1. Introduction
Mars is a dry and barren planet with promising potential for future human colonisation. A Martian day consists of 24.65 h, varying surface temperatures averaging −63 °C and UV wavelengths ranging from 200 nm to 400 nm, which is comparable to the surface wavelengths that reach Earth (290–400 nm) [
1,
2,
3,
4].
As we enter the New Space Age, Mars is quickly emerging as humanities chosen destination for space exploration and colonisation. The Mars Exploration Programme consisting of previous missions including The Mars Reconnaissance Orbiter, The Mars Exploration Rovers, The Curiosity Rover, The Phoenix Lander, and in more recent times, The Opportunity Rover, have all paved the way in increasing knowledge of the Martian landscape. Current/future missions to Mars include NASA’s (National Aeronautics and Space Administration) 2020 Rover, which aims to study Martian astrobiology, the Manglyaan Orbiter 2 in 2022, and the Martian Moons Exploration in 2024, which aims to collect samples from the orbiting moons Phobos and Deimos. There are also plans to have manned missions to Mars as early as the 2030s. However, in addition to the immediate challenges posed by Mars’ climate, future colonists face another threat, the presence of sodium perchlorate within the Martian dust and soil [
5,
6].
Sodium perchlorate is an inorganic, water-soluble compound with the chemical formula NaClO
4. The Phoenix Mars Lander, which landed on the Vastitas Borealis plains of Mars in 2008, contained a wet chemistry lab which could perform in situ aqueous chemical analyses of Martian soil components. Within these soil samples, it was found that there were dissolved salts (10mM), with a 0.4–0.6% concentration of perchlorate, by mass [
5]. The presence of perchlorate in the form of NaClO
4, Mg(ClO
4)
2 and Mg(ClO
3)
2 was found in once free flowing brine steeps across the surface of Mars. This was observed by the onboard instrumentation of the Mars Reconnaissance Orbiter [
7]. While this comes as helpful information in understanding the history of the Red Planet, for future colonists aboard manned missions to Mars, this provides a significant obstacle in terms of sustaining human life on Mars [
8,
9]. The presence of sodium perchlorate within the environment provides a significant challenge to human, animal, and plant life. If ingested, perchlorates are highly hazardous to human health, eventually causing hypothyroidism [
10] within healthy individuals and developing foetuses. Perchlorate is a potent competitive inhibitor of the sodium-iodide symporter (NIS) found on the basolateral memndhuberne of thyroid cells. In doses ranging between 600 and 1000 mg it decreases the active transport of iodine into the thyroid [
11]. The challenges posed by omni-present environmental perchlorates on human health reside around the inhibition of iodine uptake within the body, which may be particularly harmful to those already deficient in iodine (roughly 35–45% of the world’s population is estimated to be deficient in iodine [
12]).
While this study has a shared terrestrial and extraterrestrial focus, we acknowledge the numerous global challenges to planetary health on Earth, including climate change, environmental destruction, war, famine, and gross social and economic injustices. This raises important ethical debates about investing Earths’ finite resources into the bioremediation and detoxification of distant planets, potentially at the expense of improving our own planetary health first. At the same time, exploration of environmental technologies and ecological solutions (including novel application of microbes) has the potential to support terrestrial systems on Earth as sodium perchlorate pollution is also a terrestrial cause for concern. Common sources of perchlorate released into the terrestrial environment come as a by-product of industrial products including missile fuel, fireworks, safety and hazard flares, and fertilizers [
13,
14,
15,
16]. On Earth, perchlorate is created by lightning discharges in the presence of chloride, with rain and snow samples gathered in Texas and Florida presenting with detectable levels of perchlorate [
17]. Naturally occurring perchlorate can be found in the Atacama Desert, Chile (0.03 wt %) [
18,
19], and Antarctica (1100 μg/kg) [
20]. Results from surveys of ground water, ice, and relatively unperturbed deserts have been used to estimate a 100,000 to 3,000,000 tonnes (110,000 to 3,310,000 tons) “global inventory” of natural perchlorate presently on Earth [
21]. Worryingly, perchlorate is a common pollutant within many public water supplies [
22], with the USA deciding, as of 2022, not to impose limits on perchlorate presence within US domestic water supplies [
23]. Due to its soluble nature, if perchlorate is deposited into water-sources or within soils, it will cause pollution and damage to the native ecosystem. Consequentially, perchlorate can build up within the tissues of both terrestrial and aquatic plants through bioaccumulation [
13]. Following the development of more sensitive analytical methods such as ion chromatography and mass spectroscopy, within the USA, perchlorate has since been identified in items such as tobacco, alfalfa, tomatoes, cow’s milk, cucumber, lettuce, soybeans and eggs [
13,
24,
25]. As such, future strategies geared towards the bioremediation of Mars, in turn, benefits Earth and its various systems which depend on safe drinking water, healthy soils and pristine ecosystems.
One way to challenge the issue of perchlorate contamination both terrestrially and extraterrestrially is through the use of bioremediation microbes. Bioremediation is the use of either native or transplanted microbial species into a polluted site to help metabolise and breakdown a pollutant into less harmful by-products. Microbial metabolism of perchlorates is a natural phenomenon common to a handful of microbial species which exist within perchlorate contaminated environments, such as
Dechloromonas aromatica and
Azospira oryzae [
26,
27,
28]. The utilisation of these microbial perchlorate reducing strains has the added potential to allow for the production of oxygen on an extraterrestrial planet, while simultaneously breaking down contaminant perchlorates. The use of bioremediation microbes to aid in the restoration of pristine environments terrestrially, the scrubbing of municipal water supplies, and the future colonisation of Mars provides a promising natural remedy to a human health and environmental crisis, with the potential to stem beyond the reduction of perchlorate within the water and regolith and exist within the field of biomanufacturing and biomining [
29,
30].
Focusing solely on the presence of sodium perchlorate due to its wide availability and consequential toxic effects on the human body if ingested, we aimed to determine if the chosen soil and sediment-dwelling [
27,
31,
32,
33] candidate strains (
Escherichia coli ATCC 25922,
Paraburkholderia fungorum DSM 17061,
Escherichia coli DSM 9079,
Deinococcus radiodurans ATCC 13939 and
Dechloromonas aromatica ATCC-BAA 1848) were capable of metabolising supplied sodium perchlorate both individually and in a co-culture.
E. coli ATCC 25922, a model test organism [
34,
35], has been shown to form a both moderate protective biofilm [
36] and can withstand extreme temperature cycles and UVC exposure [
37]. For these reasons, it was deemed a suitable test microbe chosen for this study.
P. fungorum is a well-characterised bioremediator and biofilm former, capable of bioremediating dibenzothiophene, fluorene, naphthalene and phenanthrene, in addition to polycyclic aromatic hydrocarbons (apart from condensed thiophenes) [
38]. It has also been reported that
Paraburkholderia sp., have been implicated in perchlorate reduction [
39]. Recent experimentation documents that
P. fungorum is both highly freeze-thaw and UVC resistant, capable of withstanding 20 freeze-thaw cycles in the absence of a cryoprotectant (and retaining over 70% viability) and able to withstand UVC doses of over 44,400 J/m
2 [
37].
D. radiodurans is a renowned radiation resistant microbe, known to have multiple copies of its genomes [
37,
40,
41]. It is capable of remediating heavy metals such as ionic mercury [
42], Di-n-butyl phthalate (an endocrine disruptor) [
43], and is also able to bioremediate uranium when it forms a biofilm [
44]. As there are limited studies examining how the presence of sodium perchlorate affects its biofilm forming capabilities, it was chosen as a test candidate.
D. aromatica is an established bioremediator, particularly of sodium perchlorate, capable of degrading benzene, reducing perchlorate, and oxidising chlorobenzoate, toluene, and xylene [
27]. Due to its established perchlorate reducing abilities, it served as a robust positive control for Raman-based perchlorate bioremediation studies.
With the use of Raman spectroscopy, reduction rates were examined within a minimal medium environment and within environments void of all nutrients, in an attempt to investigate the bioremediation potential, and employment of these strains, within perchlorate-polluted environments. Raman spectroscopy was chosen for this analysis as this has become established as an optical technique that generates a spectroscopic fingerprint of the constituent molecules in a sample and provides both quantitative and qualitative information [
45,
46,
47,
48].
2. Materials and Methods
2.1. Test Microorganism and Culture Preparation
E. coli ATCC strain 25922 (American Type Culture Collection, Manassas, VA, USA), E. coli DSM 9079 (DSMZ-German Collection of Microorganisms and Cell Cultures GmbH, Braunschweig, Germany), D. radiodurans ATCC strain 13939, P. fungorum DSM strain 17061 and D. aromatica ATCC-BAA strain 1848 were used in this experimental study.
Cell cultures were prepared in triplicate from −80 °C 40% glycerol stock solutions. Three streaks were performed on nutrient agar plates (Formedium, Norfolk, UK; NAO02) to isolate individual colonies. Three colonies were picked and separately incubated in 10 mL of nutrient broth (Formedium, Norfolk, UK; NBO02). All strains were incubated at 30 °C for 48 h, except for both E. coli strains which were incubated at 37 °C for 18 h.
2.2. Determination of Cell Absorbance for Standardised Testing
Perchlorate toxicity testing mirrored a standard minimum inhibitory concentration-like assay, whereby a concentration of 1 × 10
5 CFU/mL is required [
49].
Adapted from the paper by Twomey et al., (2021) a sample of E. coli ATCC strain 25922 was inoculated in a final volume of 10 mL. Following incubation, 2 mL of the overnight culture was sub-cultured into 8 mL of nutrient broth. This was incubated at the 37 °C while periodically checking the optical density (OD) at 600 nm (CamSpec M501, Crawley, UK), until an OD600 nm of 0.5 was obtained. Once an OD600 nm of 0.5 had been achieved, 20 µL of the culture strain was sub-cultured into 980 µL to fresh broth to create a final concentration of 105 CFU/mL. When tested in the UV spectrophotometer, an OD600 nm of 0.012 was achieved. Each test sample was standardised to an OD600 nm of 0.012, as a result, to achieve comparability. Note: while differences in cell-sizes are recorded, based on E. coli ATCC 25922 strain, an OD600 nm of 0.012 was chosen for all strains to ensure standardisation and absorbance comparability across test strains.
2.3. Creating Mixed Bacterial Populations
An overnight culture of each strain was standardised to a 0.5 MacFarland turbidity equivalent using a densitometer (DEN-1 Densitometer; BIOSAN; Riga, Latvia) and sterile Ringers solution. A 2 mL aliquot of the standardised test sample was placed into a collection tube. Additional 2 mL aliquots of other standardised strains were placed into the same tube to create a 1:1 co-culture ratio.
A ratio of 1:1, 1:1:1, 1:1:1:1 for bacterial cell density in solution was used throughout each combination. For example, in the 1:1 ratio, 1 mL of culture A was combined with 1 mL of culture B in order to obtain the AB combination.
The bacterial combinations were as follows with A = E. coli ATCC strain 25922, B = P. fungorum DSM strain 17061, C = D. radiodurans ATCC strain 13939, D = D. aromatica ATCC-BAA strain 1848: A, B, C, D, AB, AC, AD, BC, BD, CD, ABC, ABD, BCD, CAD, ABCD.
2.4. Raman Spectroscopy for Perchlorate Detection
2.4.1. Sample Preparation
A 0.3% sodium perchlorate solution was prepared by dissolving 3 g of sodium perchlorate (Merck: 410241-100G, Darmstadt, Germany) in 1000 mL of Davis minimal medium (Sigma Aldrich; 93753-500G-F, St. Louis, MO, USA) and autoclaved. Note: All media were formulated using RODI water to prevent any possible background contamination. A 0.3% sodium perchlorate solution was also formulated in RODI alone. At a concentration of 3000 mg/L (0.3%
w/
v) this study aims to assess the bioremedial potential of the chosen strains against levels exceeding recorded domestic perchlorate levels ((300–1000 mg) [
50]) and Mars-like perchlorate concentrations (0.4–0.6% [
5,
51]). A lower concentration of 0.3% was chosen for this preliminary study to determine the perchlorate reduction levels of the test microbes at a less-harmful lower concentration before exploring higher, more stressful, perchlorate concentrations.
Once dissolved and autoclaved a 30 mL aliquot of the perchlorate solution was placed in a 50 mL falcon tube. A 2 mL aliquot was taken from this and stored in a microcentrifuge tube to be used as a negative control. This work was carried out in triplicate.
A 1 mL aliquot of the previously formulated individual/co-culture test culture was added to the perchlorate solution and incubated at 30 °C for 48 h.
Post-incubation, a 5 mL aliquot of the metabolised sample was filtered through a 0.2 μM filter to remove any microbial life and cease the reduction of perchlorate. The filtrate was stored in a falcon tube and the perchlorate content was examined using Raman spectroscopy.
2.4.2. Maintaining Sample Integrity
To maintain the integrity of the perchlorate test media before, and after, metabolisation, several quality control steps were adhered to. Sodium perchlorate salt was always added to RODI water to ensure no other chemical contamination which could alter/influence the experimental results. Once added to the RODI water, DMM broth power was then added and autoclaved at 121 °C at 15 PSI for 15 min to ensure sterility and help with the homegenisation of the perchlorate into solution. It is noted while some perchlorate species such as potassium perchlorate and ammonium perchlorate are highly reactive to high temperatures, sodium perchlorate doesn’t degrade in temperatures lower than 490 °C [
52,
53,
54]. Perchlorate-containing broth was stored at room temperature to avoid the perchlorate falling out of solution at low-temperatures. Metabolised samples were passed through 0.2 μM filter to remove all microbial life and prevent further reduction of the sample. Stored samples were passed through the filter each time they were tested to maintain sterility.
2.4.3. Raman Spectroscopy for Perchlorate Detection
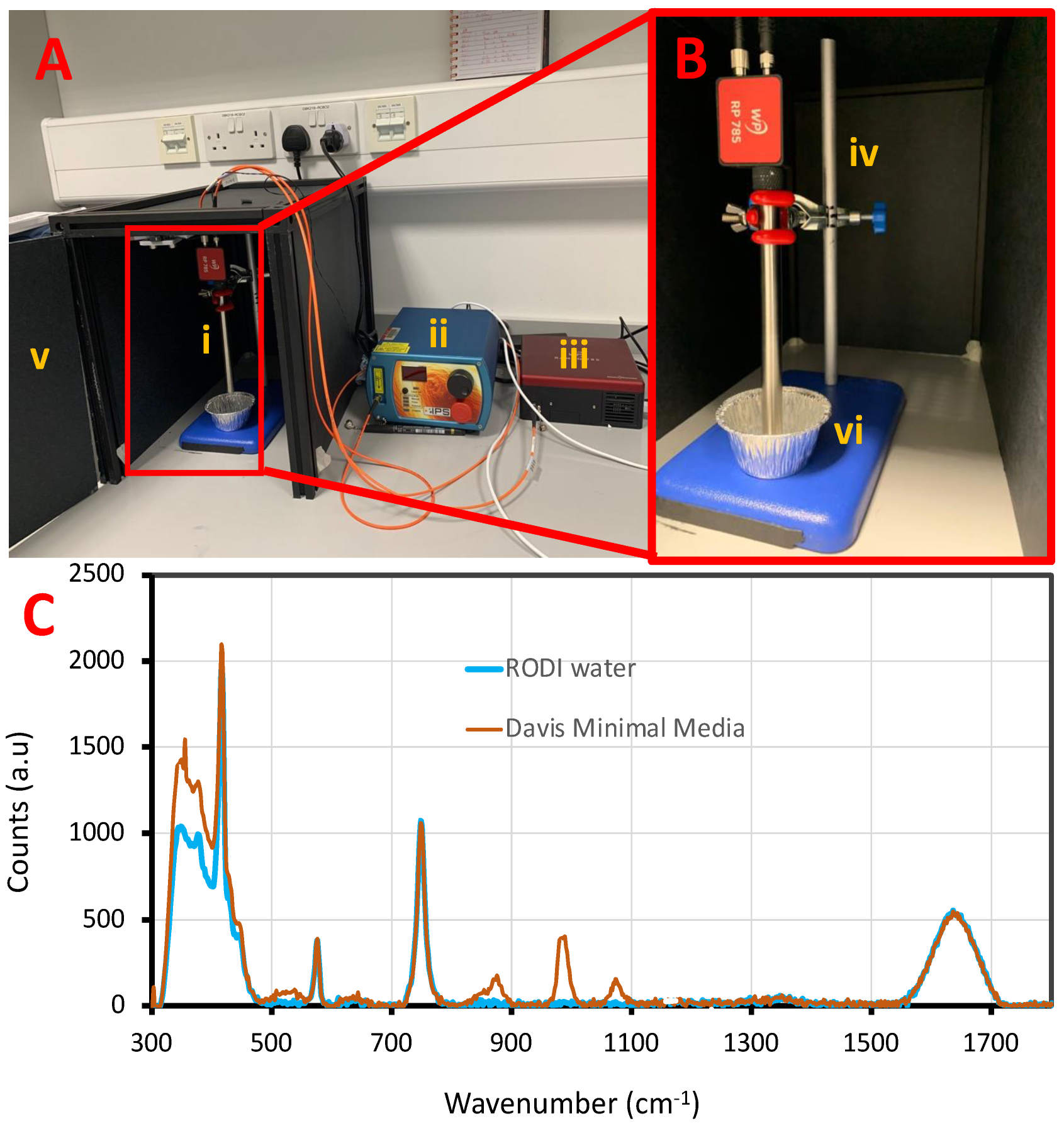
Hardware setup: The Raman spectroscopy setup consists of three main components: a Raman Optical Probe, a laser excitation source and a spectrometer as shown in
Figure 1. A Wasatch Pro (WP) Raman 785 probe was used for both excitation and collecting Raman spectra of all the analyte samples. This probe has the all the required filters and focusing optics packaged in a 12.7 mm of stainless steel barrel so that it can be submerged in liquid samples directly and can withstand a wide range of temperatures from −20 °C to +150 °C. The Raman probe also consists of optical fiber connectors to make it easy to connect to a laser source and a spectrometer.
A 350 mW laser (IPSlasers, I0785MM0350MF; Plainsborro Township, NJ, USA) with a 785 nm wavelength was employed as the excitation source. The output power of the laser source is controlled by the dial on the front panel that sets the drive current of the laser varying from 0 to 1.5 A. A 785 nm wavelength was chosen as it provides a good compromise between minimal autofluorescence and strength of the Raman signals generated. Many samples, particularly those organic in nature, tend to autofluoresce when exposed to visible laser light. Appearing as a broad background that tends to fall off at longer Raman shifts, autofluorescence signal can overwhelm the Raman spectrum, or at the very least degrade signal to noise and make Raman peaks difficult to resolve. On the other hand, longer wavelengths of excitation beyond the 785 nm lead to weaker Raman signals as Raman effect decreases quadratically with increasing wavelength [
55].
The Raman probe was mounted on a titration clamp stand. The input optical fiber connector (FC-connector) of the Raman probe was connected to the 785 laser source using a 105 μm multimode mode fiber (Thorlabs M43L01; Newton, NJ, USA). The output end (SMA-connector) of the probe was connected to the Wasatch Photonics 785 Raman spectrophotometer using a 600um core multimode fiber (Thorlabs M29L02; Newton, NJ, USA).
Aluminium foil cups were used as sample holders to reduce background interference from the sample container. A sample volume of not less than 3 mL was used for each measurement. The height of the Raman probe was carefully adjusted to allow right exposure to the sample without touching the sides or the sample holder bottom (shown in
Figure 1). A black box (Thorlabs XE25C7/M; Newton, NJ, USA) equipped with an emergency laser power interlock switch was used as an enclosure for laser safety during the experiments. Throughout the experiments, Enlighten Software version 2.7.3 (WP) was used for collecting the spectral data. In
Figure 1C, an example Raman spectra of DMM and RODI water obtained with the Raman probe set-up is shown. It is evident from the Raman peaks in the shaded regions of the
Figure 1C that these peaks are common to both DMM and RODI media. Raman peaks in the left hand shaded region (between the spectral regime of 300 cm
−1 to 800 cm
−1) corresponds to sapphire ball lens in the Raman probe are shown [
56]. On the other hand, the Raman peaks in the right-hand shaded region (between the spectral regime of 1200 cm
−1 to 1800 cm
−1) corresponds to water.
Calibration and validation using Perchlorate standards: The characterisation of the Raman spectroscopy set-up with perchlorates within both Davis minimal medium broth and RODI water was carried out and a calibration curve is plotted. All the experiments were performed in triplicates. Unmetabolised samples (6 mL in total) were combined together and placed in the clean aluminium tray. Starting concentration 0.3% perchlorate was used across all the test samples. A concentration of 0.3% was used so as to allow for the detection of perchlorate metabolism while simultaneously aiming to reduce the stress placed on the cells in the presence of perchlorate. The power of the laser was fixed by driving the laser with a current setting of 0.8 A through the front dial. For each data point, 5 Raman spectra of the unmetabolised sample were collected with an acquisition time of 3 s for each spectra. Spectral data were normalised using baseline correction. Following the unmetabolised samples, the filtered samples were examined following the same parameters and the difference in the recorded levels of sodium perchlorate present was calculated via extrapolation from the standard curve.
Data collection and processing: Enlighten Software version 2.7.3 (WP) was used to gather and process data. Settings used within the Raman spectroscope are: baseline correction with Adaptive Iteratively Reweighted Penalized Least Squares (AirPLS) enabled; integration time of 3000 ms; number of averages 5. Linear regression data analysis was carried out on GraphPad Prism version 9.4.0 with extrapolated values from the standard curves being extracted from GraphPad Prism and converted to percentage reduction using Microsoft Excel (Version 2308).
3. Results
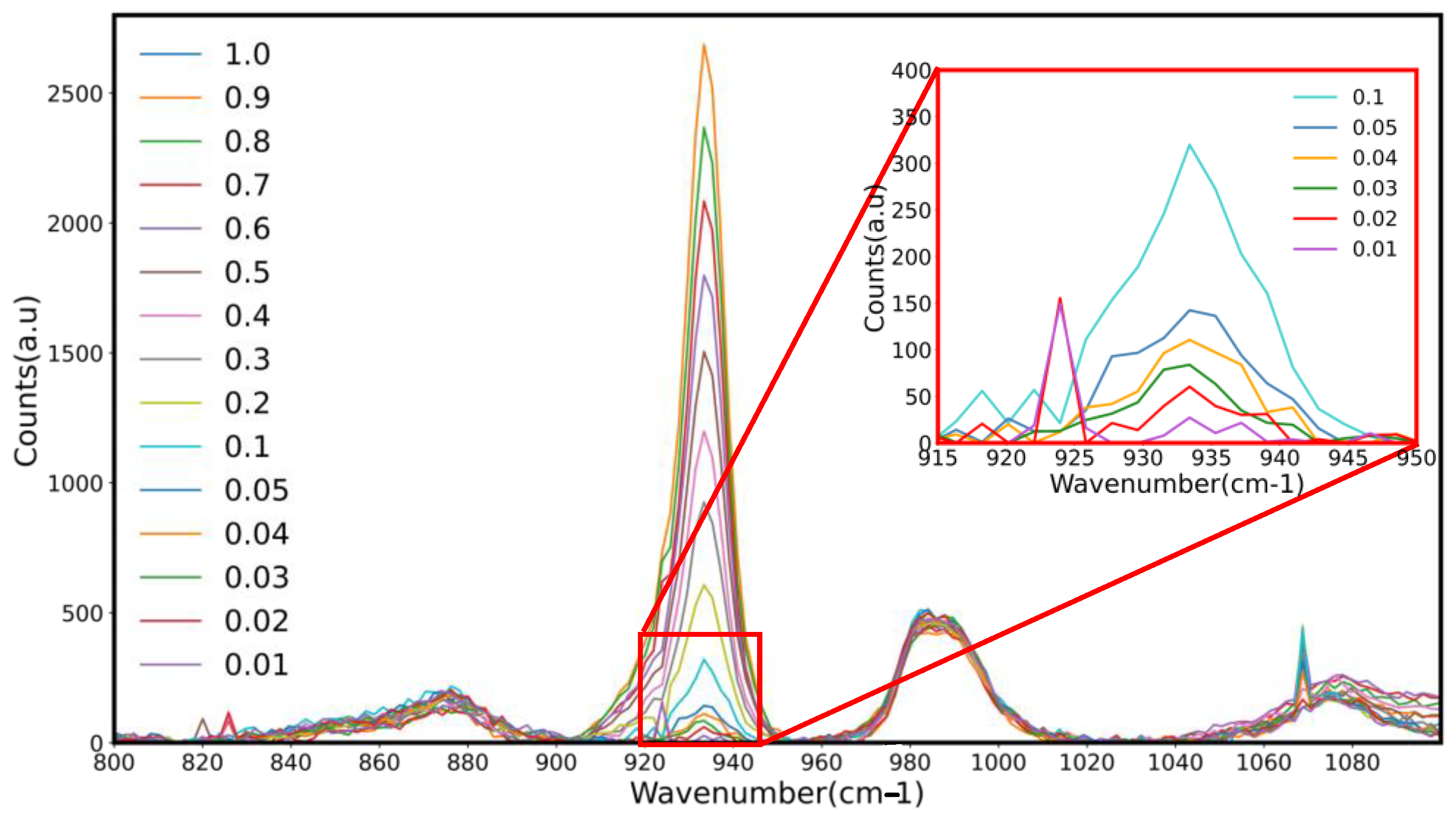
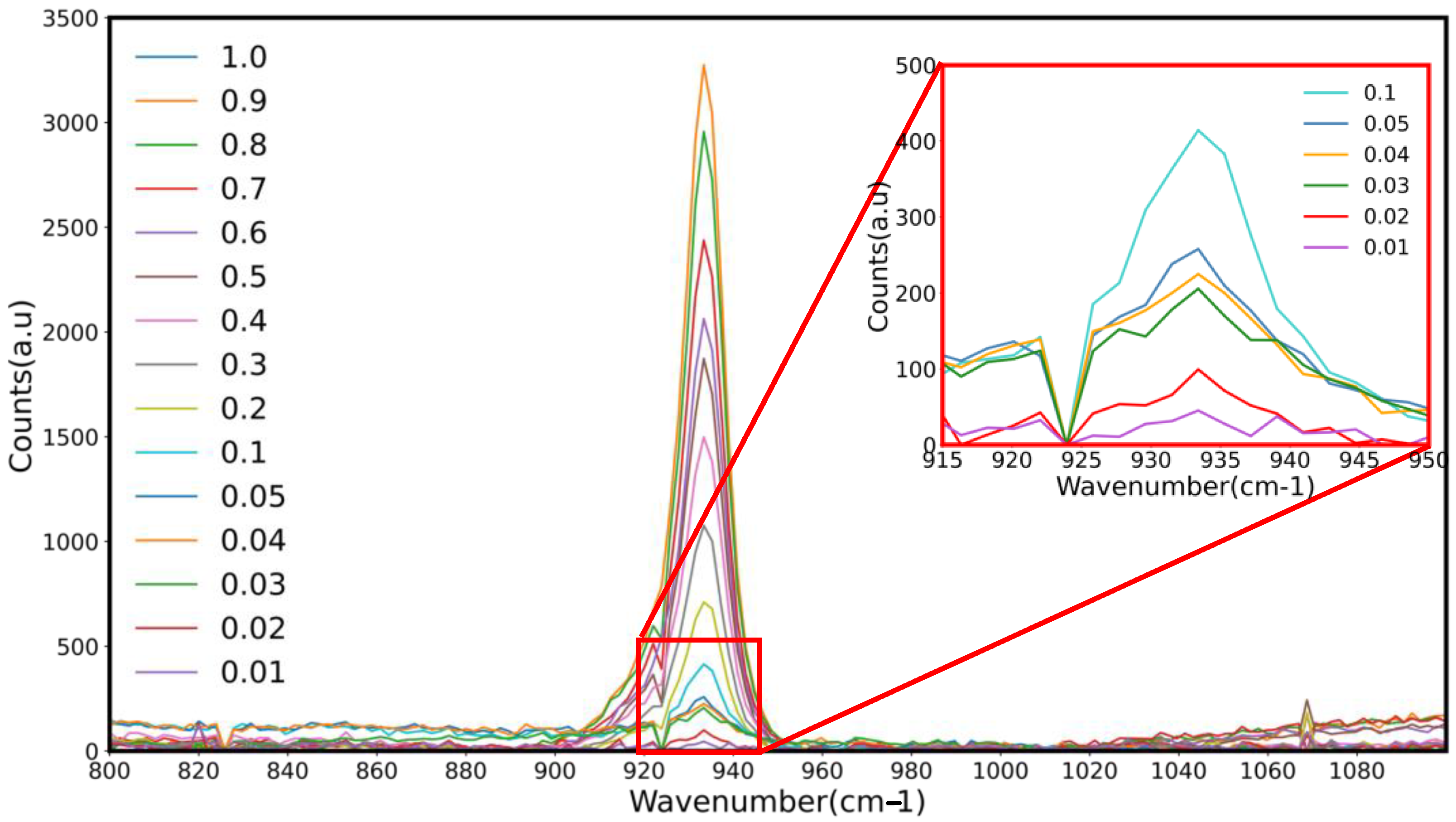
Figure 2 and
Figure 3 show Raman spectra with over-lapping peaks showing differences in peak heights associated with the varying percentages of sodium perchlorate (0.01–1%) in Davis minimal medium and RODI, respectively, used in the creation of a standard curve. Five spectra were taken every 3 s resulting in the recorded peak. Each concentration was recorded in triplicate, therefore a total of 15 technical repeats encompass a single peak on final shown spectra. Sodium perchlorate was found to have an absorbance peak of 847.07 nm. As the concentration of each perchlorate standard decreased, so too did the corresponding peak height. This proved that the portable Raman probe system is capable of detecting as little as 0.01% sodium perchlorate within DMM and RODI solution [
57].
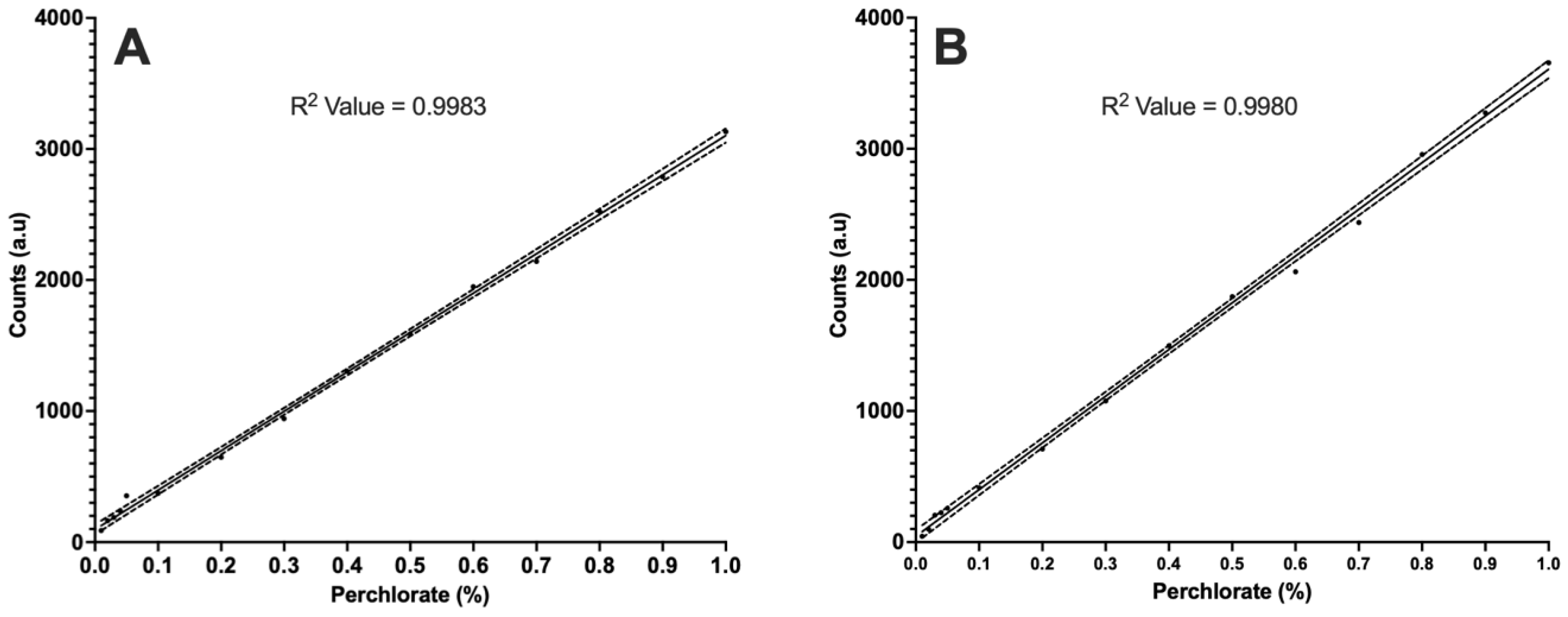
Figure 4 shows the observed linearities of (A) DMM and (B) RODI as recorded from
Figure 1 and
Figure 2, respectively. The R
2 of DMM was found to be 0.9983 and for RODI it was found to be 0.9980.
In order to detail both the metabolisation of sodium perchlorate and quantify the degree of microbial-based metabolism, Raman laser spectroscopy was used to detect/quantify the amount of sodium perchlorate present within the metabolised samples. Prior to experimentation a standard curve was created using known concentrations of sodium perchlorate and linearity was assessed to ensure validation. The laser was tested using iso-propyl alcohol before each run to ensure all fibre optic components were intact.
The test strains were standardised to an OD600 nm reading of 0.012 and grown in 0.3% sodium perchlorate dissolved in sterile Davis minimal broth that had been formulated using RODI water to ensure the absence of background interference from any possible perchlorate contamination. Another medium consisting of only sterile RODI water with a final concentration 0.3% sodium perchlorate solution was also tested.
Experimentation was performed in triplicate with the average of the Raman spectroscopy readings of the samples being extrapolated from previously established standard curve, thus allowing quantitation.
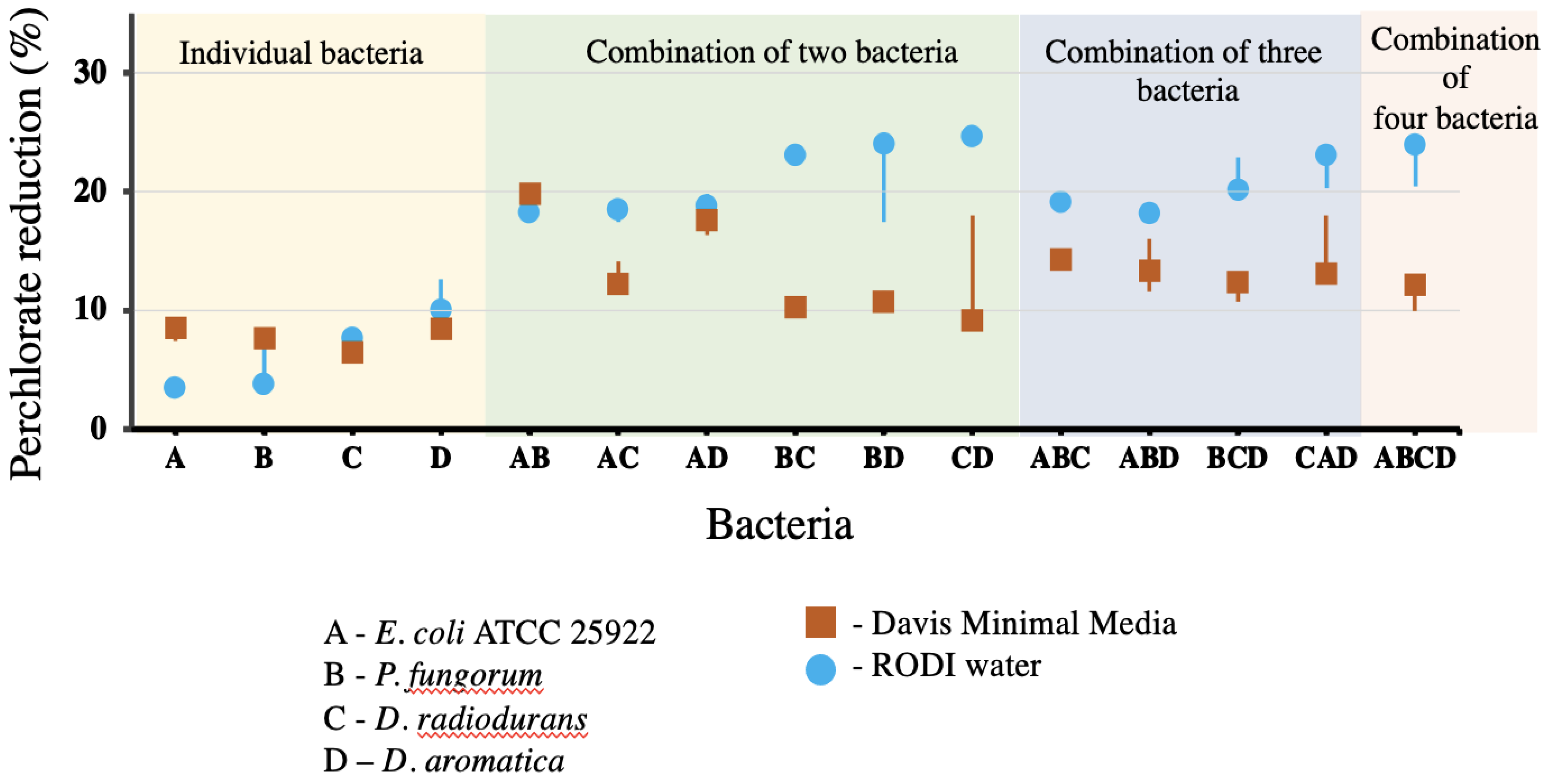
Figure 5 shows a boxplot denoting the recorded percentage of perchlorate within Davis minimal medium and RODI metabolised by the individual and combined strains. The starting sodium perchlorate concentration of each sample was 0.3%. The top three perchlorate reducers from the individual/non-combination strains within DMM were
D. aromatica (8.8%),
E. coli ATCC 25922 (8.1%) and
P. fungorum (7.7%). Meanwhile, the top three perchlorate reducers from the individual/non-combination in RODI were
D. aromatica (11.4%),
D. radiodurans (7.4%) and
P. fungorum (6.25%). The over-all average reduction of perchlorate from the individual/non-combination strains within DMM was 7.7%, and 7.2% in RODI.
The most effective perchlorate-reducing bacterial combinations within DMM were AB (E. coli ATCC 25922/P. fungorum (19.9%)), AD (E. coli ATCC 25922/D. aromatica (17.3%)) and CAD (D. radiodurans/E. coli ATCC 25922/D. aromatica (15.8%)). The overall average percentage perchlorate reduction for the combined strains within DMM was 13.3%.
The most effective perchlorate-reducing combinations within RODI were CD (D. radiodurans/D. aromatica (25%)), BC (P. fungorum/D. radiodurans (23%)), and ABCD (E. coli ATCC 25922/P. fungorum/D. radiodurans/D. aromatica (22.5%)). The overall average percentage perchlorate reduction for the combined strains within RODI was 20.7%.
4. Discussion
Restoring pristine environments and maintaining human health is one of the world’s greatest challenges. The purpose of the current study was to assess perchlorate reduction potential of our microbial counterparts both individually and in co-cultures to understand both the survival and bioremediation potential of the candidate strains when exposed to the sodium perchlorate levels found in excess to that on planet Earth and in likeness with the topography of Mars, with the dual aim of both terrestrial and extraterrestrial perchlorate reduction. Without microbial bioremediation practices, human processes on Earth and on the surface of Mars, such as the growth of crops in clean soil, would not be sustainable. The effects of Mars-like sodium perchlorate levels on both individual and co-culture strains and their ability for growth was assessed to determine how these species would behave if exposed to perchlorate from indigenous Martian soils, in a future where Mars was to be colonised by humans. This same rhetoric is applicable to the reduction of perchlorate form terrestrial soils and domestic waterways.
The novel use of Raman spectroscopy in this experiment (see experimental set-up in
Figure 1) for the detection of perchlorate within solution, pre- and post-metabolism, was found to be a robust and reliable detection method. Standard solutions were created for assessing perchlorate detection and linearity in a solution ranging from 0.01 to 1% (as seen in
Figure 2 and
Figure 3). The resulting R
2 values of each graph were close to 1.0 (see
Figure 4A,B), thus confirming the robustness in the detection technique and was deemed fit for use in the detection of microbial-based sodium perchlorate metabolism. Therefore, not only is the Raman spectroscopy sensitive enough to detect concentrations of as little as 0.01% perchlorate, it is also robust and with acceptable linearity for perchlorate quantification via extrapolation from the standard curve. As previously stated, Martian perchlorate levels have been found to range from 0.4 to 0.6% [
5], thus confirming the sensitivity of the Raman probes perchlorate detection capabilities (as low as 0.01%). While this study is able to detect Martian-specific perchlorate concentrations, if Raman was to also be employed terrestrially within environmental settings (to detect perchlorate pollution from public water systems, munitions manufacturing or agriculture [
13,
14,
15,
58,
59]), while situation dependent, its use has its limits. Within the confines of this study, it has been shown that the Raman probe is capable of detecting perchlorate concentrations as low as 100 mg/L (0.01%). Again, the degree of perchlorate contamination itself corresponds to the amount of human activity and perchlorate generation. Natural terrestrial soil perchlorate levels (within arid environments) have been documented to range from 0.03 to 0.1% (300–1000 mg) [
50], thus reaffirming the robustness of Raman detection measures terrestrially in mediums beyond the broth and RODI water examined within this study. It also reaffirms the use of Raman to detect microbial perchlorate reduction levels within the range of the standards used. However, it must be stressed that the use of Raman for perchlorate detection levels below 100 mg/L is yet to be examined and may prove to be unreliable. This highlights the need for future lab-based work investigating the limits of detection of perchlorate both within a water/broth sample and within the environment also.
With the intention to employing the candidate co-cultures to bioremediate the soils of Mars, the use of Raman laser spectroscopy was used to determine the perchlorate-reducing abilities of the microbes by means of assaying the ‘before and after’ levels of perchlorate within solution. The perchlorate reduction capabilities of the test strains were tested in both Davis minimal medium broth (DMM) and reverse-osmosis deionised water (RODI) to determine if the cultures could reduce the perchlorate in a minimal nutrient environment and in the absence of all nutrients.
Figure 5 represents the degrees of perchlorate bioremediation both individually, within co-cultures and in the presence of DMM and RODI respectively.
When the percentage bioremediation potential of both the individual and co-cultured strains are combined, as separated by media type (RODI and DMM), it was found that RODI had an average of 17.1% perchlorate reduction (with a remedial range from 3.8 to 25%), while DMM had 11.8% reduction, on average (with a remedial range of 6.2 to 19.9%).
Naturally, the presence of minimal nutrients in the minimal medium will ultimately support microbial growth significantly more than a medium devoid of any nutrients (RODI), therefore allowing the replication of more cells which in theory should allow for the reduction of more sodium perchlorate. Yet, on average, the co-cultures reduced more sodium perchlorate in RODI. It was also noted that perchlorate reduction within the individual strains across both mediums was close. When co-culturing was introduced, the range in perchlorate reduction between both medium types widened. This may be due to the lack of electron acceptor choice within the RODI samples, thus forcing the microbes to reduce the sodium perchlorate in turn. DMM, while a minimal media, still contains carbon-based nutrients such as citrate which support growth and that the bacteria may choose to utilise as a primary electron acceptor before turning to perchlorate. Conversely, this is a putative reason as to why those co-cultured in RODI (as provided with only perchlorate to utilise as a primary electron acceptor) exhibited higher levels of perchlorate reduction.
It also noted that the experimental conditions were initially conducted aerobic conditions which can hinder the perchlorate reducing potential of some strains [
26]. The creation of an anaerobic environment on Mars may be too intricate for early colonisation, while an aerobic environment is commonplace terrestrially, therefore it was deemed important to conduct the study in aerobic conditions such as what may be present within a terrestrial bioremediation setting or similar to a bioreactor or on the surface of Mars. In addition to being a preliminary study, it serves to help create a realistic scenario of the potential culture conditions found on Mars.
Interestingly, when the perchlorate reducing genes (perchlorate reductase;
pcrABCD) of
Dechloromonas were analysed in a previous study, sequence analysis of
pcrAB showed similarity to the α- and β-subunits of microbial nitrate reductase, selenate reductase, dimethyl sulfide dehydrogenase, ethylbenzene dehydrogenase and chlorite reductase, all of which are type II members of the microbial dimethyl sulfoxide (DMSO) family [
26,
60]. Furthermore, when the
pcrA gene was deleted within
D. aromatica it was found to prevent growth in both perchlorate and chlorate, but not in nitrate [
26]. These data suggest that the
pcrABCD operon plays a functional role separate to the reduction of nitrate. When subjected to phylogenetic analysis, it was found that the
pcrA gene forms its own monophyletic group in the type II DMSO enzymes and has a common ancestor with the nitrate reductase gene
narG found in
E. coli [
26]. With the absence of nitrate within the RODI and the shared affinity that both nitrate and perchlorate have for the electron acceptor, this may explain why
E. coli ATCC 25922 showed such high perchlorate reducing capabilities for a species lacking chlorite dismutase and perchlorate reductase. It is possible to speculate that there may be a link between
NarG nitrate reductase gene and perchlorate reduction. While this falls outside the scope of the current study, it warrants further investigation.
The addition of a fast-growing microbe such as
E. coli ATCC 25922, to a co-culture destined for the harsh terrain of Mars may prove to be a key benefit in ensuring active microbial action.
E. coli ATCC 25922 is known to be a strong biofilm former [
61], which may help to provide protection from environmental assault on Earth and Mars [
62] and some species are even known to grow in water making it an ideal test candidate [
63,
64]. When examining the ranking of the bioremediation rankings within RODI and DMM, it is evident that co-cultures containing fast growing strains like
E. coli are higher performing in DMM in contrast to RODI where the bioremediation strains (
D. aromatica and
P. fungorum) are more efficient at perchlorate reduction. This re-affirms the hypothesis that the addition of a fast-growing strain could prove highly beneficial to future bioremediation work.
5. Conclusions
In summary, sodium perchlorate, despite its toxic nature, poses a challenge for both terrestrial life and Martian colonists alike. While found naturally within the soils of Mars and throughout the deserts of Earth, humankinds’ abiotic activities have led to an increased prevalence of perchlorate within terrestrial ecosystems and even municipal water supplies. As a result, there is a need for innovative new technologies geared towards the monitoring and reduction of perchlorate which can help support terrestrial planetary health and the subsequent health of Martian colonists. As such, the aim of this study was to test the perchlorate reducing capabilities effects of four candidate bioremediation strains, both individually and in a co-culture, in varied media types by means of Raman spectroscopy. Overall, it was found that perchlorate reduction was higher in a media devoid of any nutrients; RODI, than its minimal media counterpart, postulated to be due to the lack of choice in available electron acceptors within the RODI. When the bioremediation potential of the strains was tested, the top three performing co-cultures within DMM were AB (E. coli/P. fungorum), AD (E. coli/D. aromatica), and CAD (D. radiodurans/E. coli/D. aromatica), while the top three performing combinations within RODI were CD(D. radiodurans/D. aromatica), BC (P. fungorum/D. radiodurans) and ABCD (E. coli ATCC 25922/P. fungorum/D. radiodurans/D. aromatica). Through this experimentation it was found that E. coli 25922 was a strong perchlorate reducer and this was postulated to be due to the shared ancestry between the nitrate reductase gene and the perchlorate reducing genes, which share the same electron acceptor system.
While employing Raman as a means of detecting and quantifying microbial perchlorate reduction shows promise, this study underlines the need to further explore the limits of its use for the detection of minute concentrations of perchlorate within the lab and within the field for environmental monitoring purposes. Consequently, when assessing the bioremediation potential and purpose suitability of the test subjects and whether the degree of perchlorate reduction could be quantified using Raman, each of the strains A, B, C and D, showed promise. It can be concluded, that the use of Raman detection methods coupled with these bioremediation strains demonstrates potential to be used to benefit both Earth and Mars to assist in the restoration of pristine environments (with benefits for natural ecosystems and for agriculture) and potential future colonisation processes.

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




