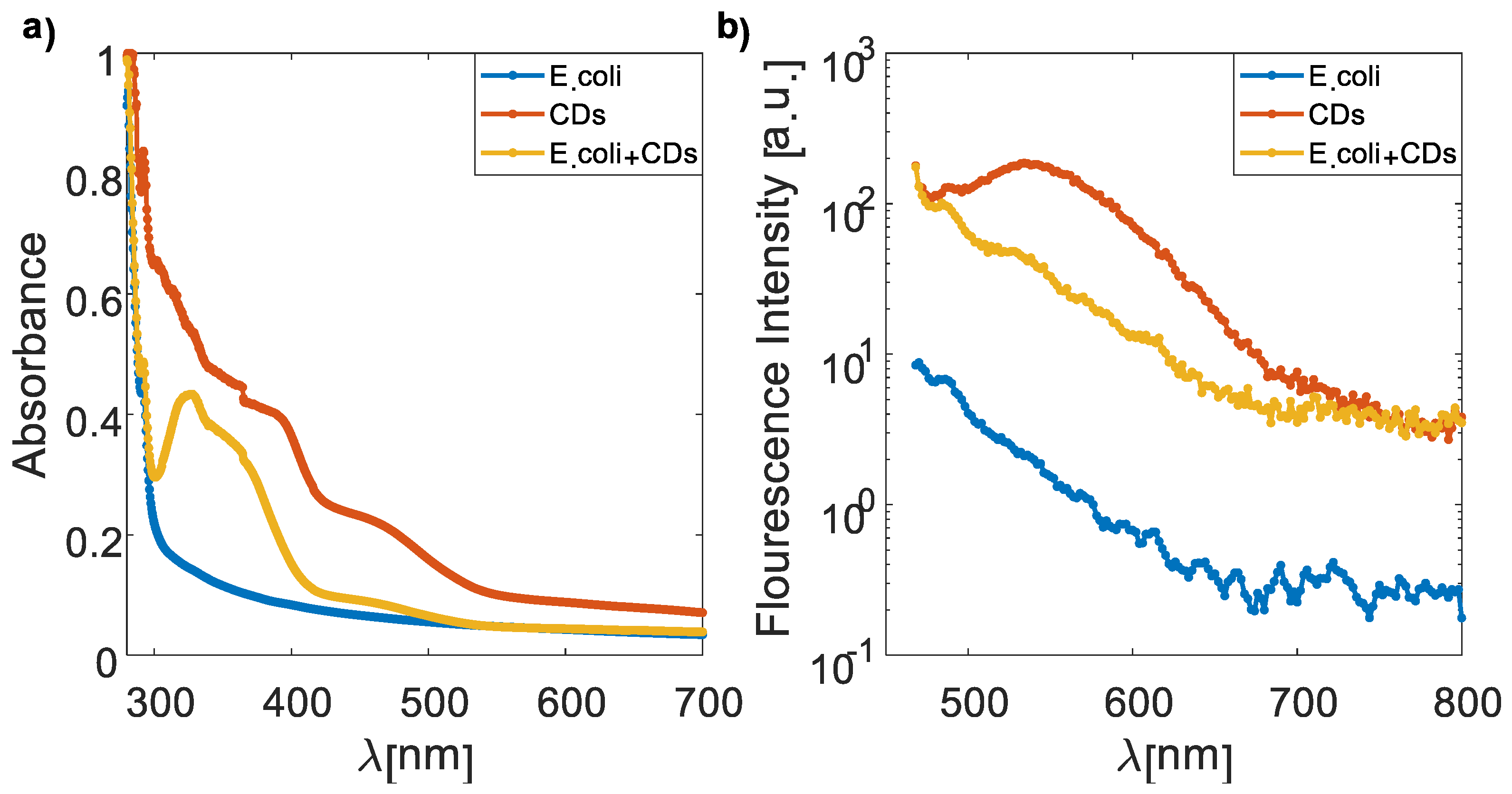
3.2. Absorption and Emission Spectra of CDs, E. coli, and the CD–E. coli Complex
3.3. Investigation of the Polarity and the pH Responsiveness of CDs
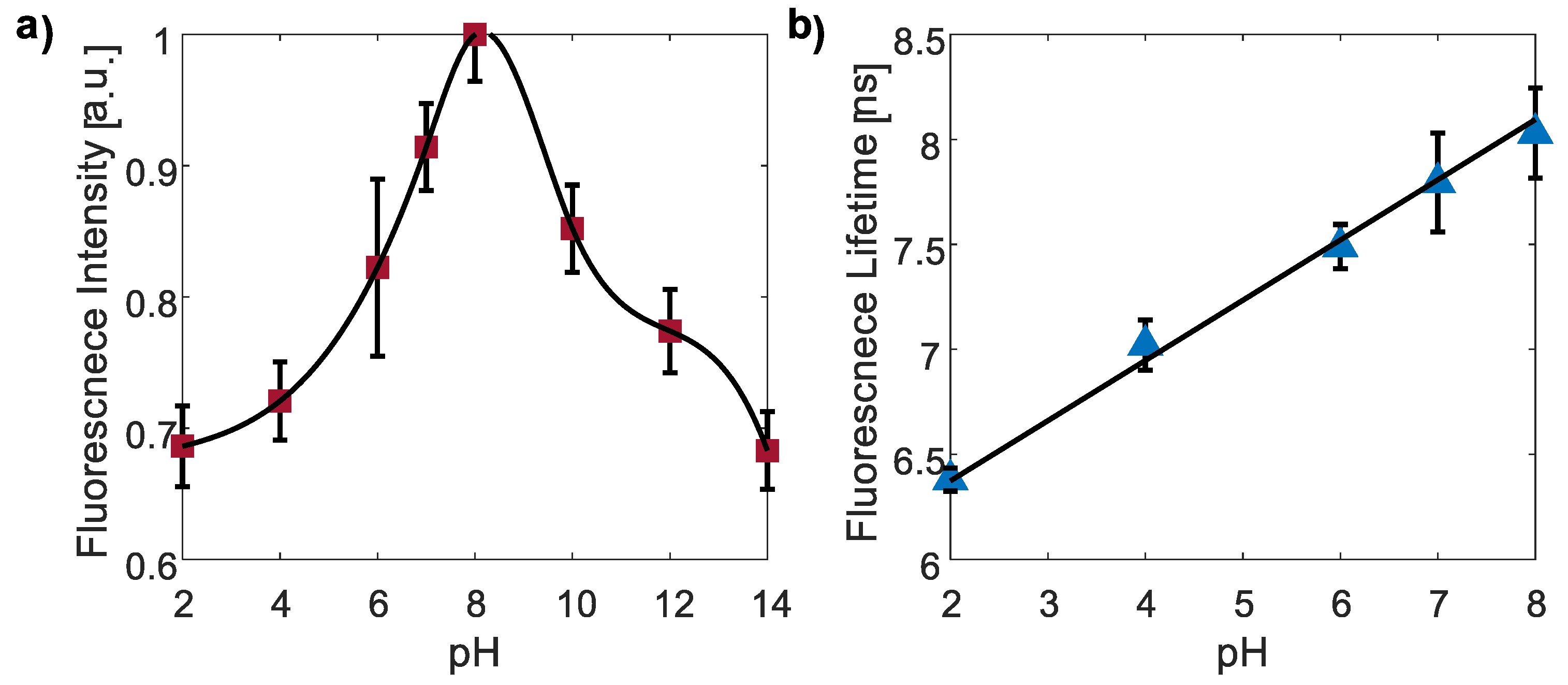
3.3.1. pH Responsivity of CDs
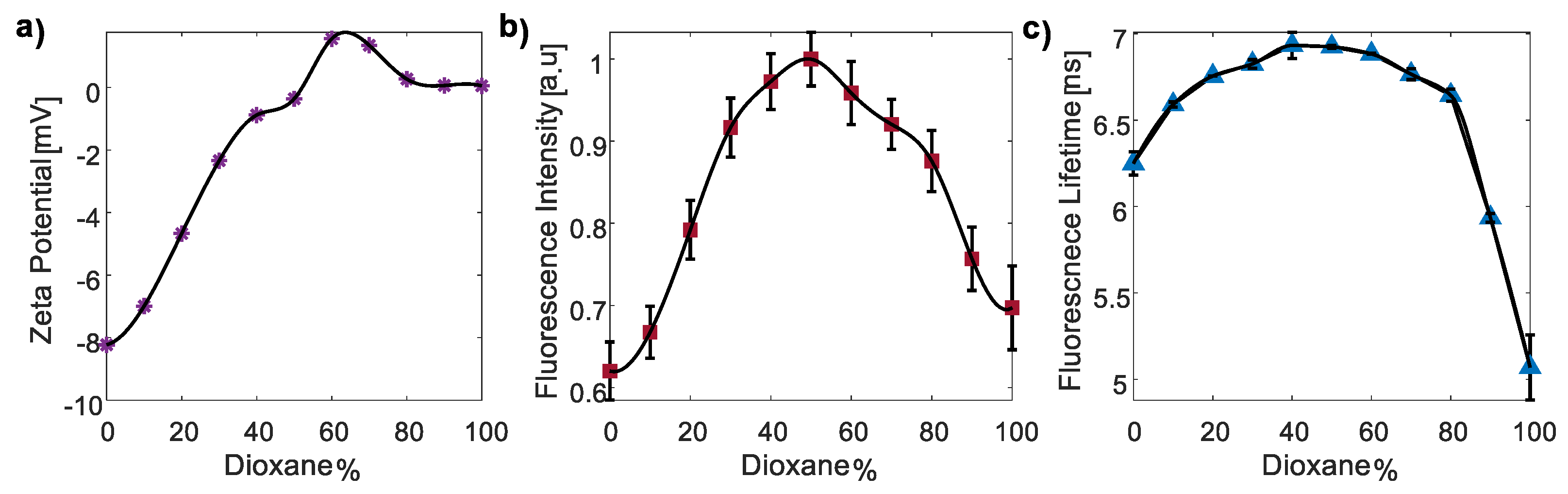
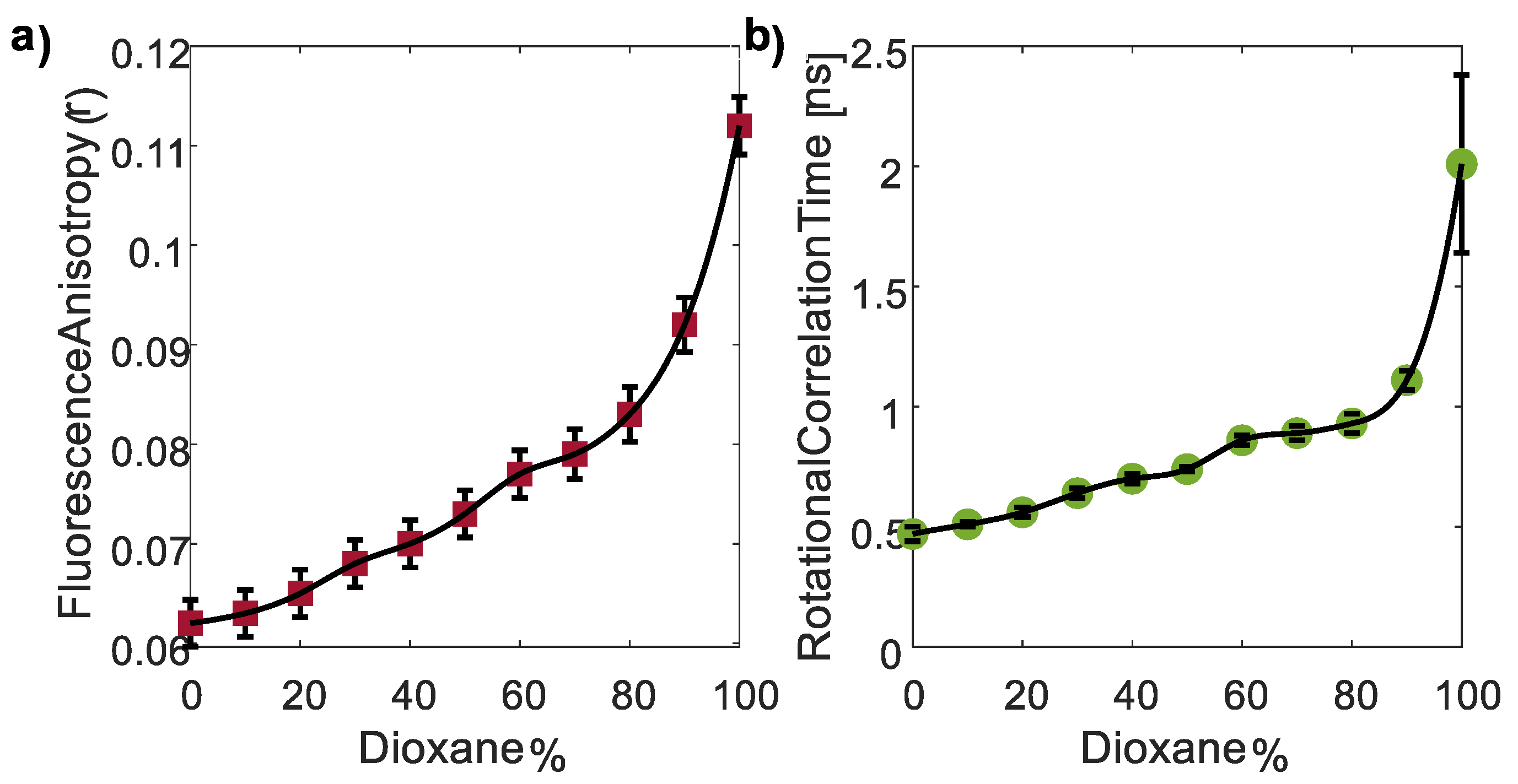
3.3.2. The Polarity Responsivity of CDs
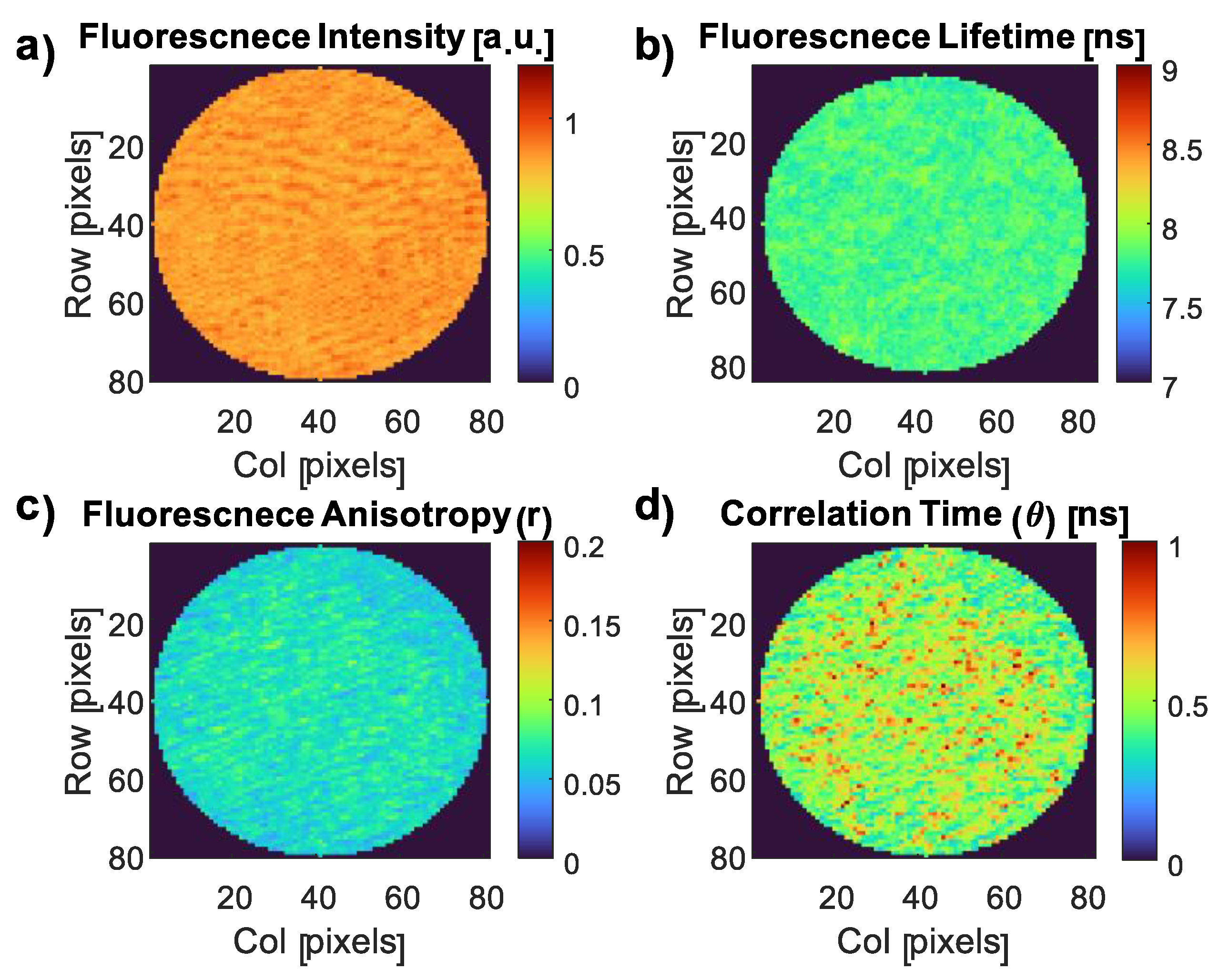
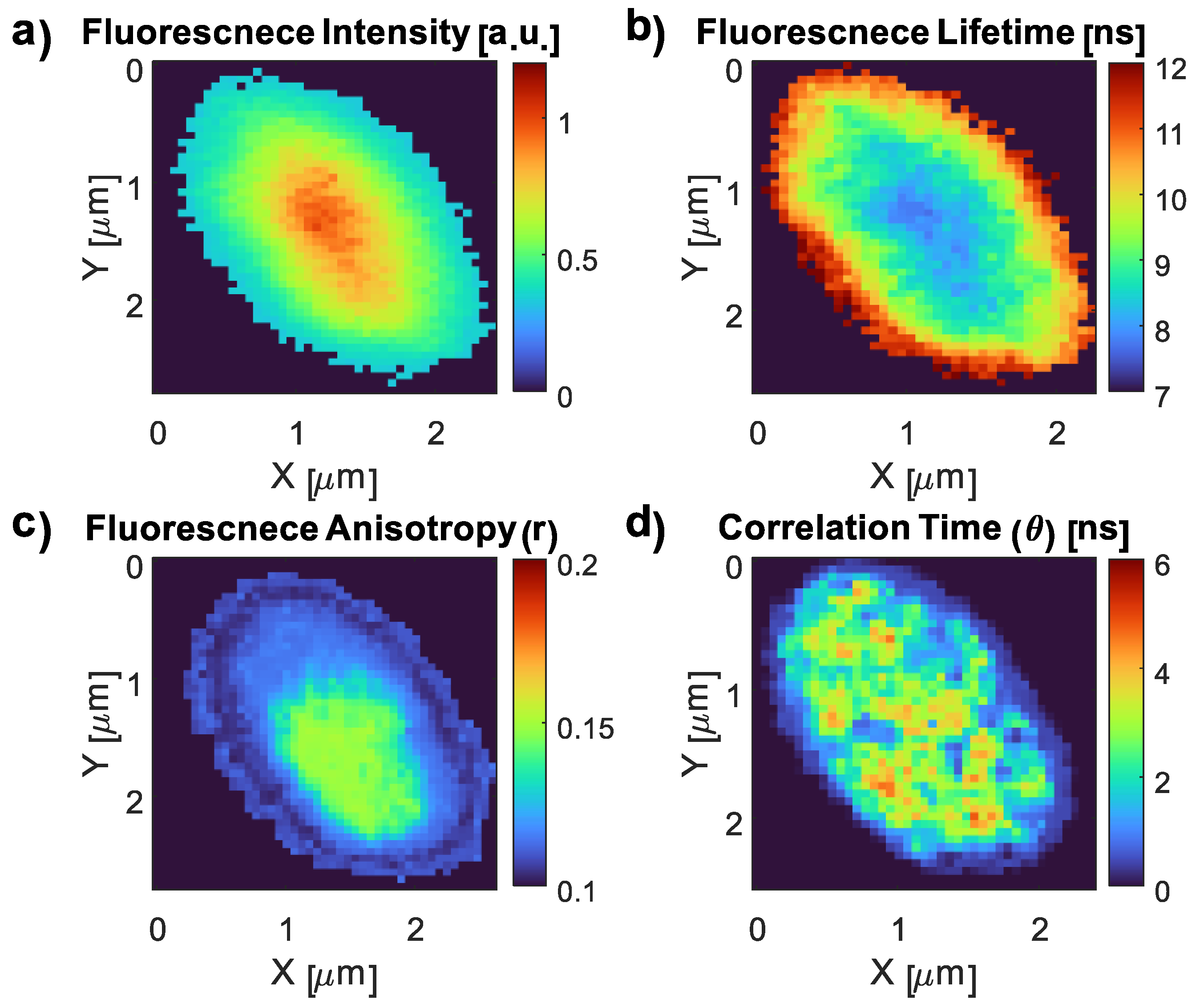
3.4. Fluorescence Imaging of the CDs and the CD–E. coli Complex
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A
Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | TCSPC | FD-FLIM | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| α1 | τ1 | α2 | τ2 | τ3 * | α1 | τ1 | α2 | τ2 | τ3 * | |
| E. coli | 0.55 | 0.43 | 0.27 | 2.24 | 7.85 | N.A | ||||
| CDs | 0.33 | 0.42 | 0.11 | 1.91 | 7.39 | 0.16 | 0.51 ± 0.15 | 0.23 | 1.85 ± 0.16 | 7.80 ± 0.29 |
| CD–E. coli | 0.30 | 0.51 | 0.11 | 2.37 | 7.25 | 0.40 | 0.55 ± 0.24 | 0.08 | 2.38 ± 0.16 | 7.70 ± 0.18 |
| CD | FD TR-FAIM | ||||||
|---|---|---|---|---|---|---|---|
| Exponential Fit | r1 | θ1 [ns] | r2 | θ2 [ns] | r3 | θ3 [ns] | χ2 |
| Single | 0.18 ± 0.01 | 1.83 ± 0.01 | / | / | / | / | 3.91 ± 0.29 |
| Double | 0.36 ± 0.01 | 0.50 ± 0.11 | 0.04 ± 0.01 | 10.00 ± 0.47 | / | / | 1.37 ± 0.12 |
| Triple | 0.28 ± 0.03 | 0.50 ± 0.01 | 0.09 ± 0.01 | 0.50 ± 0.10 | 0.03 ± 0.01 | 10.00 ± 0.10 | 2.20 ± 0.24 |
References
- Han, J.; Burgess, K. Fluorescent indicators for intracellular pH. Chem. Rev. 2010, 110, 2709–2728. [Google Scholar] [CrossRef]
- Xiao, H.; Li, P.; Tang, B. Recent progresses in fluorescent probes for detection of polarity. Coord. Chem. Rev. 2021, 427, 213582. [Google Scholar] [CrossRef]
- Wang, J.; Qiu, J. A review of carbon dots in biological applications. J. Mater. Sci. 2016, 51, 4728–4738. [Google Scholar] [CrossRef]
- Wang, Y.; Anilkumar, P.; Cao, L.; Liu, J.-H.; Luo, P.G.; Tackett, K.N.; Sahu, S.; Wang, P.; Wang, X.; Sun, Y.-P. Carbon dots of different composition and surface functionalization: Cytotoxicity issues relevant to fluorescence cell imaging. Exp. Biol. Med. 2011, 236, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Molaei, M.J. A review on nanostructured carbon quantum dots and their applications in biotechnology, sensors, and chemiluminescence. Talanta 2019, 196, 456–478. [Google Scholar] [CrossRef]
- Ding, H.; Li, X.-H.; Chen, X.-B.; Wei, J.-S.; Li, X.-B.; Xiong, H.-M. Surface states of carbon dots and their influences on luminescence. J. Appl. Phys. 2020, 127, 231101. [Google Scholar] [CrossRef]
- Ai, L.; Yang, Y.; Wang, B.; Chang, J.; Tang, Z.; Yang, B.; Lu, S. Insights into photoluminescence mechanisms of carbon dots: Advances and perspectives. Sci. Bull. 2021, 66, 839–856. [Google Scholar] [CrossRef]
- Zheng, H.; Wang, Q.; Long, Y.; Zhang, H.; Huang, X.; Zhu, R. Enhancing the luminescence of carbon dots with a reduction pathway. Chem. Commun. 2011, 47, 10650–10652. [Google Scholar] [CrossRef]
- Fang, M.; Xia, S.; Bi, J.; Wigstrom, T.P.; Valenzano, L.; Wang, J.; Mazi, W.; Tanasova, M.; Luo, F.-T.; Liu, H. A cyanine-based fluorescent cassette with aggregation-induced emission for sensitive detection of pH changes in live cells. Chem. Commun. 2018, 54, 1133–1136. [Google Scholar] [CrossRef]
- Li, S.; Song, X.; Hu, Z.; Feng, G. A carbon dots-based ratiometric fluorescence probe for monitoring intracellular pH and bioimaging. J. Photochem. Photobiol. A Chem. 2021, 409, 113129. [Google Scholar] [CrossRef]
- Susumu, K.; Field, L.D.; Oh, E.; Hunt, M.; Delehanty, J.B.; Palomo, V.; Dawson, P.E.; Huston, A.L.; Medintz, I.L. Purple-, blue-, and green-emitting multishell alloyed quantum dots: Synthesis, characterization, and application for ratiometric extracellular pH sensing. Chem. Mater. 2017, 29, 7330–7344. [Google Scholar] [CrossRef]
- Shi, B.; Su, Y.; Zhang, L.; Liu, R.; Huang, M.; Zhao, S. Nitrogen-rich functional groups carbon nanoparticles based fluorescent pH sensor with broad-range responding for environmental and live cells applications. Biosens. Bioelectron. 2016, 82, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Barati, A.; Shamsipur, M.; Abdollahi, H. Carbon dots with strong excitation-dependent fluorescence changes towards pH. Application as nanosensors for a broad range of pH. Anal. Chim. Acta 2016, 931, 25–33. [Google Scholar] [CrossRef]
- Qi, P.; Zhang, D.; Zeng, Y.; Wan, Y. Biosynthesis of CdS nanoparticles: A fluorescent sensor for sulfate-reducing bacteria detection. Talanta 2016, 147, 142–146. [Google Scholar] [CrossRef] [PubMed]
- McGoverin, C.; Steed, C.; Esan, A.; Robertson, J.; Swift, S.; Vanholsbeeck, F. Optical methods for bacterial detection and characterization. APL Photonics 2021, 6, 080903. [Google Scholar] [CrossRef]
- Váradi, L.; Luo, J.L.; Hibbs, D.E.; Perry, J.D.; Anderson, R.J.; Orenga, S.; Groundwater, P.W. Methods for the detection and identification of pathogenic bacteria: Past, present, and future. Chem. Soc. Rev. 2017, 46, 4818–4832. [Google Scholar] [CrossRef]
- Wang, Y.; Salazar, J.K. Culture-independent rapid detection methods for bacterial pathogens and toxins in food matrices. Compr. Rev. Food Sci. Food Saf. 2016, 15, 183–205. [Google Scholar] [CrossRef]
- Ma, L.-N.; Zhang, J.; Chen, H.-T.; Zhou, J.-H.; Ding, Y.-Z.; Liu, Y.-S. An overview on ELISA techniques for FMD. Virol. J. 2011, 8, 419. [Google Scholar] [CrossRef] [Green Version]
- Mullis, K.; Faloona, F.; Scharf, S.; Saiki, R.; Horn, G.; Erlich, H. Specific enzymatic amplification of DNA in vitro: The polymerase chain reaction. Biotechnol. Ser. 1992, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.; Chin, L.K.; Ser, W.; Ayi, T.; Yap, P.; Bourouina, T.; Leprince-Wang, Y. Real-time measurement of single bacterium’s refractive index using optofluidic immersion refractometry. Procedia Eng. 2014, 87, 356–359. [Google Scholar] [CrossRef]
- Bonten, M.; Johnson, J.R.; van den Biggelaar, A.H.; Georgalis, L.; Geurtsen, J.; de Palacios, P.I.; Gravenstein, S.; Verstraeten, T.; Hermans, P.; Poolman, J.T. Epidemiology of Escherichia coli bacteremia: A systematic literature review. Clin. Infect. Dis. 2021, 72, 1211–1219. [Google Scholar] [CrossRef] [PubMed]
- Nash, J.H.; Villegas, A.; Kropinski, A.M.; Aguilar-Valenzuela, R.; Konczy, P.; Mascarenhas, M.; Ziebell, K.; Torres, A.G.; Karmali, M.A.; Coombes, B.K. Genome sequence of adherent-invasive Escherichia coli and comparative genomic analysis with other E. coli pathotypes. BMC Genom. 2010, 11, 667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basavaraju, M.; Gunashree, B. Escherichia coli: An Overview of Main Characteristics. In Escherichia coli; Intech Open: London, UK, 2022. [Google Scholar]
- Hu, X.; Li, Y.; Xu, Y.; Gan, Z.; Zou, X.; Shi, J.; Huang, X.; Li, Z.; Li, Y. Green one-step synthesis of carbon quantum dots from orange peel for fluorescent detection of Escherichia coli in milk. Food Chem. 2021, 339, 127775. [Google Scholar] [CrossRef]
- Zuo, P.; Lu, X.; Sun, Z.; Guo, Y.; He, H. A review on syntheses, properties, characterization and bioanalytical applications of fluorescent carbon dots. Microchim. Acta 2016, 183, 519–542. [Google Scholar] [CrossRef]
- Yoon, S.A.; Park, S.Y.; Cha, Y.; Gopala, L.; Lee, M.H. Strategies of detecting bacteria using fluorescence-based dyes. Front. Chem. 2021, 9, 743923. [Google Scholar] [CrossRef]
- Berezin, M.Y.; Achilefu, S. Fluorescence lifetime measurements and biological imaging. Chem. Rev. 2010, 110, 2641–2684. [Google Scholar] [CrossRef] [Green Version]
- Clayton, A.H.; Hanley, Q.S.; Arndt-Jovin, D.J.; Subramaniam, V.; Jovin, T.M. Dynamic fluorescence anisotropy imaging microscopy inthe frequency domain (rFLIM). Biophys. J. 2002, 83, 1631–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zukin, R.S. Evidence for a conformational change in the Escherichia coli maltose receptor by excited-state fluorescence lifetime data. Biochemistry 1979, 18, 2139–2145. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, Y.; Zhang, L.; Zhang, M.; Tian, C. Conformational change of E. coli sulfurtransferase YgaP upon SCN− in intact native membrane revealed by fluorescence lifetime and anisotropy. Chin. Chem. Lett. 2018, 29, 1513–1516. [Google Scholar] [CrossRef]
- Jameson, D.M.; Ross, J.A. Fluorescence polarization/anisotropy in diagnostics and imaging. Chem. Rev. 2010, 110, 2685–2708. [Google Scholar] [CrossRef] [Green Version]
- Kühn, O.; Sundström, V.; Pullerits, T. Fluorescence depolarization dynamics in the B850 complex of purple bacteria. Chem. Phys. 2002, 275, 15–30. [Google Scholar] [CrossRef]
- Lakowicz, J.R. Time-Dependent Anisotropy Decays. In Principles of Fluorescence Spectroscopy, 3rd ed.; Lakowicz, J., Ed.; Springer: New York, NY, USA, 2013; pp. 383–412. [Google Scholar]
- Lakowicz, J.R. Time-Domain Lifetime Measurements. In Principles of Fluorescence Spectroscopy, 3rd ed.; Lakowicz, J., Ed.; Springer: New York, NY, USA, 2013; pp. 98–154. [Google Scholar]
- Lakowicz, J.R. Frequency-Domain Lifetime Measurements. In Principles of Fluorescence Spectroscopy, 3rd ed.; Lakowicz, J., Ed.; Springer: New York, NY, USA, 2013; pp. 158–203. [Google Scholar]
- Vetromile, C.M.; Jameson, D.M. Frequency domain fluorometry: Theory and application. In Fluorescence Spectroscopy and Microscopy; Springer: New York, NY, USA, 2014; pp. 77–95. [Google Scholar]
- Yahav, G.; Gershanov, S.; Salmon-Divon, M.; Ben-Zvi, H.; Mircus, G.; Goldenberg-Cohen, N.; Fixler, D. Pathogen detection using frequency domain fluorescent lifetime measurements. IEEE Trans. Biomed. Eng. 2018, 65, 2731–2741. [Google Scholar] [CrossRef]
- Yahav, G.; Diamandi, H.H.; Preter, E.; Fixler, D. The squared distance approach to frequency domain time-resolved fluorescence analysis. J. Biophotonics 2019, 12, e201800485. [Google Scholar] [CrossRef]
- Yahav, G.; Pawar, S.; Weber, Y.; Atuar, B.; Duadi, H.; Fixler, D. Imaging the rotational mobility of carbon dot-gold nanoparticle conjugates using frequency domain wide-field time-resolved fluorescence anisotropy. J. Biomed. Opt. 2023, 28, 056001. [Google Scholar] [CrossRef] [PubMed]
- Geest, L.v.; Stoop, K. FLIM on a wide field fluorescence microscope. Lett. Pept. Sci. 2003, 10, 501–510. [Google Scholar] [CrossRef]
- Lakowicz, J.R.; Cherek, H.; Kuśba, J.; Gryczynski, I.; Johnson, M.L. Review of fluorescence anisotropy decay analysis by frequency-domain fluorescence spectroscopy. J. Fluoresc. 1993, 3, 103–116. [Google Scholar] [CrossRef]
- Fixler, D.; Namer, Y.; Yishay, Y.; Deutsch, M. Influence of fluorescence anisotropy on fluorescence intensity and lifetime measurement: Theory, simulations and experiments. IEEE Trans. Biomed. Eng. 2006, 53, 1141–1152. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, L.; McAdams, H.H.; Losick, R. Generating and exploiting polarity in bacteria. Science 2002, 298, 1942–1946. [Google Scholar] [CrossRef]
- Bowman, G.R.; Lyuksyutova, A.I.; Shapiro, L. Bacterial polarity. Curr. Opin. Cell Biol. 2011, 23, 71–77. [Google Scholar] [CrossRef]
- Bhattacharjee, S. DLS and zeta potential—What they are and what they are not? J. Control. Release 2016, 235, 337–351. [Google Scholar] [CrossRef]
- Ran, H.-H.; Cheng, X.; Bao, Y.-W.; Hua, X.-W.; Gao, G.; Zhang, X.; Jiang, Y.-W.; Zhu, Y.-X.; Wu, F.-G. Multifunctional quaternized carbon dots with enhanced biofilm penetration and eradication efficiencies. J. Mater. Chem. B 2019, 7, 5104–5114. [Google Scholar] [CrossRef] [PubMed]
- Zühlke, M.; Meiling, T.T.; Roder, P.; Riebe, D.; Beitz, T.; Bald, I.; Löhmannsröben, H.G.; Janßen, T.; Erhard, M.; Repp, A. Photodynamic Inactivation of E. coli Bacteria via Carbon Nanodots. ACS Omega 2021, 6, 23742–23749. [Google Scholar] [CrossRef] [PubMed]
- Das, P.; Bose, M.; Ganguly, S.; Mondal, S.; Das, A.K.; Banerjee, S.; Das, N.C. Green approach to photoluminescent carbon dots for imaging of gram-negative bacteria Escherichia coli. Nanotechnology 2017, 28, 195501. [Google Scholar] [CrossRef] [PubMed]
- Lakowicz, J.R.; Lakowicz, J.R. Effects of solvents on fluorescence emission spectra. In Principles of Fluorescence Spectroscopy; Springer: Berlin/Heidelberg, Germany, 1983; pp. 187–215. [Google Scholar]
- Shamsipur, M.; Barati, A.; Taherpour, A.A.; Jamshidi, M. Resolving the multiple emission centers in carbon dots: From fluorophore molecular states to aromatic domain states and carbon-core states. J. Phys. Chem. Lett. 2018, 9, 4189–4198. [Google Scholar] [CrossRef] [PubMed]
- Parhad, N.; Rao, N. Effect of pH on survival of Escherichia coli. J. Water Pollut. Control. Fed. 1974, 46, 980–986. [Google Scholar]
- Wilks, J.C.; Slonczewski, J.L. pH of the cytoplasm and periplasm of Escherichia coli: Rapid measurement by green fluorescent protein fluorimetry. J. Bacteriol. 2007, 189, 5601–5607. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.L.; Gao, M.X.; Wang, T.T.; Wan, X.Y.; Zheng, L.L.; Huang, C.Z. A general quantitative pH sensor developed with dicyandiamide N-doped high quantum yield graphene quantum dots. Nanoscale 2014, 6, 3868–3874. [Google Scholar] [CrossRef]
- Huang, M.; Liang, X.; Zhang, Z.; Wang, J.; Fei, Y.; Ma, J.; Qu, S.; Mi, L. Carbon dots for intracellular pH sensing with fluorescence lifetime imaging microscopy. Nanomaterials 2020, 10, 604. [Google Scholar] [CrossRef] [Green Version]
- Ehtesabi, H.; Hallaji, Z.; Najafi Nobar, S.; Bagheri, Z. Carbon dots with pH-responsive fluorescence: A review on synthesis and cell biological applications. Microchim. Acta 2020, 187, 150. [Google Scholar] [CrossRef]
- Huo, X.; Shen, H.; Xu, Y.; Shao, J.; Liu, R.; Zhang, Z. Fluorescence properties of carbon dots synthesized by different solvents for pH detector. Opt. Mater. 2022, 123, 111889. [Google Scholar] [CrossRef]
- Kozioł, J.; Knobloch, E. The solvent effect on the fluorescence and light absorption of riboflavin and lumiflavin. Biochim. Biophys. Acta BBA-Biophys. Incl. Photosynth. 1965, 102, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Guo, X.; Bai, X.; Wang, X.; Ding, Q.; Zhang, W.; Zhang, W.; Tang, B. Golgi apparatus polarity indicates depression-like behaviors of mice using in vivo fluorescence imaging. Anal. Chem. 2019, 91, 3382–3388. [Google Scholar] [CrossRef] [PubMed]
- Radchanka, A.; Hrybouskaya, V.; Iodchik, A.; Achtstein, A.W.; Artemyev, M. Zeta Potential-Based Control of CdSe/ZnS Quantum Dot Photoluminescence. J. Phys. Chem. Lett. 2022, 13, 4912–4917. [Google Scholar] [CrossRef]
- Siegel, J.; Suhling, K.; Lévêque-Fort, S.; Webb, S.E.; Davis, D.M.; Phillips, D.; Sabharwal, Y.; French, P.M. Wide-field time-resolved fluorescence anisotropy imaging (TR-FAIM): Imaging the rotational mobility of a fluorophore. Rev. Sci. Instrum. 2003, 74, 182–192. [Google Scholar] [CrossRef] [Green Version]
- Hirayama, F.; Lawson, C.W.; Lipsky, S. Fluorescence of p-dioxane. J. Phys. Chem. 1970, 74, 2411–2413. [Google Scholar] [CrossRef]
- Fletcher, A.N. Fluorescence emission band shift with wavelength of excitation. J. Phys. Chem. 1968, 72, 2742–2749. [Google Scholar] [CrossRef]
- Belfield, K.D.; Bondar, M.V.; Kachkovsky, O.D.; Przhonska, O.V.; Yao, S. Solvent effect on the steady-state fluorescence anisotropy of two-photon absorbing fluorene derivatives. J. Lumin. 2007, 126, 14–20. [Google Scholar] [CrossRef]
- Kartzmark, E.M. Conductances, Densities, and Viscosities of Solutions of Sodium Nitrate in Water and in Dioxane–Water, at 25 C. Can. J. Chem. 1972, 50, 2845–2850. [Google Scholar] [CrossRef]
- Renggli, S.; Keck, W.; Jenal, U.; Ritz, D. Role of autofluorescence in flow cytometric analysis of Escherichia coli treated with bactericidal antibiotics. J. Bacteriol. 2013, 195, 4067–4073. [Google Scholar] [CrossRef] [Green Version]
- Eigenfeld, M.; Kerpes, R.; Whitehead, I.; Becker, T. Autofluorescence prediction model for fluorescence unmixing and age determination. Biotechnol. J. 2022, 17, 2200091. [Google Scholar] [CrossRef]
- Thanassi, D.G.; Suh, G.; Nikaido, H. Role of outer membrane barrier in efflux-mediated tetracycline resistance of Escherichia coli. J. Bacteriol. 1995, 177, 998–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sträuber, H.; Müller, S. Viability states of bacteria—Specific mechanisms of selected probes. Cytom. Part A 2010, 77, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Benarroch, J.M.; Asally, M. The microbiologist’s guide to membrane potential dynamics. Trends Microbiol. 2020, 28, 304–314. [Google Scholar] [CrossRef] [Green Version]
- Mohapatra, S.; Ranjan, S.; Dasgupta, N.; Kumar, R.K.; Thomas, S. Nanocarriers for Drug Delivery: Nanoscience and Nanotechnology in Drug Delivery; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Bagatolli, L.A.; Stock, R.P. Lipids, membranes, colloids and cells: A long view. Biochim. Biophys. Acta BBA-Biomembr. 2021, 1863, 183684. [Google Scholar] [CrossRef] [PubMed]
- Lewis, C.L.; Craig, C.C.; Senecal, A.G. Mass and density measurements of live and dead gram-negative and gram-positive bacterial populations. Appl. Environ. Microbiol. 2014, 80, 3622–3631. [Google Scholar] [CrossRef] [Green Version]
- Scherrer, R.; Berlin, E.; Gerhardt. Density, porosity, and structure of dried cell walls isolated from Bacillus megaterium and Saccharomyces cerevisiae. J. Bacteriol. 1977, 129, 1162–1164. [Google Scholar] [CrossRef] [Green Version]
- Meselson, M.; Stahl, F.W. The replication of DNA in Escherichia coli. Proc. Natl. Acad. Sci. USA 1958, 44, 671–682. [Google Scholar] [CrossRef]
- Rowlett, V.W.; Margolin, W. The bacterial Min system. Curr. Biol. 2013, 23, R553–R556. [Google Scholar] [CrossRef] [Green Version]
- Yahav, G.; Hirshberg, A.; Salomon, O.; Amariglio, N.; Trakhtenbrot, L.; Fixler, D. Fluorescence lifetime imaging of DAPI-stained nuclei as a novel diagnostic tool for the detection and classification of B-cell chronic lymphocytic leukemia. Cytom. Part A 2016, 89, 644–652. [Google Scholar] [CrossRef]
- Gershanov, S.; Michowiz, S.; Toledano, H.; Yahav, G.; Barinfeld, O.; Hirshberg, A.; Ben-Zvi, H.; Mircus, G.; Salmon-Divon, M.; Fixler, D. Fluorescence lifetime imaging microscopy, a novel diagnostic tool for metastatic cell detection in the cerebrospinal fluid of children with medulloblastoma. Sci. Rep. 2017, 7, 3648. [Google Scholar] [CrossRef] [Green Version]
- Zahavi, T.; Yahav, G.; Shimshon, Y.; Gershanov, S.; Kaduri, L.; Sonnenblick, A.; Fixler, D.; Salmon, A.Y.; Salmon-Divon, M. Utilizing fluorescent life time imaging microscopy technology for identify carriers of BRCA2 mutation. Biochem. Biophys. Res. Commun. 2016, 480, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Nakache, G.; Yahav, G.; Siloni, G.; Barshack, I.; Alon, E.; Wolf, M.; Fixler, D. The use of fluorescence lifetime technology in benign and malignant thyroid tissues. J. Laryngol. Otol. 2019, 133, 696–699. [Google Scholar] [CrossRef] [PubMed]
- Suhling, K.; Hirvonen, L.M.; Levitt, J.A.; Chung, P.-H.; Tregidgo, C.; Le Marois, A.; Rusakov, D.A.; Zheng, K.; Ameer-Beg, S.; Poland, S. Fluorescence lifetime imaging (FLIM): Basic concepts and some recent developments. Med. Photonics 2015, 27, 3–40. [Google Scholar] [CrossRef] [Green Version]
- Suhling, K.; Siegel, J.; Lanigan, P.M.; Lévêque-Fort, S.; Webb, S.E.; Phillips, D.; Davis, D.M.; French, P.M. Time-resolved fluorescence anisotropy imaging applied to live cells. Opt. Lett. 2004, 29, 584–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yahav, G.; Pawar, S.; Lipovsky, A.; Gupta, A.; Gedanken, A.; Duadi, H.; Fixler, D. Probing Polarity and pH Sensitivity of Carbon Dots in Escherichia coli through Time-Resolved Fluorescence Analyses. Nanomaterials 2023, 13, 2068. https://doi.org/10.3390/nano13142068
Yahav G, Pawar S, Lipovsky A, Gupta A, Gedanken A, Duadi H, Fixler D. Probing Polarity and pH Sensitivity of Carbon Dots in Escherichia coli through Time-Resolved Fluorescence Analyses. Nanomaterials. 2023; 13(14):2068. https://doi.org/10.3390/nano13142068
Chicago/Turabian StyleYahav, Gilad, Shweta Pawar, Anat Lipovsky, Akanksha Gupta, Aharon Gedanken, Hamootal Duadi, and Dror Fixler. 2023. "Probing Polarity and pH Sensitivity of Carbon Dots in Escherichia coli through Time-Resolved Fluorescence Analyses" Nanomaterials 13, no. 14: 2068. https://doi.org/10.3390/nano13142068