
Co-Treatment of Caco-2 Cells with Doxorubicin and Gold Nanoparticles Produced from Cyclopia intermedia Extracts or Mangiferin Enhances Drug Effects

 , , , and
, , , and 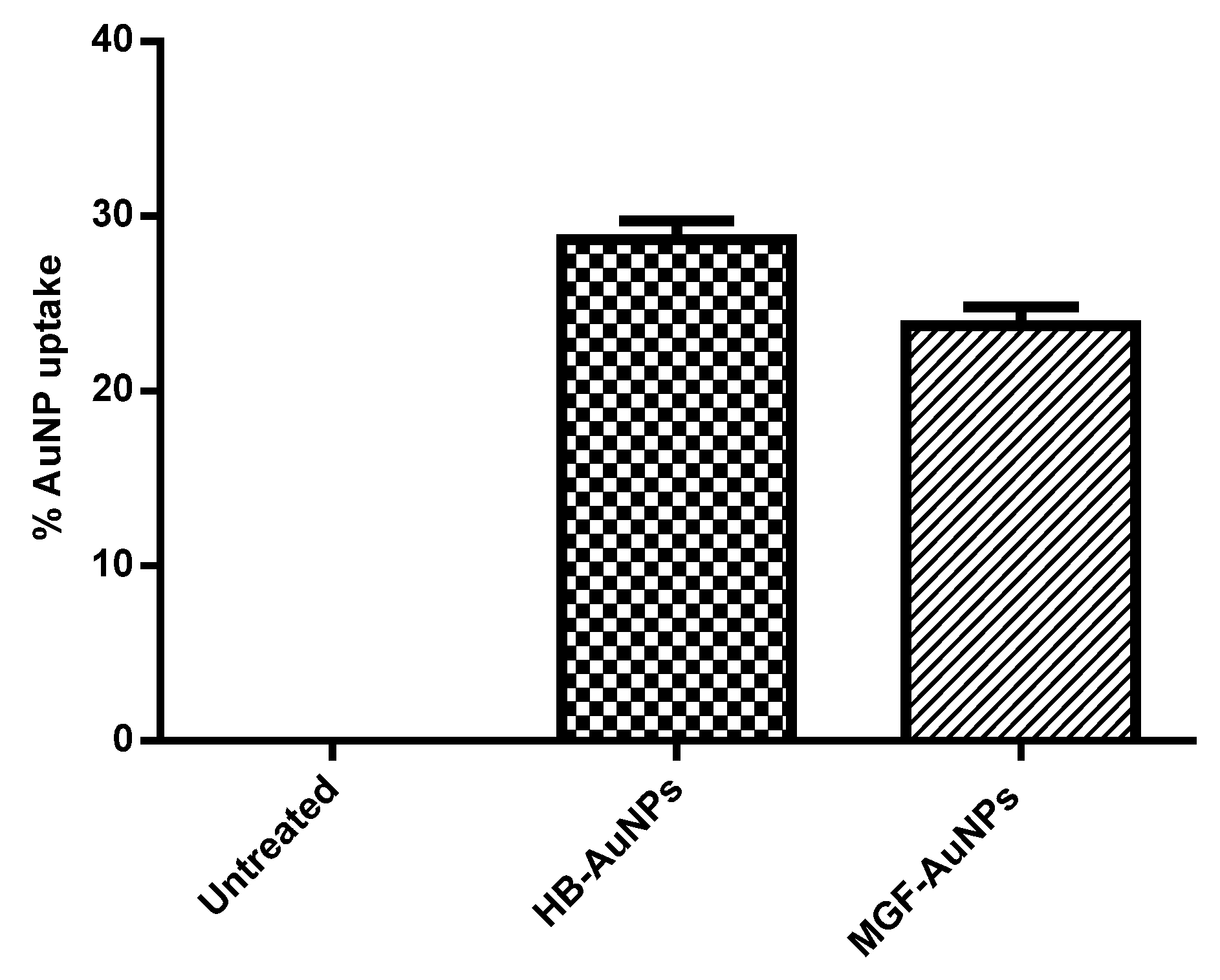{kind=link}
{kind=link}
{kind=link}
{kind=link}
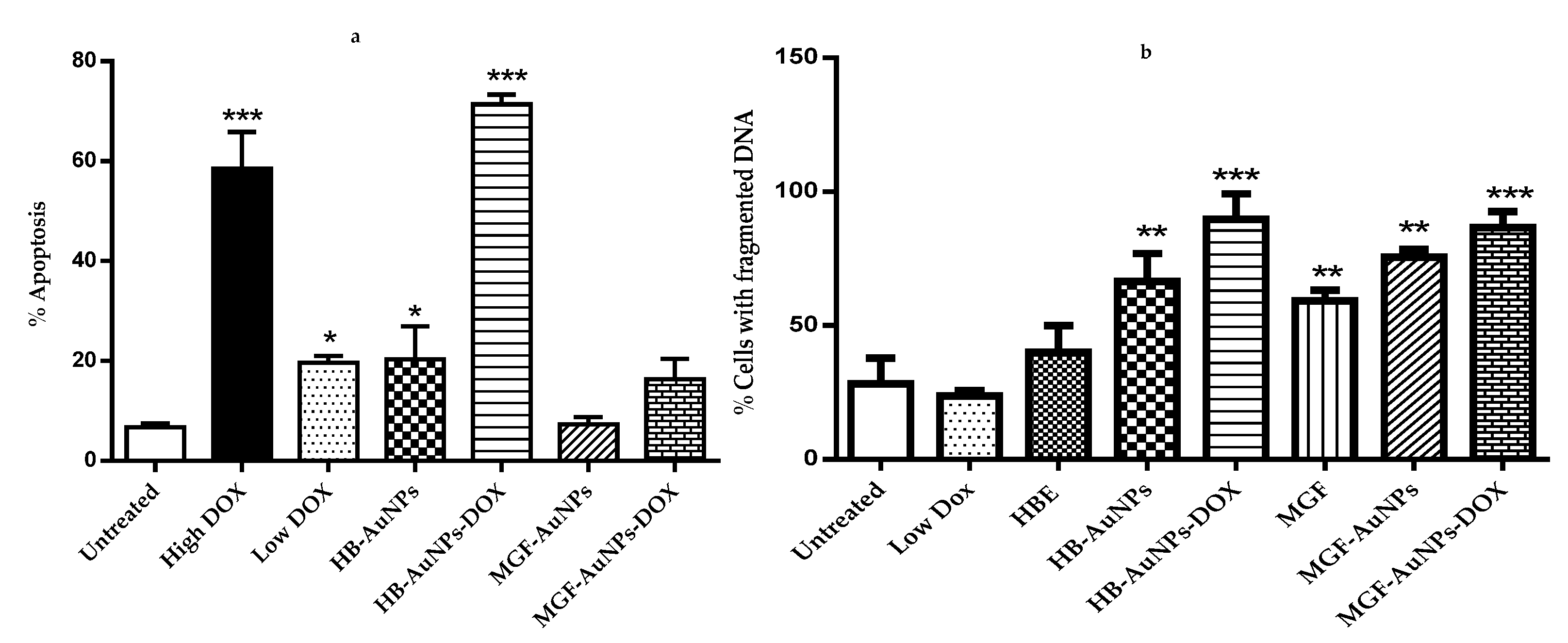{kind=link}
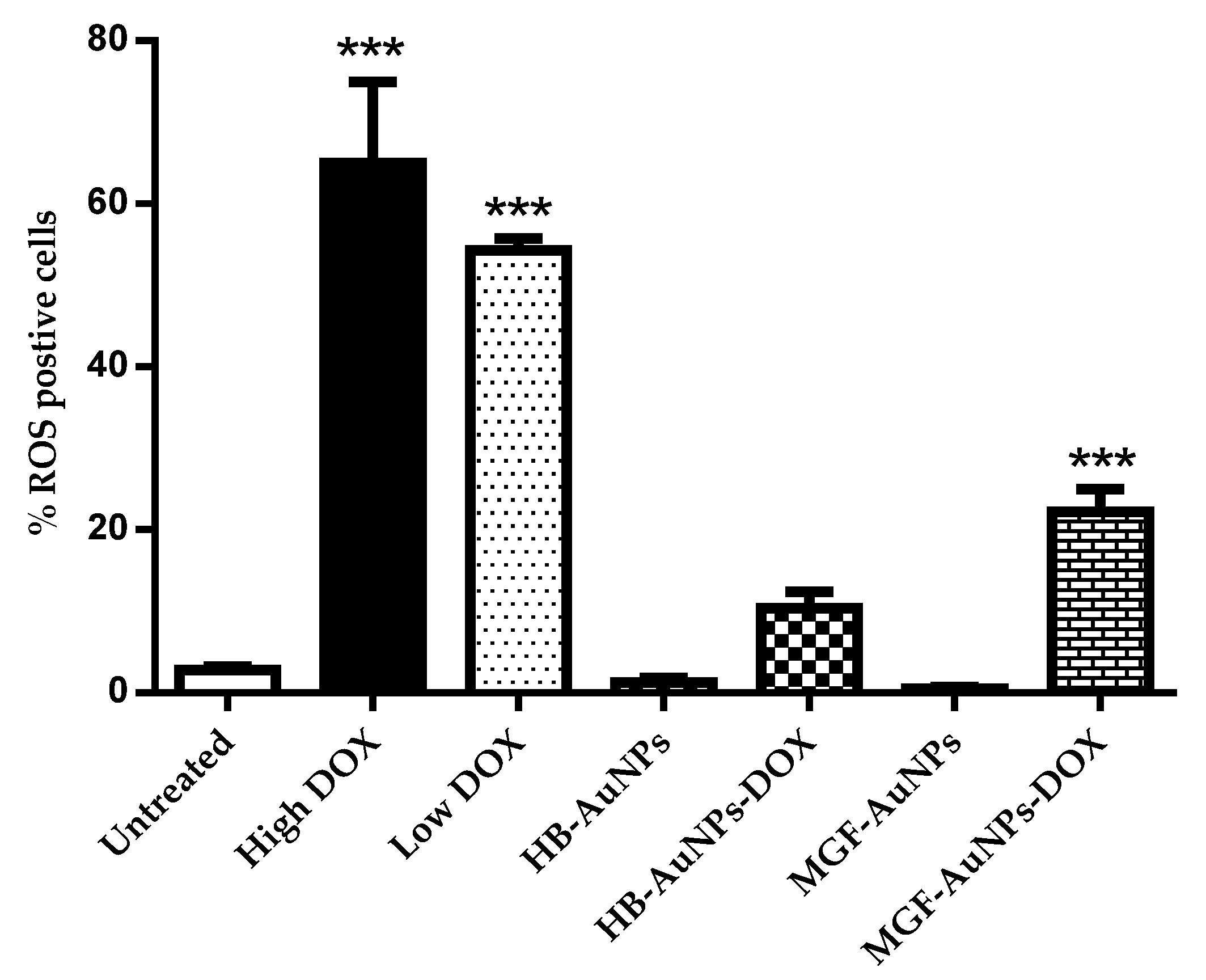{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Synthesis and Characterization of Biogenic MGF-AuNPs and HB-AuNPs
2.2. Biogenic AuNPs Cellular Uptake Study
Analysis of AuNPs Uptake by Inductively Coupled Plasma—Optical Emission Spectrometry (ICP-OES)
2.3. Investigation of the Mechanism of Co-Treatment
2.3.1. Cell Culture and Treatments
2.3.2. Cell Viability Assay
2.3.3. Mitochondrial ToxGlo Assay
2.3.4. Flow Cytometry-Based Bioassays
2.3.4.1. Mitochondrial Depolarization Assay
2.3.4.2. DNA Fragmentation Assay
2.3.4.3. APOPercentage Assay
2.3.4.4. ROS Assay
2.3.5. Colony Formation Assay
2.4. Statistical Analysis
3. Results and Discussion
3.1. Synthesis and Characterization of Biogenic AuNPs
3.2. Cellular Uptake of Biogenic AuNPs in Caco-2 Cells
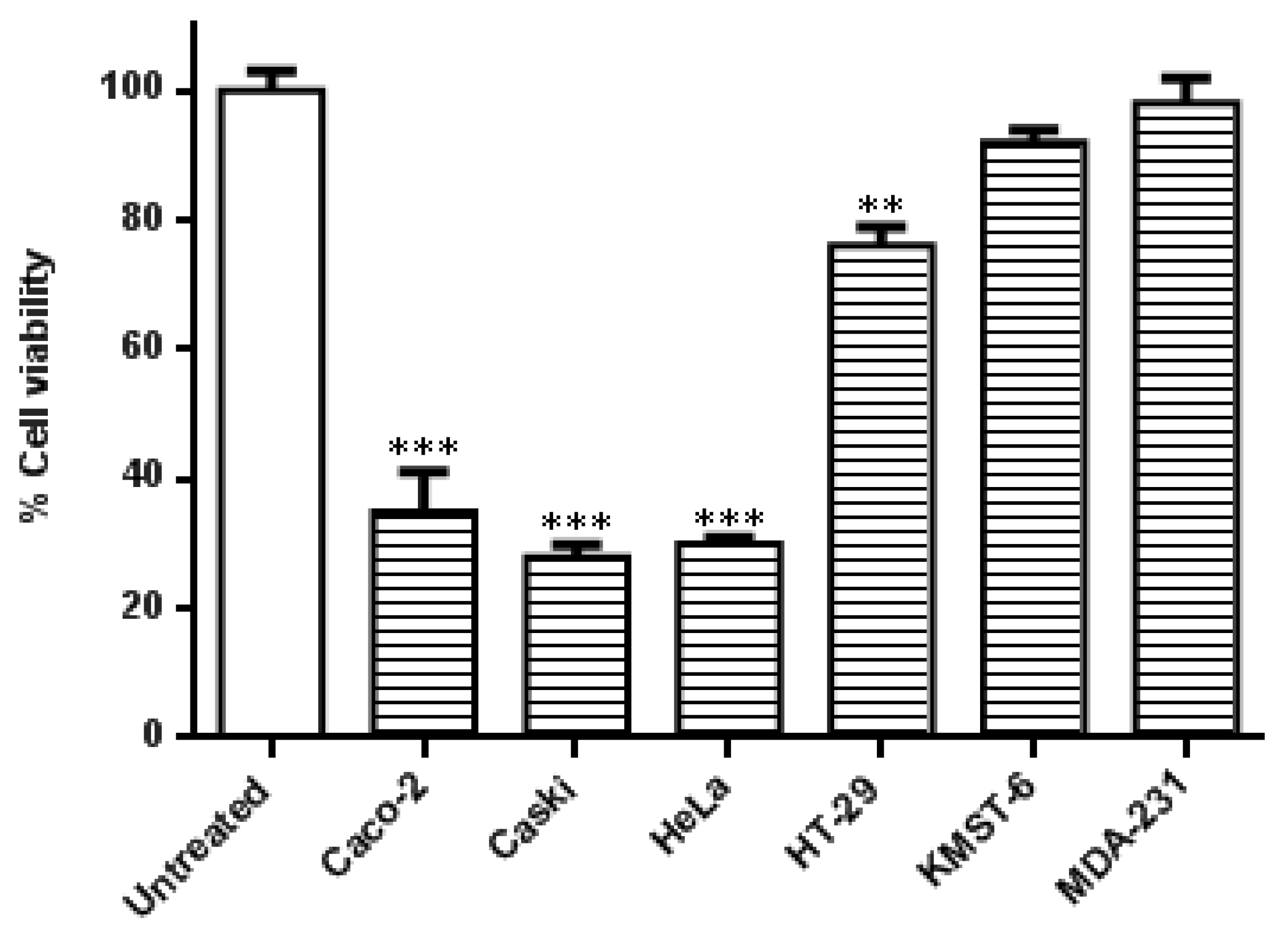
3.3. Co-Treatment of Cells with HB-AuNPs and DOX
3.4. The Effects of Co-Treatment with Biogenic AuNPs and DOX
3.4.1. The Effects of Co-Treatment on Cellular ATP Levels
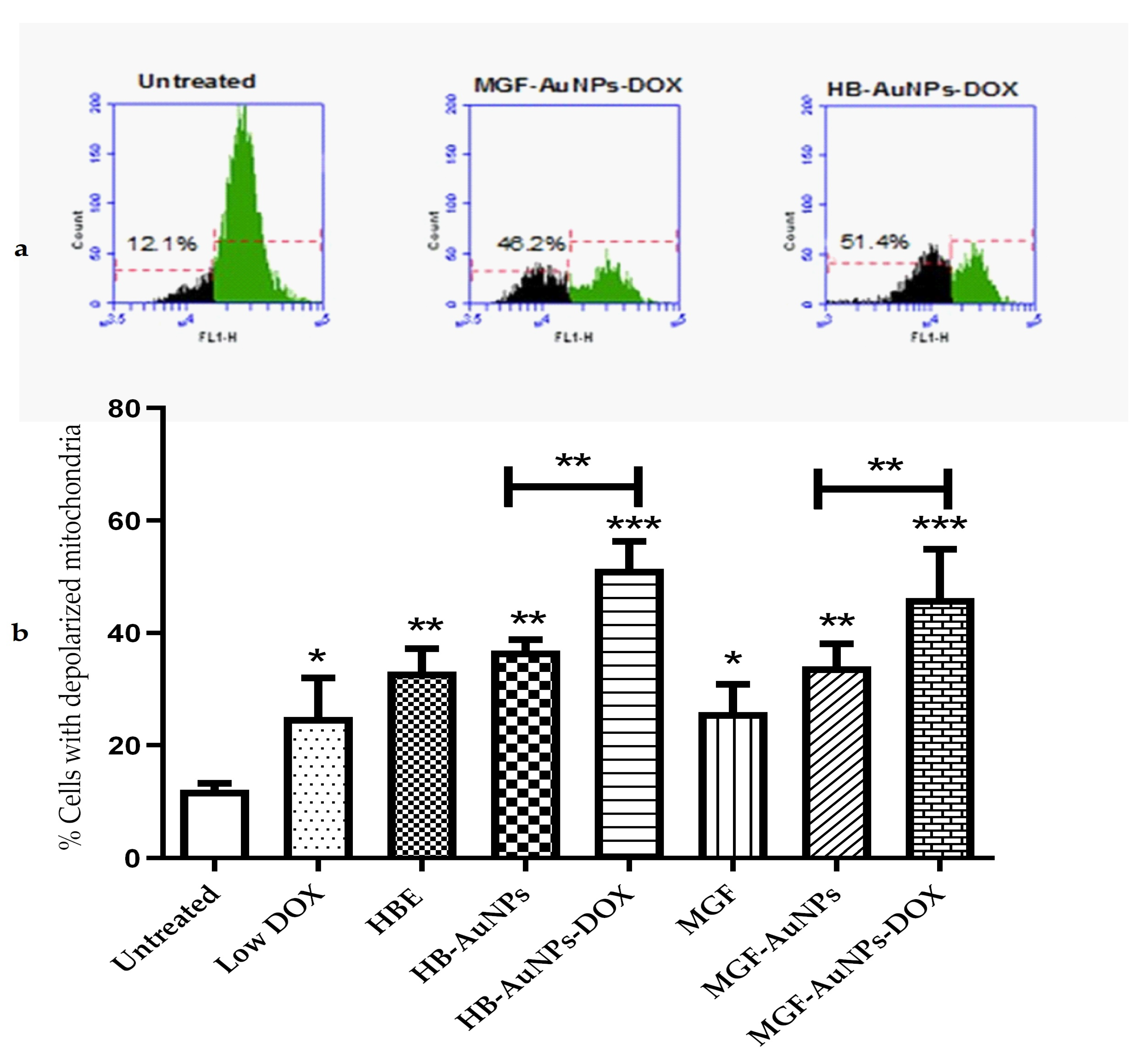
3.4.2. The Effect of Co-Treatment on Mitochondrial Function
3.4.3. The Apoptotic Effects of Co-Treatment with Biogenic AuNPs and DOX
3.4.4. The Effect of the Biogenic AuNP and DOX Co-Treatment on ROS Levels
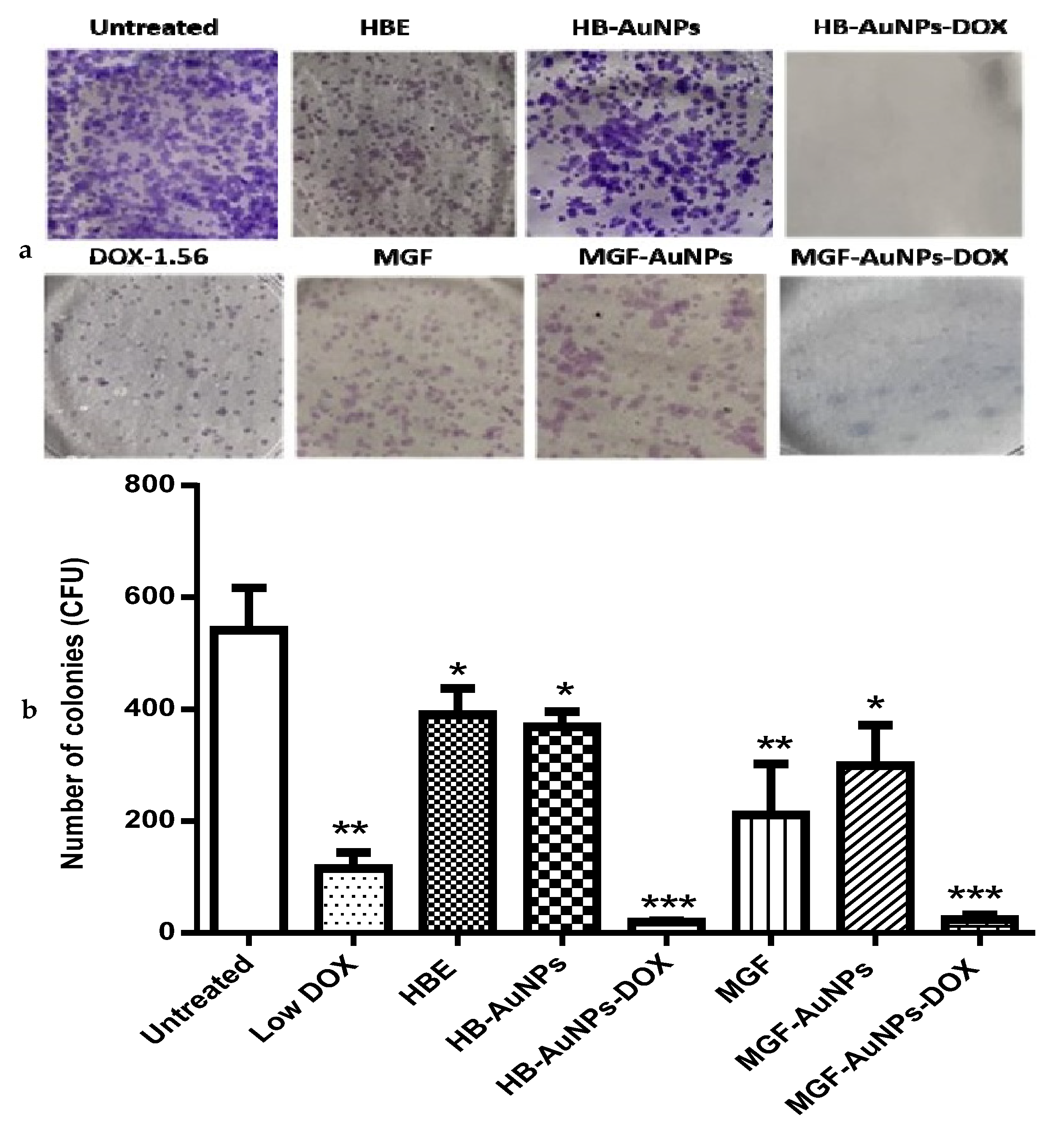
3.5. The Effect of the Biogenic AuNP and DOX Co-Treatment on the Long-Term Survival of Caco-2 Cells
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dehelean, C.A.; Marcovici, I.; Soica, C.; Mioc, M.; Coricovac, D.; Iurciuc, S.; Cretu, O.M.; Pinzaru, I. Plant-Derived Anticancer Compounds as New Perspectives in Drug Discovery and Alternative Therapy. Molecules 2021, 26, 1109. [Google Scholar] [CrossRef] [PubMed]
- Tinoush, B.; Shirdel, I.; Wink, M. Phytochemicals: Potential Lead Molecules for MDR Reversal. Front. Pharmacol. 2020, 11, 832. [Google Scholar] [CrossRef] [PubMed]
- Morozkina, S.N.; Nhung Vu, T.H.; Generalova, Y.E.; Snetkov, P.P.; Uspenskaya, M.V. Mangiferin as New Potential Anti-Cancer Agent and Mangiferin-Integrated Polymer Systems—A Novel Research Direction. Biomolecules 2021, 11, 79. [Google Scholar] [CrossRef] [PubMed]
- Khurana, R.K.; Kaur, R.; Lohan, S.; Singh, K.K.; Singh, B. Mangiferin: A Promising Anticancer Bioactive. Pharm. Pat. Anal. 2016, 5, 169–181. [Google Scholar] [CrossRef]
- Rajendran, P.; Rengarajan, T.; Nandakumar, N.; Divya, H.; Nishigaki, I. Mangiferin in Cancer Chemoprevention and Treatment: Pharmacokinetics and Molecular Targets. J. Recept. Signal Transduct. 2015, 35, 76–84. [Google Scholar] [CrossRef]
- Louisa, M.; Soediro, T.M.; Suyatna1, F.D. In Vitro Modulation of P-Glycoprotein, MRP-1 and BCRP Expression by Mangiferin in Doxorubicin-Treated MCF-7 Cells. Asian Pac. J Cancer Prev. 2014, 15, 1639–1642. [Google Scholar] [CrossRef] [Green Version]
- Takeda, T.; Tsubaki, M.; Kino, T.; Kawamura, A.; Isoyama, S.; Itoh, T.; Imano, M.; Tanabe, G.; Muraoka, O.; Matsuda, H.; et al. Mangiferin Enhances the Sensitivity of Human Multiple Myeloma Cells to Anticancer Drugs through Suppression of the Nuclear Factor ΚB Pathway. Int. J. Oncol. 2016, 48, 2704–2712. [Google Scholar] [CrossRef] [Green Version]
- Ajuwon, O.R.; Ayeleso, A.O.; Adefolaju, G.A. The Potential of South African Herbal Tisanes, Rooibos and Honeybush in the Management of Type 2 Diabetes Mellitus. Molecules 2018, 23, 3207. [Google Scholar] [CrossRef] [Green Version]
- Dube, P.; Meyer, S.; Marnewick, J.L. Antimicrobial and Antioxidant Activities of Different Solvent Extracts from Fermented and Green Honeybush (Cyclopia Intermedia) Plant Material. S. Afr. J. Bot. 2017, 110, 184–193. [Google Scholar] [CrossRef]
- Magcwebeba, T.U.; Swart, P.; Swanevelder, S.; Joubert, E.; Gelderblom, W.C.A. In Vitro Chemopreventive Properties of Green Tea, Rooibos and Honeybush Extracts in Skin Cells. Molecules 2016, 21, 1622. [Google Scholar] [CrossRef]
- Aboyewa, J.A.; Sibuyi, N.R.S.; Meyer, M.; Oguntibeju, O.O. Gold Nanoparticles Synthesized Using Extracts of Cyclopia Intermedia, Commonly Known as Honeybush, Amplify the Cytotoxic Effects of Doxorubicin. Nanomaterials 2021, 11, 132. [Google Scholar] [CrossRef] [PubMed]
- Patra, J.K.; Das, G.; Fraceto, L.F.; Campos, E.V.R.; Rodriguez-Torres, M.D.P.; Acosta-Torres, L.S.; Diaz-Torres, L.A.; Grillo, R.; Swamy, M.K.; Sharma, S.; et al. Nano Based Drug Delivery Systems: Recent Developments and Future Prospects. J. Nanobiotechnol. 2018, 16, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, S.; Sau, S.; Madhuri, D.; Bollu, V.S.; Madhusudana, K.; Sreedhar, B.; Banerjee, R.; Patra, C.R. Green Synthesis and Characterization of Monodispersed Gold Nanoparticles: Toxicity Study, Delivery of Doxorubicin and Its Bio-Distribution in Mouse Model. J. Biomed. Nanotechnol. 2016, 12, 165–181. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Konorev, E.A.; Kotamraju, S.; Joseph, J.; Kalivendi, S.; Kalyanaraman, B. Doxorubicin Induces Apoptosis in Normal and Tumor Cells via Distinctly Different Mechanisms: Intermediacy of H2O2- and p53-Dependent Pathways. J. Biol. Chem. 2004, 279, 25535–25543. [Google Scholar] [CrossRef] [Green Version]
- Keeney, J.T.R.; Ren, X.; Warrier, G.; Noel, T.; Powell, D.K.; Brelsfoard, J.M.; Sultana, R.; Saatman, K.E.; St.Clair, D.K.; Butterfield, D.A. Doxorubicin-Induced Elevated Oxidative Stress and Neurochemical Alterations in Brain and Cognitive Decline: Protection by MESNA and Insights into Mechanisms of Chemotherapy-Induced Cognitive Impairment (“chemobrain”). Oncotarget 2018, 9, 30324–30339. [Google Scholar] [CrossRef] [Green Version]
- Cappetta, D.; De Angelis, A.; Sapio, L.; Prezioso, L.; Illiano, M.; Quaini, F.; Rossi, F.; Berrino, L.; Naviglio, S.; Urbanek, K. Oxidative Stress and Cellular Response to Doxorubicin: A Common Factor in the Complex Milieu of Anthracycline Cardiotoxicity. Oxid. Med. Cell. Longev. 2017, 2017, 1521020. [Google Scholar] [CrossRef]
- Meredith, A.M.; Dass, C.R. Increasing Role of the Cancer Chemotherapeutic Doxorubicin in Cellular Metabolism. J. Pharm. Pharmacol. 2016, 68, 729–741. [Google Scholar] [CrossRef] [Green Version]
- Christowitz, C.; Davis, T.; Isaacs, A.; Van Niekerk, G.; Hattingh, S.; Engelbrecht, A.M. Mechanisms of Doxorubicin-Induced Drug Resistance and Drug Resistant Tumour Growth in a Murine Breast Tumour Model. BMC Cancer 2019, 19, 757. [Google Scholar] [CrossRef] [Green Version]
- Sonowal, H.; Pal, P.B.; Wen, J.-J.; Awasthi, S.; Ramana, K.V.; Srivastava, S.K. Aldose Reductase Inhibitor Increases Doxorubicin-Sensitivity of Colon Cancer Cells and Decreases Cardiotoxicity. Sci. Rep. 2017, 7, 3182. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Ding, X.; Shen, J.; Zhang, X.; Ding, X.; Xu, B. Use of Liposomal Doxorubicin for Adjuvant Chemotherapy of Breast Cancer in Clinical Practice. J. Zhejiang Univ. B Biomed. Biotechnol. 2017, 18, 15–26. [Google Scholar] [CrossRef]
- Gyöngyösi, M.; Lukovic, D.; Zlabinger, K.; Spannbauer, A.; Gugerell, A.; Pavo, N.; Traxler, D.; Pils, D.; Maurer, G.; Jakab, A.; et al. Liposomal Doxorubicin Attenuates Cardiotoxicity via Induction of Interferon-Related DNA Damage Resistance. Cardiovasc. Res. 2020, 116, 970–982. [Google Scholar] [CrossRef] [PubMed]
- Sibuyi, N.R.S.; Thovhogi, N.; Gabuza, K.B.; Meyer, M.D.; Drah, M.; Onani, M.O.; Skepu, A.; Madiehe, A.M.; Meyer, M. Peptide-Functionalized Nanoparticles for the Selective Induction of Apoptosis in Target Cells. Nanomedicine 2017, 12, 1631–1645. [Google Scholar] [CrossRef] [PubMed]
- Badmus, J.A.; Ekpo, O.E.; Sharma, J.R.; Sibuyi, N.R.S.; Meyer, M.; Hussein, A.A.; Hiss, D.C. An Insight into the Mechanism of Holamine- and Funtumine-Induced Cell Death in Cancer Cells. Molecules 2020, 25, 5716. [Google Scholar] [CrossRef]
- Alimoradi, H.; Greish, K.; Barzegar-Fallah, A.; Alshaibani, L.; Pittalà, V. Nitric Oxide-Releasing Nanoparticles Improve Doxorubicin Anticancer Activity. Int. J. Nanomed. 2018, 13, 7771–7787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, M.; Essack, M.; Kanyanda, S.; Rees, J.G. A Low-Cost Flow Cytometric Assay for the Detection and Quantification of Apoptosis Using an Anionic Halogenated Fluorescein Dye. Biotechniques 2008, 45, 317–320. [Google Scholar] [CrossRef]
- Ahamed, M.; Akhtar, M.J.; Khan, M.A.M.; Alrokayan, S.A.; Alhadlaq, H.A. Oxidative Stress Mediated Cytotoxicity and Apoptosis Response of Bismuth Oxide (Bi2O3) Nanoparticles in Human Breast Cancer (MCF-7) Cells. Chemosphere 2019, 216, 823–831. [Google Scholar] [CrossRef]
- Rafehi, H.; Orlowski, C.; Georgiadis, G.T.; Ververis, K.; El-Osta, A.; Karagiannis, T.C. Clonogenic Assay: Adherent Cells. J. Vis. Exp. 2011, 49, e2573. [Google Scholar] [CrossRef] [Green Version]
- Xiong, S.; Xiao, G.W. Reverting Doxorubicin Resistance in Colon Cancer by Targeting a Key Signaling Protein, Steroid Receptor Coactivator. Exp. Ther. Med. 2018, 15, 3751–3758. [Google Scholar] [CrossRef]
- Shlomovitz, I.; Speir, M.; Gerlic, M. Flipping the Dogma—Phosphatidylserine in Non-Apoptotic Cell Death. Cell Commun. Signal. 2019, 17, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ramos-silva, A.; Tavares-carreón, F.; Figueroa, M.; Torre-zavala, S. De Anticancer Potential of Thevetia Peruviana Fruit Methanolic Extract. BMC Complement. Altern. Med. 2017, 17, 241. [Google Scholar] [CrossRef]
- Monier, B.; Suzanne, M. Orchestration of Force Generation and Nuclear Collapse in Apoptotic Cells. Int. J. Mol. Sci. 2021, 22, 10257. [Google Scholar] [CrossRef]
- Ribeiro, S.C.; Muratori, M.; De Geyter, M.; De Geyter, C. TUNEL Labeling with BrdUTP/Anti-BrdUTP Greatly Underestimates the Level of Sperm DNA Fragmentation in Semen Evaluation. PLoS ONE 2017, 12, e181802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirangelo, I.; Liccardo, M.; Iannuzzi, C. Hydroxytyrosol Prevents Doxorubicin-Induced Oxidative Stress and Apoptosis in Cardiomyocytes. Antioxidants 2022, 11, 1087. [Google Scholar] [CrossRef] [PubMed]
- Pilco-Ferreto, N.; Calaf, G.M. Influence of Doxorubicin on Apoptosis and Oxidative Stress in Breast Cancer Cell Lines. Int. J. Oncol. 2016, 49, 753–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubiliūtė, R.; Šulskytė, I.; Daniūnaitė, K.; Daugelavičius, R.; Jarmalaitė, S. Molecular Features of Doxorubicin-Resistance Development in Colorectal Cancer CX-1 Cell Line. Medicina 2016, 52, 298–306. [Google Scholar] [CrossRef]
- Khaleel, S.A.; Al-abd, A.M.; Ali, A.A.; Abdel-naim, A.B. Didox and Resveratrol Sensitize Colorectal Cancer Cells to Doxorubicin via Activating Apoptosis and Ameliorating P-Glycoprotein Activity. Sci. Rep. 2016, 6, 36855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Małek, A.; Taciak, B.; Sobczak, K.; Grzelak, A.; Wójcik, M.; Mieczkowski, J.; Lechowski, R.; Zabielska-Koczywaş, K.A. Enhanced Cytotoxic Effect of Doxorubicin Conjugated to Glutathione-Stabilized Gold Nanoparticles in Canine Osteosarcoma—in Vitro Studies. Molecules 2021, 26, 3487. [Google Scholar] [CrossRef]
- Wang, L.; Mercer, R.R.; Rojanasakul, Y.; Qiu, A.; Lu, Y.; Scabilloni, J.F.; Wu, N.; Castranova, V. Direct Fibrogenic Effects of Dispersed Single-Walled Carbon Nanotubes on Human Lung Fibroblasts. J. Toxicol. Environ. Health Part A 2010, 73, 410–422. [Google Scholar] [CrossRef]
- Mercer, R.R.; Scabilloni, J.; Wang, L.; Kisin, E.; Murray, A.R.; Schwegler-Berry, D.; Shvedova, A.A.; Castranova, V. Alteration of Deposition Pattern and Pulmonary Response as a Result of Improved Dispersion of Aspirated Single-Walled Carbon Nanotubes in a Mouse Model. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 294, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Manke, A.; Wang, L.; Rojanasakul, Y. Mechanisms of Nanoparticle-Induced Oxidative Stress and Toxicity. Biomed. Res. Int. 2013, 2013, 942916. [Google Scholar] [CrossRef]
- He, H.; Wang, L.; Qiao, Y.; Zhou, Q.; Li, H.; Chen, S.; Yin, D.; Huang, Q.; He, M. Doxorubicin Induces Endotheliotoxicity and Mitochondrial Dysfunction via ROS/ENOS/NO Pathway. Front. Pharmacol. 2020, 10, 1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terasaki, A.; Kurokawa, H.; Ito, H.; Komatsu, Y.; Matano, D.; Terasaki, M.; Bando, H.; Hara, H.; Matsui, H. Elevated Production of Mitochondrial Reactive Oxygen Species via Hyperthermia Enhanced Cytotoxic Effect of Doxorubicin in Human Breast Cancer Cell Lines MDA-MB-453 and MCF-7. Int. J. Mol. Sci. 2020, 21, 9522. [Google Scholar] [CrossRef] [PubMed]
- Omoruyi, S.I.; Ekpo, O.E.; Semenya, D.M.; Jardine, A.; Prince, S. Exploitation of a Novel Phenothiazine Derivative for Its Anti—Cancer Activities in Malignant Glioblastoma. Apoptosis 2020, 25, 261–274. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aboyewa, J.A.; Sibuyi, N.R.S.; Goboza, M.; Murtz, L.-A.; Oguntibeju, O.O.; Meyer, M. Co-Treatment of Caco-2 Cells with Doxorubicin and Gold Nanoparticles Produced from Cyclopia intermedia Extracts or Mangiferin Enhances Drug Effects. Nanomaterials 2022, 12, 3918. https://doi.org/10.3390/nano12213918
Aboyewa JA, Sibuyi NRS, Goboza M, Murtz L-A, Oguntibeju OO, Meyer M. Co-Treatment of Caco-2 Cells with Doxorubicin and Gold Nanoparticles Produced from Cyclopia intermedia Extracts or Mangiferin Enhances Drug Effects. Nanomaterials. 2022; 12(21):3918. https://doi.org/10.3390/nano12213918
Chicago/Turabian StyleAboyewa, Jumoke A., Nicole R. S. Sibuyi, Mediline Goboza, Lee-Ann Murtz, Oluwafemi O. Oguntibeju, and Mervin Meyer. 2022. "Co-Treatment of Caco-2 Cells with Doxorubicin and Gold Nanoparticles Produced from Cyclopia intermedia Extracts or Mangiferin Enhances Drug Effects" Nanomaterials 12, no. 21: 3918. https://doi.org/10.3390/nano12213918