
Prussian Blue Nanoparticle-Mediated Scalable Thermal Stimulation for In Vitro Neuronal Differentiation

 , , , , , and
, , , , , and
Abstract
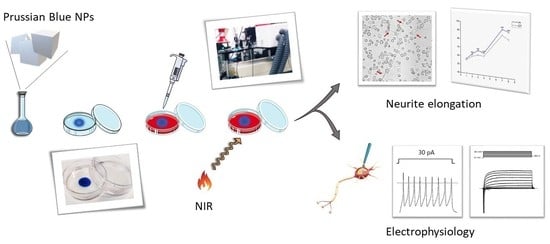
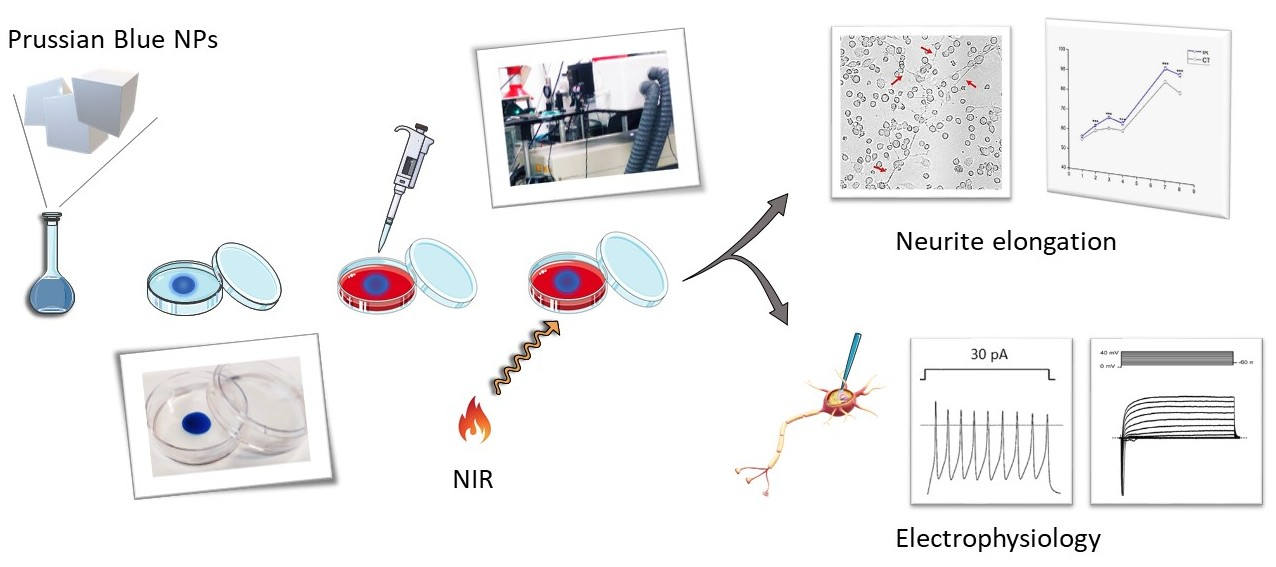
:
1. Introduction
2. Materials and Methods
2.1. Cell Cultures
2.2. Bulk Heating Protocol
2.3. Prussian Blue Nanoparticle Preparation
2.4. PBNP-PVA Layer Preparation
2.5. Irradiation Protocol by Heating Nanoparticle Layers
2.6. Morphological Analysis
2.7. Electrophysiological Analysis
2.8. Lactate-Dehydrogenase (LDH) Assay
2.9. Statistical Analysis
3. Results
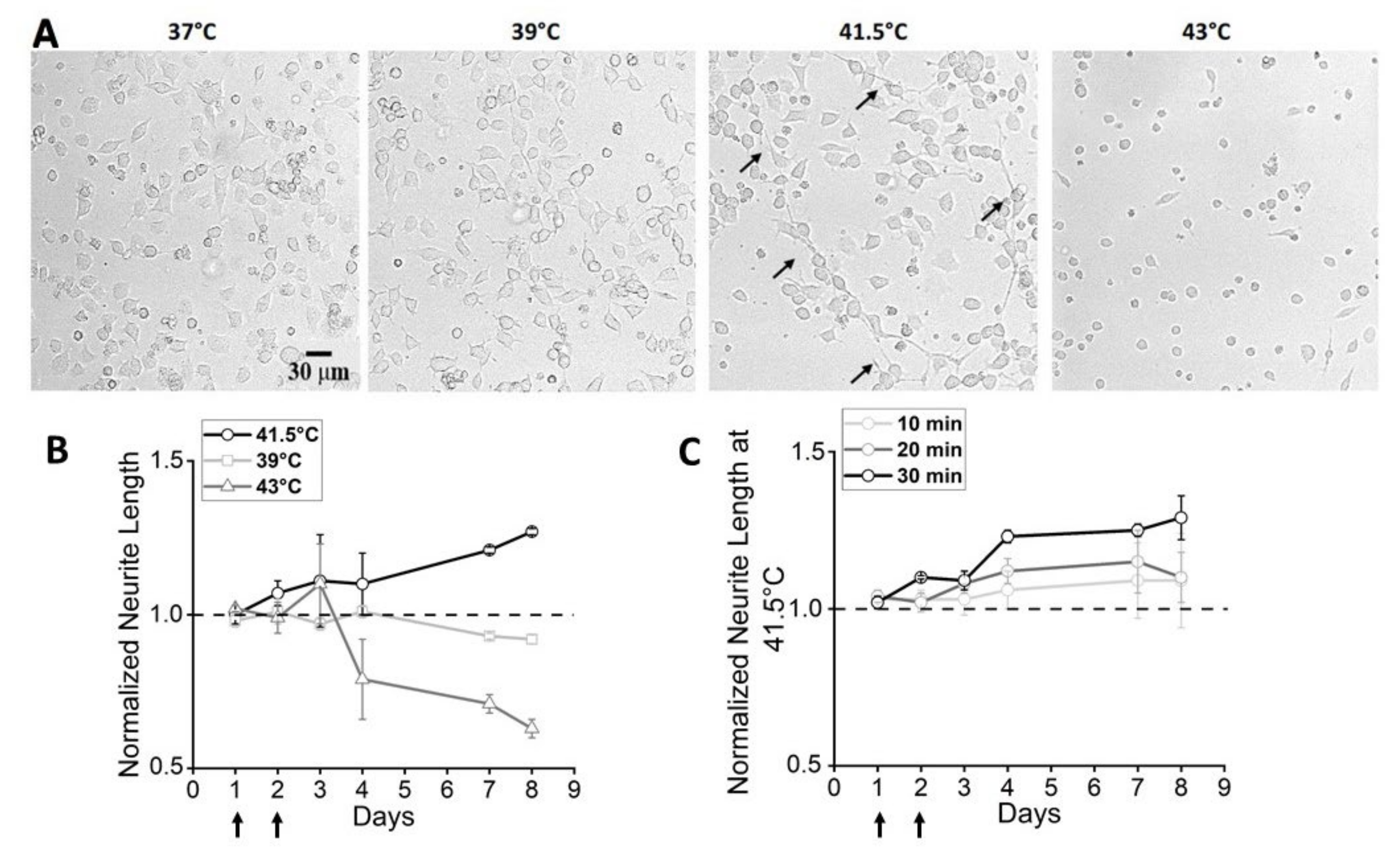
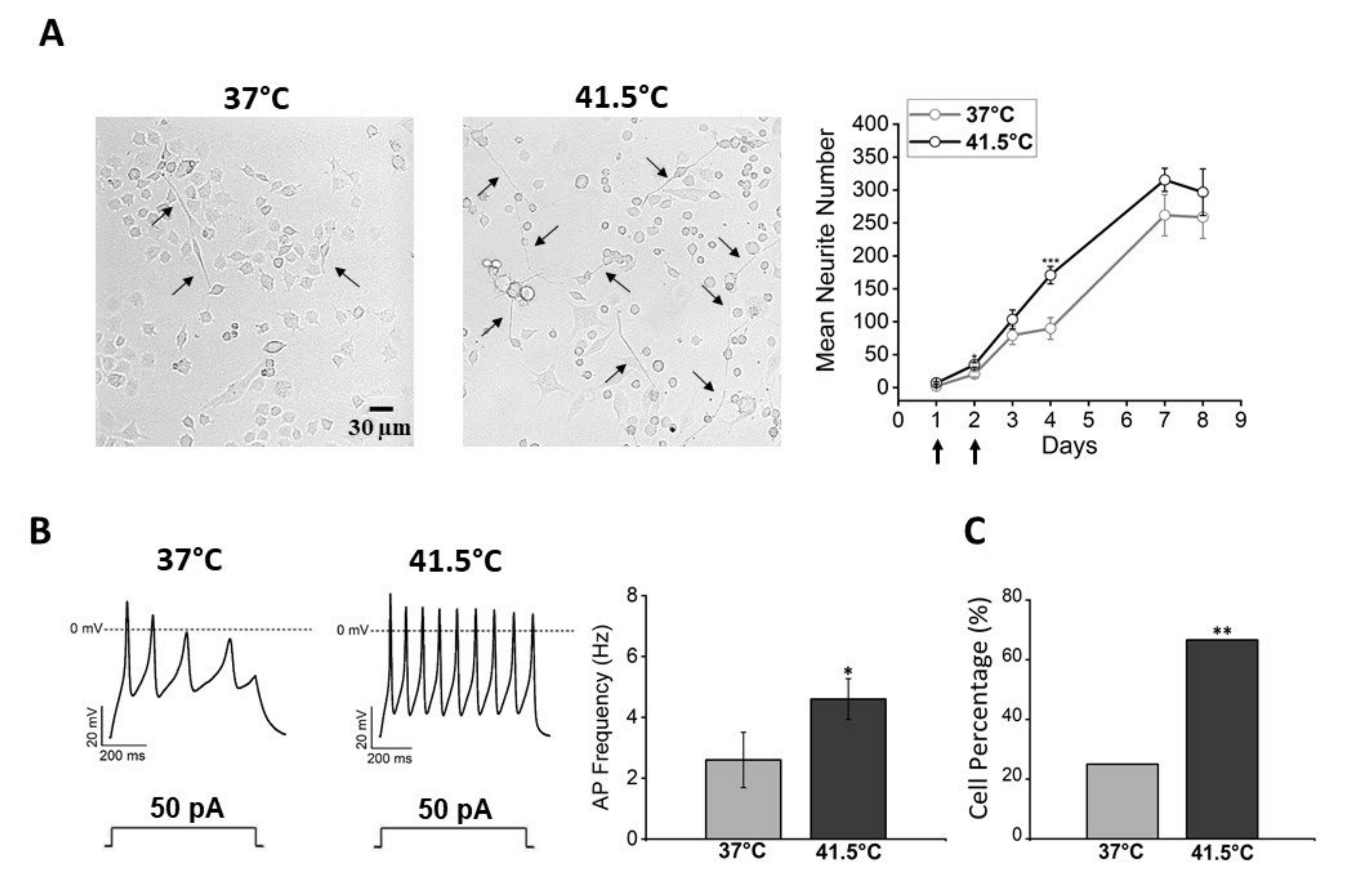
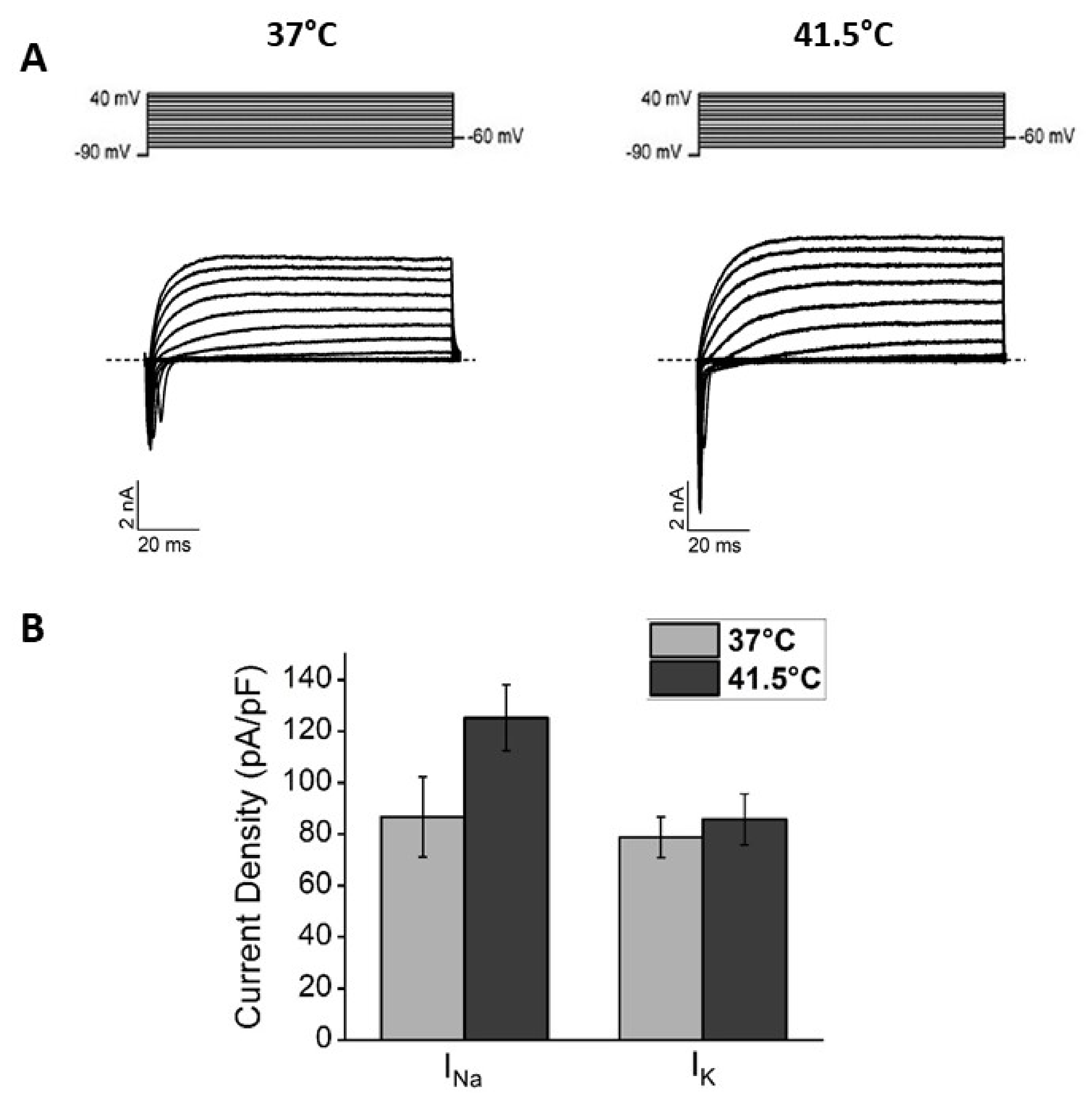
3.1. Effects of Bulk Heating: Morphological and Functional Characterization
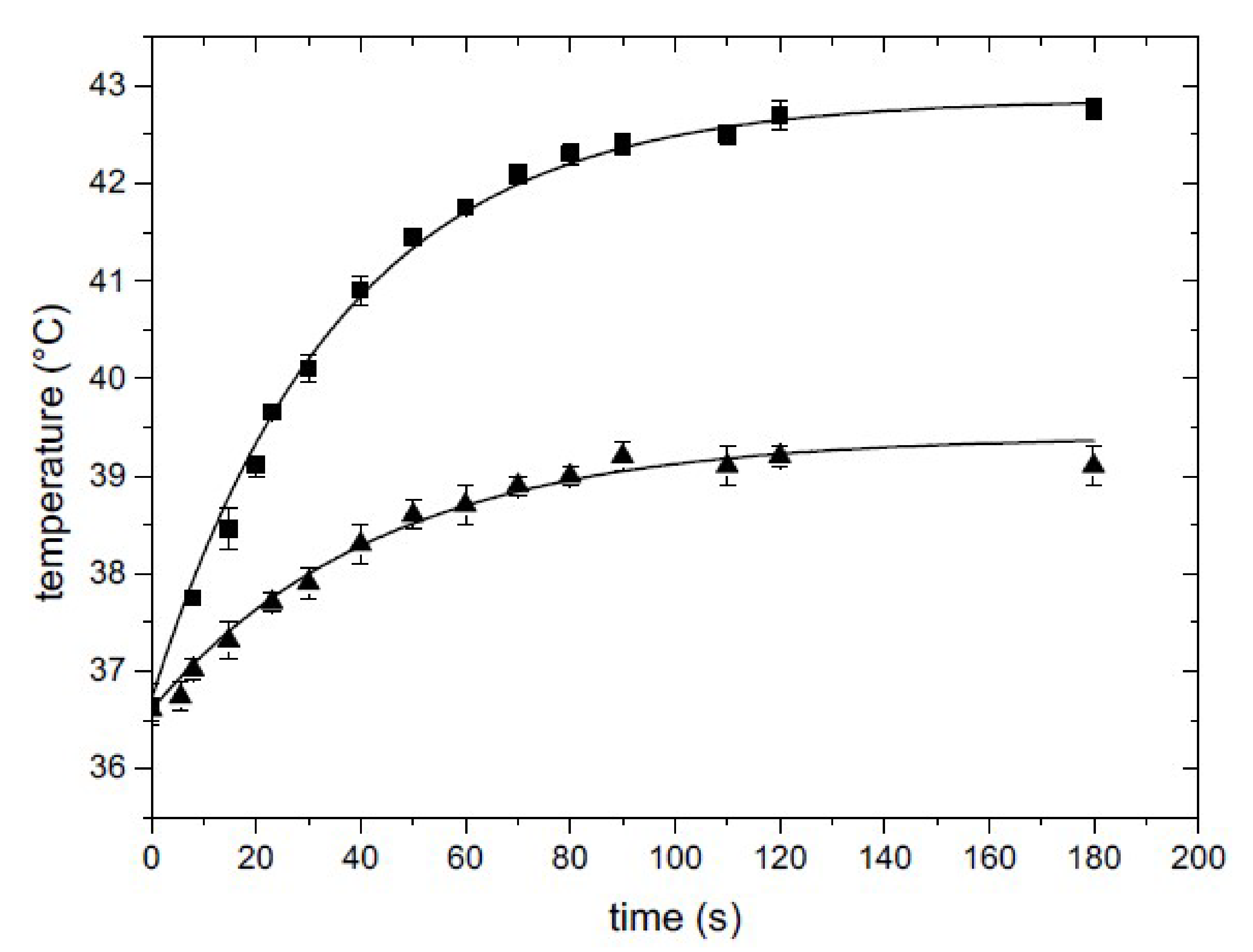
3.2. Smart Petri Dish Characterization
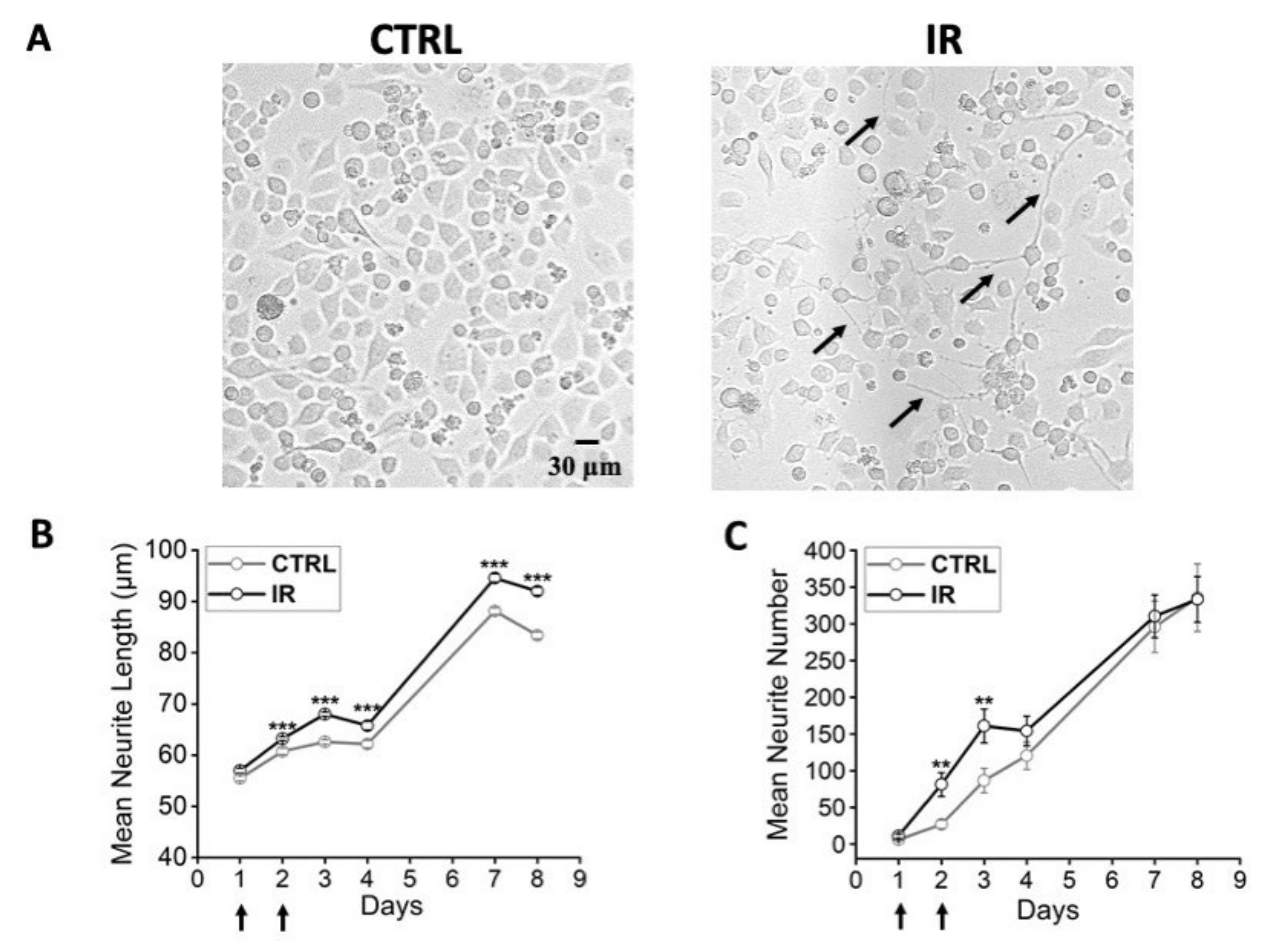
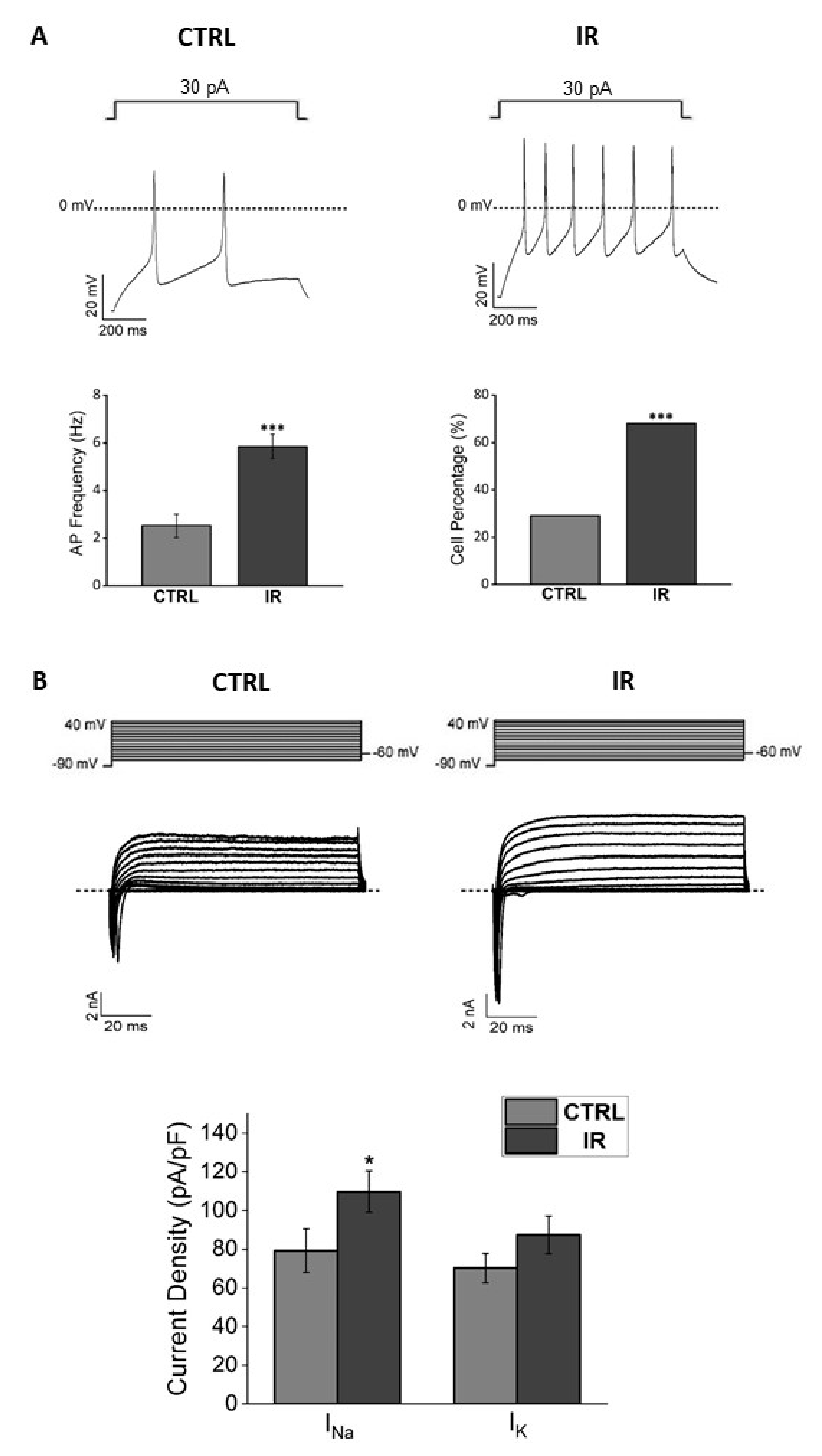
3.3. Effects of Thermal Increase by PBNP Irradiation
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shapiro, M.; Homma, K.; Villarreal, S.; Richter, C.-P.; Benazilla, P. Infrared light excites cells by changing their electrical capacitance. Nat. Commun. 2012, 3, 736. [Google Scholar] [CrossRef] [PubMed]
- Akhavan, O.; Ghaderi, E.; Shirazian, S.A. Near infrared laser stimulation of human neural stem cells into neurons on graphene nanomesh semiconductors. Colloids Surf. B Biointerfaces 2015, 126, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Kudo, T.-A.; Kanetaka, H.; Mochizuki, K.; Tominami, K.; Nunome, S.; Abe, G.; Kosukegawa, H.; Abe, T.; Mori, H.; Mori, K.; et al. Induction of Neurite Outgrowth in PC12 Cells Treated with Temperature-Controlled Repeated Thermal Stimulation. PLoS ONE 2015, 10, e0124024. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.E.; Matsuzaki, K.; Katakura, M.; Sugimoto, N.; Mamun, A.A.; Islam, R.; Hashimoto, M.; Shido, O. Direct exposure to mild heat promotes proliferation and neuronal differentiation of neural stem/progenitor cells in vitro. PLoS ONE 2017, 12, e0190356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thang, D.C.; Wang, Z.; Lu, X.; Xing, B. Precise cell behaviors manipulation through light-responsive nano-regulators: Recent advance and perspective. Theranostics 2017, 9, 3308–3340. [Google Scholar] [CrossRef] [PubMed]
- Paviolo, C.; Haycock, J.W.; Yong, J.; Yu, A.; Stoddart, P.R.; McArthur, S.L. Laser exposure of gold nanorods can increase neuronal cell outgrowth. Biotechnol. Bioeng. 2013, 110, 2277–2291. [Google Scholar] [CrossRef]
- Paviolo, C.; Thompson, A.C.; Yong, J.; Brown, W.G.; Stoddart, P.R. Nanoparticle-enhanced infrared neural stimulation. J. Neural Eng. 2014, 11, 065002. [Google Scholar] [CrossRef]
- Paviolo, C.; Stoddart, P.R. Metallic nanoparticles for peripheral nerve regeneration: Is it a feasible approach? Neural Regen. Res. 2015, 10, 1065–1066. [Google Scholar]
- Eom, K.; Kim, J.; Choi, J.M.; Kang, T.; Chang, J.W.; Byun, K.M.; Jun, S.B.; Kim, S.J. Enhanced Infrared Neural Stimulation using Localized Surface Plasmon Resonance of Gold Nanorods. Small 2014, 10, 3853–3857. [Google Scholar] [CrossRef]
- Yong, J.; Needham, K.; Brown, W.G.; Nayagam, B.A.; McArthur, S.L.; Yu, A.; Stoddart, P.R. Gold-nanorod-assisted near-infrared stimulation of primary auditory neurons. Adv. Healthc. Mater. 2014, 3, 1862–1868. [Google Scholar] [CrossRef]
- Pallavicini, P.; Chirico, G.; Taglietti, A. Harvesting light to produce heat: Photothermal nanoparticles for technological applications and biomedical devices. Chem. Eur. J. 2021, 27, 15361–15374. [Google Scholar] [CrossRef] [PubMed]
- Weissleder, R. A clearer vision for in vivo imaging. Nat. Biotechnol. 2001, 19, 316–317. [Google Scholar] [CrossRef]
- Wang, L.; Meng, D.; Hao, Y.; Zhao, Y.; Li, D.; Zhang, B.; Zhang, Y.; Zhang, Z. Gold nanostars mediated combined photothermal and photodynamic therapy and X-ray imaging for cancer theranostic applications. J. Biomater. Appl. 2015, 30, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Lewinski, N.; Colvin, V.; Drezek, R. Cytotoxicity of Nanoparticles. Small 2015, 4, 26–49. [Google Scholar] [CrossRef] [PubMed]
- Patra, C.R. Prussian blue nanoparticles and their analogues for application to cancer theranostics. Nanomedicine 2016, 11, 569–572. [Google Scholar] [CrossRef] [Green Version]
- Dacarro, G.; Grisoli, P.; Borzenkov, M.; Milanese, C.; Fratini, E.; Ferraro, G.; Taglietti, A.; Pallavicini, P. Self-assembled monolayers of Prussian blue nanoparticles with photothermal effect. Supramol. Chem. 2017, 29, 823–833. [Google Scholar] [CrossRef]
- Dacarro, G.; Taglietti, A.; Pallavicini, P. Prussian Blue Nanoparticles as a Versatile Photothermal Tool. Molecules 2018, 23, 1414. [Google Scholar] [CrossRef] [Green Version]
- Borzenkov, M.; D’Alfonso, L.; Polissi, A.; Sperandeo, P.; Collini, M.; Dacarro, G.; Taglietti, A.; Chirico, G.; Pallavicini, P. Novel photothermally active polyvinyl alcohol-prussian blue nanoparticles hydrogel films capable of eradicating bacteria and mitigating biofilms. Nanotechnology 2019, 30, 295702. [Google Scholar] [CrossRef]
- Gautam, M.; Poudel, K.; Yong, C.S.; Kim, O.J. Prussian blue nanoparticles: Synthesis, surface modification, and application in cancer treatment. Int. J. Pharm. 2018, 549, 31–49. [Google Scholar] [CrossRef]
- Platika, D.; Boulos, M.H.; Baizer, L.; Fishman, C.M. Neuronal traits of clonal cell lines derived by fusion of dorsal root ganglia neurons with neuroblastoma cells. Proc. Natl. Acad. Sci. USA 1985, 82, 3499–3503. [Google Scholar] [CrossRef] [Green Version]
- Pastori, V.; D’Aloia, A.; Blasa, S.; Lecchi, M. Serum-deprived differentiated neuroblastoma F-11 cells express functional dorsal root ganglion neuron properties. PeerJ 2019, 7, e7951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobner, D. Comparison of the LDH and MTT assays for quantifying cell death: Validity for neuronal apoptosis? J Neurosci. Methods 2000, 96, 147–152. [Google Scholar] [CrossRef]
- Koh, J.Y.; Choi, D.W. Quantitative determination of glutamate mediated cortical neuronal injury in cell culture by lactate dehydrogenase efflux assay. J. Neurosci. Methods 1987, 20, 83–90. [Google Scholar] [CrossRef]
- Russo, L.; Sgambato, A.; Lecchi, M.; Pastori, V.; Raspanti, M.; Natalello, A.; Doglia, S.M.; Nicotra, F.; Cipolla, L. Neoglucosylated collagen matrices drive neuronal cells to differentiate. ACS Chem. Neurosci. 2014, 5, 261–265. [Google Scholar] [CrossRef] [Green Version]
- Wieringa, P.; Tonazzini, I.; Micera, S.; Cecchini, M. Nanotopography induced contact guidance of the F11 cell line during neuronal differentiation: A neuronal model cell line for tissue scaffold development. Nanotechnology 2012, 27. [Google Scholar] [CrossRef]
- Beal McIlvain, H.; Baudy, A.; Sullivan, K.; Liu, D.; Pong, K.; Fennell, M.; Dunlop, J. Pituitary adenylate cyclase-activating peptide (PACAP) induces differentiation in the neuronal F11 cell line through a PKA-dependent pathway. Brain Res. 2006, 1077, 16–23. [Google Scholar] [CrossRef]
- Carvalho-de-Souza, J.L.; Treger, J.S.; Dang, B.; Kent, S.B.H.; Pepperberg, D.R.; Bezanilla, F. Photosensitivity of Neurons Enabled by Cell-Targeted Gold Nanoparticles. Neuron 2015, 86, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.; Harris, N.; Niyonshuti, I.I.; Jenkins, S.V.; Hayar, A.M.; Watanabe, F.; Jamshidi-Parsian, A.; Chen, J.; Borrelli, M.J.; Griffin, R.J. Photothermal Response Induced by Nanocage-Coated Artificial Extracellular Matrix Promotes Neural Stem Cell Differentiation. Nanomaterials 2021, 11, 1216. [Google Scholar] [CrossRef]
- Wang, S.J.; Chen, C.S.; Chen, L.C. Prussian blue nanoparticles as nanocargoes for delivering DNA drugs to cancer cells. Sci. Technol. Adv. Mater. 2013, 14, 044405. [Google Scholar] [CrossRef] [Green Version]
- Hussain, G.; Wang, J.; Rasul, A.; Anwar, H.; Qasim, M.; Zafar, S.; Aziz, N.; Razzaq, A.; Hussain, R.; de Aguilar, J.L.G.; et al. Current Status of Therapeutic Approaches against Peripheral Nerve Injuries: A Detailed Story from Injury to Recovery. Int. J. Biol. Sci. 2020, 16, 116–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maimon, B.E.; Zorzos, A.N.; Bendell, R.; Harding, A.; Fahmi, M.; Srinivasan, S.; Calvaresi, P.; Herr, H.M. Transdermal optogenetic peripheral nerve stimulation. J. Neural Eng. 2017, 14, 034002. [Google Scholar] [CrossRef] [PubMed]
- Geuna, S.; Raimondo, S.; Fregnan, F.; Haastert-Talini, K.; Grothe, C. In vitro models for peripheral nerve regeneration. Eur. J. Neurosci. 2016, 43, 287–296. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bulk Heating (Days 7–8) | |||
|---|---|---|---|
| 37 °C | S.E. | 41.5 °C | S.E. |
| 4.98 × 10−8 | 1.28 × 10−8 | 4.86 × 10−8 | 1.16 × 10−8 |
| Thermal Stimulation (Days 7–8) | |||
|---|---|---|---|
| CTRL | S.E. | IR | S.E. |
| 9 × 10−8 | 1.2 × 10−8 | 15 × 10−8 | 2.3 × 10−8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blasa, S.; Borzenkov, M.; Pastori, V.; Doveri, L.; Pallavicini, P.; Chirico, G.; Lecchi, M.; Collini, M. Prussian Blue Nanoparticle-Mediated Scalable Thermal Stimulation for In Vitro Neuronal Differentiation. Nanomaterials 2022, 12, 2304. https://doi.org/10.3390/nano12132304
Blasa S, Borzenkov M, Pastori V, Doveri L, Pallavicini P, Chirico G, Lecchi M, Collini M. Prussian Blue Nanoparticle-Mediated Scalable Thermal Stimulation for In Vitro Neuronal Differentiation. Nanomaterials. 2022; 12(13):2304. https://doi.org/10.3390/nano12132304
Chicago/Turabian StyleBlasa, Stefania, Mykola Borzenkov, Valentina Pastori, Lavinia Doveri, Piersandro Pallavicini, Giuseppe Chirico, Marzia Lecchi, and Maddalena Collini. 2022. "Prussian Blue Nanoparticle-Mediated Scalable Thermal Stimulation for In Vitro Neuronal Differentiation" Nanomaterials 12, no. 13: 2304. https://doi.org/10.3390/nano12132304