
Comparing Contemporary Evangelical Models Regarding Human Origins

Discovery Institute, Seattle, WA 98104, USA
Religions 2023, 14(6), 748; https://doi.org/10.3390/rel14060748
Submission received: 12 May 2023
/
Revised: 1 June 2023
/
Accepted: 1 June 2023
/
Published: 5 June 2023
(This article belongs to the Special Issue Exploring Science from a Biblical Perspective)
Abstract
:Multiple viewpoints exist among Protestant Evangelical Christians regarding human origins, with each offering different answers to questions regarding the existence of Adam and Eve and their relationship to humanity, common human–ape ancestry, evolution and intelligent design, humanity’s relationship to other members of the genus Homo (e.g., Neanderthals and Denisovans), and the timing of human origins. This article will review eight models for human origins which have recently received attention: (1) the Classical Theistic Evolution/Evolutionary Creationism model, (2) the Homo divinus model, (3) the Genealogical Adam and Eve model, (4) the Homo heidelbergensis model, (5) the Unique Origins Design model, (6) the Classical Old Earth Creationist model, (7) the Classical Young Earth Creationist model, and (8) an Old Earth/Recent Humans Hybrid model. Key features of each model will be described, and critical responses will be discussed in light of agreement or disagreement with traditional Judeo-Christian theological views and the scientific evidence. Most of these models maintain that science does not force one to abandon belief in core tenets of a traditional Adam and Eve, though they resolve the relevant scientific and theological questions in different ways and with varying degrees of success.
1. Introduction
Traditionally, Christians have believed that humanity began when God created Adam and Eve, who are typically considered to be the initial sole progenitors of all living humans. This view is important to many Christians who view Adam and Eve’s fall into sin as theologically crucial, because that sinful nature then spread into all of their descendants, spurring the later coming of Christ to die on the cross and atone for the sins of humanity (Reeves 2009; Grudem 2017a, 2017b; Waters 2017). However, this view is not universally accepted among Christians, some of whom do not see Adam and Eve as necessary to central Christian theological doctrines.
While the debate among Christians over human origins and Adam and Eve is nothing new, it reached a renewed intensity in 2011 when Christianity Today (CT) published a cover story titled, “The Search for the Historical Adam.” The article did not insist on a traditional view of Adam and Eve, and highlighted evangelical thinkers who accept modern evolutionary biology and are skeptical that Adam and Eve existed. For example, CT highlighted the views of Francis Collins, the evangelical Christian geneticist who headed the Human Genome Project and wrote the 2006 bestselling book The Language of God, stating that he “reported scientific indications that anatomically modern humans…originated with a population that numbered something like 10,000, not two individuals” (Ostling 2011).
Four years before that article, Collins had founded the BioLogos Foundation to promote theistic evolution, or evolutionary creationism (TE/EC), aiming to show that an evolutionary scientific viewpoint is generally correct and fully compatible with Christianity (Luskin 2014). The CT article featured the viewpoints of BioLogos-affiliated scientists. “Dennis R. Venema, the BioLogos senior fellow for science and the biology chairman at Trinity Western University, is among the BioLogos writers who are not only advocating theistic evolution but also rethinking Adam,” explained the story. Venema and then-BioLogos president Darrel Falk co-wrote an informational document on Adam and Eve, which CT quoted: “The BioLogos paper by Venema and Falk declares it more flatly: The human population, they say, ‘was definitely never as small as two…. Our species diverged as a population. The data are absolutely clear on that.’” Other evangelical scientists were quoted as saying things such as Adam and Eve “do not fit the evidence,” or although there was “wiggle room in the past” to believe in Adam and Eve, “human genome sequencing took that wiggle room away” (Ostling 2011). Some years later Venema appeared to concede that the issue was more complex and that there were in fact ways to reconcile the latest genomic evidence with a literal Adam and Eve (Venema 2018; see also Section 2.1.1). However, that is to get ahead of the story. Here, suffice to say that in the years following the CT article, multiple novel viewpoints on human origins were articulated, refined, and promoted within the Christian community—with most aiming to reconcile the latest scientific findings with some form of belief in Adam and Eve.
2. Results
If there is one space where standard evolutionary models come into sharpest conflict with traditional Christian beliefs, it is within human origins (Collins 2011). Traditionally, Christians have believed that humanity began when God created Adam and Eve. All humanity is thus descended from these two individuals—and they are usually described as not only our universal progenitors (i.e., progenitors of all living humans), but as our initial and sole progenitors (Collins 2011, 2013; Barrett and Caneday 2013; Barrick 2013b; Walton 2013). Their initial sole progenitorship of humanity is important to many Christians who view Adam’s sin as spreading through the entire human race (Reeves 2009; Collins 2011). Moreover, historically Adam and Eve were not said to have evolved or descended from apelike ancestors or more-primitive non-human species, but were believed to have been specially and miraculously created by God de novo in their present human form (Ostling 2011; Walton 2013; Barrick 2013b). Old Testament scholar John Walton explains the traditional understanding of human creation:
Formed from dust. The most obvious statement about Adam—and the one most important to this discussion—is the statement that God formed (yaṣar) him from the dust (‘apar) of the earth. Is this intended to be a statement about the material origins of the first human being? Traditionally, it has been common to think about this statement as describing a material process of special creation characterized by discontinuity with any previously existing creature.
It was also traditionally believed that Adam and Eve lived just a few thousand years ago (Lamoureux 2009, pp. 77–78; Barrett and Caneday 2013; Barrick 2013a, 2013b). As Barrett and Caneday (2013) explain, major Protestant authorities have historically held this traditional view:
The Christian view of creation that Darwinism rivaled was not monolithic, even if the popular belief among Christians was that God created the cosmos approximately 4000 years before the coming of the Christ. This belief preceded the publication of The Annals of the World by James Ussher, Archbishop of Armagh, who attempted to identify the time of creation’s beginning with precision. For example, both leading church Reformers Martin Luther and John Calvin believed that creation was not yet 6000 years old and that God created all things within six twenty-four-hour days. Calvin is thoroughly conversant with Augustine, agreeing with him at several points, but also rejecting his belief that God created all things instantaneously.
Likewise, biologist and theologian Denis Lamoureux states that “Generations of Christians have firmly believed that the creation of Adam and Eve in Gen 2 is an elaboration of the brief account of human origins on the sixth creation day in Gen 1. This traditional literal interpretation asserts that human history begins with the events in the garden of Eden” (Lamoureux 2009, pp. 77–79).
In contrast to the traditional view, the standard evolutionary viewpoint holds that modern humans evolved from a population of thousands of individuals which traces back to the most recent common ancestor (MRCA) we share with living apes, and the human population was never reduced to a mere two individuals. Moreover, under the standard evolutionary viewpoint, the ancestral lineage of modern humans leading back to the MRCA with apes is one of descent with modification where purely natural evolutionary mechanisms—e.g., random mutation, natural selection, genetic drift, etc.—were responsible for human evolution. Other models have been developed which propose different variations and mixtures of traditional theological views and mainstream scientific viewpoints regarding human origins.
2.1. Classical Theistic Evolution/Evolutionary Creationist Model
Proponents of theistic evolution/evolutionary creationism (TE/EC) seek to reconcile Christianity with the standard evolutionary model of human origins. As such their view is essentially indistinguishable from the standard evolutionary model, described above, although they believe that somehow God oversaw the evolutionary process. They generally believe that “scientific evidence is irrelevant to the Bible,” because the Bible “is simply not a science book” (Davis 2012), and are frequently willing to reshape theology when necessary to fit within an evolutionary context.
TE/EC proponents frequently accept that humans “are utterly unique and distinguished from the rest of creation because only they bear the Image of God and have fallen into sin” (Lamoureux 2009, p. 28). However, they typically reject the idea that humanity is descended from merely two individuals such as Adam and Eve, normally doubting their existence as historical individuals. Biologist and theologian Denis Lamoureux, a self-described “evolutionary creationist,” notes that evolutionary creationists “do not accept the existence of Adam and Eve” (Lamoureux 2009, p. 71), and summarizes this perspective on human origins:
The greatest problem with evolutionary creation is that it does not embrace the traditional literal interpretation of the opening chapters of the Bible. Church history shows that nearly all Christians have understood Gen 1–3 to be a basic record of actual events in the past. Specifically, most have believed that Gen 2 reveals that human history began with Adam and Eve.
Instead, TE/ECs opt for the view that modern humans are descended from a hominid population that never dipped below a size of thousands of individuals. Under this view, all living humans are also descended from a common ancestor we share with apes. Humans are said to have historical evolutionary relationships with all other living species, including relatively recent humanlike hominids such as the Neanderthals or Denisovans, and also with earlier apelike hominids such as the australopithecines. Some of these species may even have been our direct ancestors (e.g., the australopithecines).
Moreover, because humanity evolved via natural mechanisms, a full-throated TE/EC view holds there was no miraculous or special creative activity from God involved in the origin of humanity. Even if God somehow guided this process, as Francis Collins explained, under TE/EC there would be no observable evidence for design in the biological history of human origins, because “from our perspective, limited as it is by the tyranny of linear time, this would appear a random and undirected process” (Collins 2006, p. 205). Although TE/EC proponents believe God oversaw the process of human origins, by accepting a standard evolutionary model they effectively believe our biological origins were the result of strictly natural and mechanistic causes, and reject the idea that intelligent design is scientifically detected in nature (Luskin 2014). One possible exception to natural causation could be God creating the human soul (Collins 2006, p. 207), although some TE/EC proponents maintain that human psychology—including our religious impulses—arose via natural evolutionary mechanisms (Rossano 2010; Luskin 2011).
TE/ECs commonly cite certain specific lines of scientific evidence in favor of their views. This often includes fossil evidence for human evolution from apelike ancestors (Falk 2017), such as claims that australopithecine hominids had “transitional features” between apelike creatures and humans, including jaws, teeth, and a combination of “an ape-like rib cage and human-like hip bones” (Lamoureux 2009, pp. 125, 131). Genetic evidence is also cited to claim that “chimpanzees are our closest ‘cousins’” with “99%” similar DNA (Lamoureux 2009, p. 126). Another common genetic argument for human–ape common ancestry cites shared “junk” DNA between humans and apes (especially pseudogenes or endogenous retroviruses), and general genetic similarities between humans and apes (Falk 2004; Collins 2006; Alexander 2008; Lamoureux 2009; Venema and McKnight 2017). Under a classical TE/EC view, it is unlikely that God would place non-functional DNA into the same locations of the genomes of separate species, and a more likely explanation is that we inherited this “broken” or “junk” DNA from a common ancestor. For example, in The Language of God, Francis Collins maintained that some “45 percent of the human genome” consists of “genetic flotsam and jetsam,” making “the conclusion of a common ancestor for humans and mice…virtually inescapable” (Collins 2006, pp. 134–37).
The 2011 CT article highlighted a new TE/EC argument—that modern day human genetic diversity is so great that it could not be explained by humans descending from a mere initial pair of two individuals. Many thousands of initial ancestors to modern humans would be necessary to account for the genetic diversity observed in humans today. TE/ECs have placed great confidence in this argument. In their book Adam and the Genome, Venema and McKnight (2017), wrote that the existence of Adam and Eve is as decisively refuted by the data as the geocentric model of the solar system:
[W]e can be confident that finding evidence that we were created independently of other animals or that we descend from only two people just isn’t going to happen. Some ideas in science are so well supported that it is highly unlikely new evidence will substantially modify them, and these are among them. The sun is at the center of our solar system, humans evolved, and we evolved as a population.
This population genetics argument against the existence of Adam and Eve has become one of the most forceful arguments advanced for human evolution.
2.1.1. Responses
An obvious benefit of this model is that it is fully compatible with mainstream evolutionary science. However, in denying the existence of a historical Adam and Eve, it explicitly rejects major traditional theological beliefs about human origins, which will not be acceptable to many seeking to preserve those beliefs (Grudem 2017a, 2017b). Before accepting the TE/EC model, those who value these major traditional theological beliefs may wish to first consider the extent to which scientific evidence against the existence of Adam and Eve and in favor of human evolution is as open and shut as is being claimed.1
Fossil evidence: First, the fossil record shows a distinct break between the apelike australopithecines, which are supposedly directly ancestral to our genus Homo, and the first humanlike members of the genus Homo (Luskin 2022). Such evidence has led to observations from mainstream evolutionists conceding that there is a “large, unbridged gap” between humanlike members of Homo and the australopithecines (Mayr 2004, p. 198), which required a “genetic revolution” since “no australopithecine species is obviously transitional” (Hawks et al. 2000, p. 4), and implies a “big bang” model of human origins (University of Michigan News Service 2000). While evolutionary paleoanthropologists generally believe that “the transition from Australopithecus to Homo was undoubtedly one of the most critical in its magnitude and consequences,” they admit that “many details of this transition are obscure because of the paucity of the fossil and archaeological records” (Lieberman et al. 2009, p. 1). This lack of fossil evidence for the evolution of the humanlike body plan in the fossil record weakens the necessity of adopting standard evolutionary explanations of human origins.
Genetic evidence: Genomic comparisons between humans and chimpanzees are becoming more sophisticated, with recent proposals proposing lower estimates of human–chimp genetic similarity estimates to between 84% and 96% (Buggs 2018c; Seaman and Buggs 2020). It is also unclear how any percent similarity between human and chimp DNA yields an argument that requires common ancestry (Luskin 2022).
Junk DNA genetic arguments for common human–ape ancestry have also come under significant critique in recent years due to the discovery of mass-functionality for non-coding or “junk” DNA in the human genome. A major 2012 Nature paper by the ENCODE consortium reported “biochemical functions for 80%” of the human genome (ENCODE Project Consortium 2012, p. 57). Lead ENCODE scientists predicted that with further research, “80 percent will go to 100” since “almost every nucleotide is associated with a function.” (Yong 2012). In the wake of this research, the journal Science published an article titled “ENCODE Project Writes Eulogy for Junk DNA” which stated that these findings “sound the death knell for the idea that our DNA is mostly littered with useless bases” (Pennisi 2012, p. 1159). Evidence of functions for non-coding DNA has continued to mount at a high pace. A 2021 article in Nature reported that over 130,000 specific “genomic elements, previously called junk DNA” have seen specific functions identified (Gates et al. 2021, p. 215), followed by a paper in Genome Biology and Evolution which concluded, “The days of ‘junk DNA’ are over” (Stitz et al. 2021, p. 11). There is still much we do not understand about the genome and there are many specific genetic elements for which no function has yet been discovered. Nonetheless, this evidence suggests a strong trendline in the research literature away from non-functionality for “junk” DNA.
One frequently mentioned example of junk DNA is pseudogenes, which TE/EC advocates commonly cite as “the mutated remains of once-functional genes” (Venema and Falk 2010) that show our common ancestry with apes. Yet, a 2012 paper found pseudogene function is “widespread,” and since “the study of functional pseudogenes is just at the beginning” it predicted “more and more functional pseudogenes will be discovered as novel biological technologies are developed” (Wen et al. 2012, p. 31). Indeed, the literature is now replete with papers reporting functions for pseudogenes (Hirotsune et al. 2003; Pain et al. 2005; Zhang et al. 2006; Tam et al. 2008; Piehler et al. 2008; Poliseno et al. 2010; Muro et al. 2011; Rapicavoli et al. 2013; Ji et al. 2015; Hayashi et al. 2015; Prieto-Godino et al. 2016; Suzuki et al. 2018; Fiddes et al. 2018; Habib et al. 2019), including producing functional RNA transcripts or performing functions without producing any RNA (Poliseno 2012). A 2012 paper notes that although “pseudogenes have long been dismissed as junk DNA,” recent advances have established that “the DNA of a pseudogene, the RNA transcribed from a pseudogene, or the protein translated from a pseudogene can have multiple, diverse functions,” concluding that “pseudogenes have emerged as a previously unappreciated class of sophisticated modulators of gene expression” (Poliseno 2012, pp. 1, 10). Indeed, many pseudogenes are known to produce proteins, with a study in Nature reporting “more than 200 peptides that are encoded by 140 pseudogenes” (Kim et al. 2014, p. 579).
There are good reasons to understand why DNA labeled a “pseudogene” is now turning out to have a function. Pseudogenes are often identified by comparison to some other similar gene sequences encoding a functional protein from which the pseudogene is thought to have been derived and subsequently degenerated. However, regulatory functions of pseudogenes often require them to have some sequence similarity (homology) to their protein-coding counterparts so their RNA transcripts can interact with transcripts from the protein-coding gene, thereby regulating protein production (Salmena et al. 2011). Therefore, the reason our genomes contain sequences that resemble protein-coding-genes but do not produce proteins is not because they are discarded evolutionary “pseudogenes,” but because they are designed that way as important genomic regulatory and control elements.
Many papers have criticized the assumption that pseudogenes are functionless junk DNA and now suggest abandoning the term (Pink et al. 2011; Poliseno 2012; Wen et al. 2012; Kovalenko and Patrushev 2018; Troskie et al. 2021). A 2020 paper in Nature Reviews Genetics observes, “Where pseudogenes have been studied directly they are often found to have quantifiable biological roles,” and warns that “the dominant limitation in advancing the investigation of pseudogenes now lies in the trappings of the prevailing mindset that pseudogenic regions are intrinsically non-functional.” It cautions that pseudogene function is “prematurely dismissed” due to “dogma” (Cheetham et al. 2020).
There are prominent examples of prematurely dismissing pseudogene function, only to be proven wrong later. During the 2005 Kitzmiller v. Dover trial, biologist Kenneth Miller testified that our beta-globin pseudogene is “broken” because it has “molecular errors that render the gene non-functional,” indicating humans share a common ancestor with apes (Miller 2005, p. 79). Two years later, leading evolution advocate Eugenie Scott claimed this pseudogene “isn’t going to do diddly. It’s just going to sit there” and “not do a thing” (Scott 2007). However, a 2013 study reported that this precise pseudogene is functional (Moleirinho et al. 2013), and a 2021 study found it is “essential” and has “indispensability” for human red blood cell formation (Ma et al. 2021, pp. 478, 490).
Endogenous retroviruses (ERVs) are another class of “junk” DNA commonly cited by TE/ECs in favor of human/ape common ancestry. The structure of the arguments here is similar to those used for pseudogenes: ERVs also show widespread evidence of function, particularly gene regulatory roles (Conley et al. 2008; Jacques et al. 2013; Chuong 2018; Robson and Mundlos 2019; Jönsson et al. 2021; Sakashita et al. 2023), and their similarity to viral DNA may exist for functional reasons, as they are involved in immune-functions to repel viral infections (Badarinarayan and Sauter 2021).
Again, it is true that there is still much we do not know about junk DNA and there are many specific genetic elements (including pseudogenes and ERVs) for which specific functions have not yet been discovered. However, recent trends in research show that far more functionality is being discovered than was anticipated, leading to the possibility of mass functionality for junk DNA. As a 2023 academic book on RNA states:
While the story is still unfolding, we conclude that the genomes of humans and other complex organisms are not full of junk but rather are highly compact information suites that are largely devoted to the specification of regulatory RNAs. These RNAs drive the trajectories of differentiation and development, underpin brain function and convey transgenerational memory of experience, much of it contrary to long-held conceptions of genetic programming and the dogmas of evolutionary theory.
If noncoding/junk DNA is in fact functional, then genetic similarities could be the result of common design due to the need to meet similar functional requirements. Even Francis Collins has acknowledged that shared genetic similarity “alone does not, of course, prove a common ancestor” because “such similarities could simply demonstrate that God used successful design principles over and over again” (Collins 2006, p. 134).
Population genetics: Third, arguments against Adam and Eve based upon human genetic diversity and population genetics seem to have been undermined by subsequent modeling analyses. One of the first scientific responses to this population-genetics argument against Adam and Eve was published in 2012, when biologist Ann Gauger, a senior fellow with the Discovery Institute (a pro-intelligent-design think tank, where this author also works), reported that genetic diversity in HLA genes—some of the most diverse genes in the human genome—could still be explained if we originated from an initial couple (Gauger et al. 2012, p. 120).
After the publication of Adam and the Genome in 2017, biologist Dennis Venema was engaged on the BioLogos discussion forum by Richard Buggs, a geneticist at Queen Mary University London. During the discussion Venema acknowledged that various papers he had cited as having refuted the existence of Adam and Eve had not actually addressed the question of whether humanity descended from an initial couple (Buggs 2017). From this discussion emerged a consensus among various Christian biologists that if an initial pair of humans lived far enough in the past, then modern human genetic diversity could in fact be accounted for by natural biological processes. Eventually, Buggs stated to Venema: “You would do your readers a service if you wrote a blog to tell them now, as far as you are able, that present day genomic diversity in humans does not preclude a bottleneck in the human lineage between approx 700 K and 7myr ago. I think you owe this to them” (Buggs 2018b). (The bottleneck he refers to is the human population being reduced to two individuals—effectively the same as humanity descending from Adam and Eve.) Venema then publicly acknowledged the veracity of Buggs’s critique, replying: “I’ve already agreed with this….You’re welcome to publicize it” (Venema 2018). Buggs later summarized this conversation on his blog with Nature Ecology and Evolution, reporting that the question was no longer whether a historical Adam and Eve could have existed, but rather at what point in time they lived (Buggs 2018a).
Gauger along with Ola Hössjer, a professor of mathematics at the University of Stockholm, had already begun a project to address how deep into the past an initial couple had to live in order to account for modern human genetic diversity. They published a series of papers developing and testing a population genetics model which allows for an initial pair of humans to be given “designed variants” of genes representing “primordial diversity” built into the initial genomes of Adam and Eve. Under their model, natural biological processes then govern the subsequent genetic history of the human race. They showed that modern-day human genetic diversity can be explained by a single pair of ancestors—e.g., what one might call Adam and Eve—provided that they lived at least 500,000 years ago (Hössjer et al. 2016a, 2016b; Hössjer and Gauger 2019). S. Joshua Swamidass, a Christian scholar and professor of computational biology at Washington University in St. Louis, performed an analysis that yielded a similar result, finding that Adam and Eve could have lived 495,000 years ago as our sole genetic progenitors (Swamidass 2017).
There is another population genetics argument relevant to human origins—but this one is posed as a mathematical challenge to unguided evolutionary models. The MRCA of humans and chimpanzees is said to have lived approximately 4 to 6 million years ago (Wood and Harrison 2011). Though estimates vary, a reasonable accounting proposes that the genetic differences between humans and chimps amount to some “35 million base-pair changes, 5 million indels [sequences of multiple nucleotide bases] in each species, and 689 extra genes in humans” (Cohen 2007). Yet, a population genetics study in the journal Genetics found that if just two specific mutations were required to provide some evolutionary advantage in the line that led to humans, then “this type of change would take >100 million years,” which was determined to be “very unlikely to occur on a reasonable timescale” (Durrett and Schmidt 2008). This “waiting times” problem (Hössjer et al. 2021) suggests there may be far too little time available from the fossil record for standard unguided evolutionary mechanisms to generate observed genetic, morphological, and behavioral differences between humans and chimps. Some have proposed intelligent design as a possible explanation for the rapid appearance of biological information necessary to overcome the waiting times problem and generate these complex traits (Thorvaldsen and Hössjer 2020).
2.2. Homo Divinus Model of Denis Alexander
The term “Homo divinus” was coined by theologian John Stott, who viewed Adam as a possible descendant of Homo erectus that was chosen by God to be the “first man to whom may be given the specific biblical designation ‘made in the image of God’” (Stott [1972] 1999, pp. 55–56). This model built upon the ideas of Old Testament scholar Derek Kidner, who proposed that Adam was given “federal headship” over humanity, meaning that Adam’s original sin spread to both his “offspring” and his “contemporaries” (Kidner 1967, p. 29). Under this view, therefore, Adam may not be genetically or genealogically ancestral to all humans, but his sin impacted the whole of humanity, whether descended from Adam or not. This view is said to be consistent with “covenant theology,” where Jesus Christ saves all of humanity even if he is not their biological ancestor, but because he is their “federal head” (Kidner 1967, p. 30).
In more recent discourse, TE/EC proponent and biologist Denis Alexander has also promoted the Homo divinus model as a possible way to reconcile evolution with some form of a historical Adam and Eve. Alexander’s model essentially adopts the standard evolutionary view of human origins, but then proposes that at some point in the Neolithic period, God “chose a couple of Neolithic farmers in the Near East, or maybe a community of farmers, to whom he chose to reveal himself in a special way.” This “marked the time at which God chose to reveal himself and his purposes for humankind for the first time” (Alexander 2008, pp. 236–37).
2.2.1. Responses
This model fully adopts an evolutionary perspective on the origin of humans, and proposes that Adam and Eve were real people chosen by God for a special purpose who lived just a few thousand years ago. However, this is where its similarity to traditional theological beliefs ends. The “Adam and Eve” in this model are natural-born descendants from earlier hominids that evolved through standard evolutionary mechanisms, and they are in no way proposed to be specially created by God nor are they said to be the progenitors of all humans.
In fact, under this model Adam and Eve have no necessary genealogical or ancestral relationship to any other humans and play essentially no role in the physical or biological origins of humanity. Rather, their role is strictly spiritual: it is a model “about spiritual life and revealed commands and responsibilities, not about genetics” (Alexander 2008, p. 238). Thus, although the Homo divinus model allows for a recent Adam and Eve, this version of Adam and Eve does not seem genealogically connected to later humans in a manner that fulfills traditional biblical views of human descent.
This model also raises theological concerns. In proposing an Adam and Eve that are in no way biologically connected to the rest of the human race, some may feel this model does not satisfy Old Testament doctrines about Adam and Eve somehow transferring the “image of God” to the rest of humanity, nor New Testament doctrines about Adam’s sin and death somehow spreading through all humanity (Donald 2009, p. 21; Reeves 2009, p. 48; Collins 2011, p. 127; Waters 2017).
2.3. Genealogical Adam and Eve Model of S. Joshua Swamidass
Like the TE/EC model, the Genealogical Adam and Eve (GAE) framework fully adopts a standard evolutionary model of human origins, with one important exception: the special creation of Adam and Eve. In the GAE view, modern humans are descended from a population composed of tens of thousands of hominids who arose via standard evolutionary mechanisms, plus two individuals who were miraculously created. “The genealogical hypothesis, with details filled this way, is entirely consistent with the findings of evolutionary science,” writes S. Joshua Swamidass, who developed the model. “The DNA of our ancestors, their genetics, would still arise from a population, not a single couple. We would all still share common ancestry with the great apes” (Swamidass 2019, p. 10).
Thus, Swamidass evokes standard evolutionary mechanisms, but allows an important exception to them: he proposes that Adam and Eve could have been “de novo created. God creates Adam and Eve by a direct act, de novo from dust and a rib (or Adam’s side)” (Swamidass 2019, p. 25). Swamidass then immediately hedges on this point, stating that de novo creation is possible but “is not required” under his model (Swamidass 2019, p. 26). He further proposes that “Adam and Eve’s lineage eventually interbreed with people outside the Garden” (Swamidass 2019, p. 26). However, he makes it clear that the people “outside the Garden”—from whom all humans are descended—evolved via standard evolutionary mechanisms:
No additional miracles allowed. No appeals to divine action are permitted to explain the data or increase confidence in the hypothesis. Yes, one direct act of God is included in the hypothesis itself, but the evidential evaluation of the hypothesis cannot infer or rely upon divine action in any way.
He continues:
The two findings of evolutionary science. The people outside the Garden would share common descent with the great apes, and the size of their population would never dip down to a single couple.
Apart from the potential de novo creation of Adam and Eve, Swamidass’s model of human origins is thus equivalent to the standard evolutionary model and involves no other miraculous or non-natural events.2 Under this view, therefore, Adam and Eve could have been specially created and eventually became the genealogical ancestors of all living humans today. Humans are also descended from thousands of other hominid ancestors who evolved via standard evolutionary mechanisms from apelike ancestors.
2.3.1. Responses
The GAE model aims to retain space for both an orthodox view of the special creation of Adam and Eve, and possibly also their recent origin, alongside a standard evolutionary view of human origins. Swamidass even states that the GAE hypothesis supports a “traditional” version of Adam and Eve (Swamidass 2019, p. 6). However, under the GAE model, Adam and Eve are not the sole progenitors of humanity—far from it. Under GAE, the ancestral contributions of Adam and Eve to humanity (in terms of both genealogy and genetics) are likely dwarfed by the ancestral contributions of a much larger population of fully evolved hominids.
The GAE model also raises profound questions about universal human equality. Swamidass splits humanity into two groups. First, there are “textual” humans who are directly descended from Adam and Eve (as well as other hominids), are made in the image of God, inherited Adam’s sin, and are those for whom Christ died. These are the people who are referenced in Scripture. Then, there’s a second category of “biological humans” who evolved from apelike ancestors, whom Scripture never references, but who make up the great bulk of our ancient ancestral stock and perhaps even represent a large proportion of humanity throughout much of historical human history. Swamidass is unclear about whether the non-textual people had “human worth and dignity,” were made in the image of God, sinned, or had “need for a Savior” (Swamidass 2019, pp. 149–50). However, his model leaves open the possibility that they possessed none of these things, thereby potentially undermining the Judeo-Christian foundation for universal human equality, and the inherent dignity and worth of all humans.
Within Swamidass’s model, textual humans included “everyone alive across the globe by, at latest AD 1” (Swamidass 2019, p. 134), such that when Christ died all living humans were descended from Adam, inherited his sinful nature, and needed salvation. Prior to this point in human history, however, there may have lived many non-textual humans—people whom Scripture never mentions, who did not necessarily inherit Adam’s original sin, and for whom Christ did not necessarily die. Evolutionary biologist and Christian apologist Jonathan McLatchie is troubled by this proposal:
Swamidass’ view would seem to suggest logically that those individuals who were biological (but not textual) humans are qualitatively indistinct from other animals. However, in that case it makes no sense to call their deeds evil, or to postulate that they had a sense of right and wrong. Moreover, if they, as Swamidass suggests, “do wrong at times”, then does this not suggest that Adam’s fall is but one of many falls that have occurred in human history? The theological ramifications that accompany this scenario are too severe for me to entertain Swamidass’ proposal.
Similarly, theologian and medical doctor Hans Madueme is concerned about the polygenetic implications of Swamidass’s model:
Swamidass zealously distances his position from earlier racist polygenetic theories. According to those accounts, racial groups that (allegedly) didn’t descend from Adam weren’t full human beings. I agree that his position is a vast improvement. However, the genealogical hypothesis remains polygenetic, at least to some degree. Swamidass defines all those outside the garden as biologically but not textually human.
This move, however, raises a host of questions: for example, in what sense are non-Adamic biological humans fully human? If these biological humans have a different origin from Adam and Eve, do they participate in original sin and salvation? Did Christ live and die for them, and were they able to experience justification by faith? And, if human beings are natural kinds—as Christians have always believed—then how is interbreeding even possible? In chapter 14, Swamidass speculates that these biological humans are made in God’s image, with minds and souls, but “they are not yet affected by Adam’s fall. They have a sense of right and wrong, written on their hearts (Rom. 2:15), but they are not morally perfect. They do wrong at times. They are subject to physical death, which prevents their wrongdoing from growing into true evil (Gen. 6:3)” (175). Although Swamidass is only speculating here, the notion of other people outside the garden, in my view, is nowhere in Scripture. In traditional Christianity, being human and being a descendant of Adam are co-extensive. As far as I can see, Swamidass’s revisionism lacks a convincing exegetical or theological basis.
Scriptural questions are also raised about this division between “textual” and “biological” humans. In Genesis 3:20, Eve is called “mother of all the living,” but a core element of Swamidass’s model is that Eve was certainly not the “mother of all the living” at the time she was created, nor did she become an ancestor of all living humans until millennia after her creation. Swamidass concedes that passages such as Acts 17:26 (“from one man, [God] made all the nations”) and Romans 5:12–18 (“all sinned” after Adam’s fall, so Christ died for “all people”) seem to “presume universal ancestry of Adam,” but dismisses them with the casual, “They do not specifically deny mixing with other lines in the distant past” (Swamidass 2019, p. 114). William Lane Craig worries that this mixing of Adam and Eve’s descendants with evolved humans (who were not necessarily created in the image of God) is tantamount to massive amounts of “bestiality, contrary to God’s will for humanity” occurring throughout human history (Craig 2021, p. 378).
Lastly, there are reasons to wonder whether this model is necessary. Its primary goal seems to be to marry the possibility of de novo creation of Adam and Eve with a standard evolutionary model. However, if the fossil and genetic evidence does not demand or support key tenets of an evolutionary model (such as common human–ape ancestry; see Section 2.1), is this marriage necessary? Moreover, Ann Gauger and Ola Hössjer’s research—as well as Swamidass’s own modeling—show that if Adam and Eve lived far enough in the past, then modern human genetic diversity is compatible with an initial pair who were our sole initial ancestors. This seemingly eliminates a major argument for invoking thousands of evolutionary ancestors outside the line of Adam and Eve, thus rendering moot a central motive for adopting the GAE model.
2.4. Homo Heidelbergensis Model of William Lane Craig
William Lane Craig is a Christian philosopher and theologian who has written extensively on adapting the Kalam cosmological argument for the existence of God to modern scientific evidence, and other arguments for cosmic design (Craig and Smith 1995; Craig 2000, 2001; Craig and Moreland 2012). In 2021, he addressed biological origins with his book In Quest of the Historical Adam, which proposes that Adam and Eve were real historical people who could have been members of Homo heidelbergensis, a hominid species that lived about 750,000 years ago (Craig 2021, p. 330). His model of human origins allows Adam and Eve to be ancestors of other members of the genus Homo including not just our species Homo sapiens, but also the Neanderthals and Denisovans, which he argues are so similar to humans genetically (including neurogenetically), morphologically, and behaviorally that they ought to be considered as bearers of the Imago Dei. He describes his model as follows:
Adam and Eve may therefore be plausibly identified as members of Homo heidelbergensis and as the founding pair at the root of all human species. Challenges to this hypothesis from population genetics fail principally because we cannot rule out on the basis of the genetic divergence exhibited by contemporary humans that our most recent common ancestors, situated more than 500 kya, are the sole genetic progenitors of the entire human race, whether past or present. The challenge of the wide geographic distribution of humanity is similarly met by situating Adam and Eve far in the past, prior to the divergence of Homo sapiens, Neanderthals, and other species, and allowing multispecies cultural evolution to proceed thereafter in response to environmental changes to produce modern human behaviors wherever their descendants are to be found.
Thus, unlike the GAE model, Craig’s model allows and even prefers that Adam and Eve were the “sole genetic progenitors” of humanity. That said, at times he seems open to limited interbreeding or admixture between Adam and Eve’s descendants and other evolved hominids, as in the GAE model—but this “admixture hypothesis” is not his preferred view and is not necessary to his approach3 (Schaffner 2021). As noted, Craig views interbreeding between Adam and Eve’s descendants and other hominids as a rare event—equivalent to “bestiality” and “contrary to God’s will for humanity” (Craig 2021, p. 378).
Craig’s model aims to be compatible with modern evolutionary science. In an interview with Christianity Today, Craig stated that he aspires to show “there is no incompatibility between contemporary evolutionary science and the affirmation of a single human pair at the headwaters of the human race, [so] we can prevent that obstacle to faith.” (Travis 2021). Likewise, a review of Craig’s model in the journal Science observes that he “takes evolution as a given” (Schaffner 2021). However, for Craig “evolution” seems to imply common ancestry but not necessarily an entirely unguided evolutionary process. He is open to the de novo creation of Adam and Eve, but sees this as creating a dilemma:
One can … postulate instead a de novo creation of Adam and Eve. But then one faces a difficult dilemma. One must explain our genetic similarity to chimps either on the basis of repetitive divine use of a similar design plan or on the basis of considerable interbreeding with nonhumans. The first has difficulty explaining broken pseudogenes that we share with chimps … The second looks as if God condones bestiality for our forebears.
Because Craig eschews the idea of massive amounts of bestiality in human history, he seeks to account for modern human genetic similarities to apes by proposing that Adam and Eve may ultimately descend from a common ancestor shared with apes. However, he argues that this does not preclude them from having been the sole progenitors of subsequent members of humanity—Adam and Eve essentially represented a bottleneck of two individuals who were derived from a population that evolved from apelike ancestors:
No such appeal to interbreeding [between humans and non-human hominids] is necessary if we envision Adam and Eve as emerging from a hominin population that shared common ancestry with chimpanzees and other great apes. Indeed, on the view proposed here, Adam and Eve could be our sole genetic progenitors, whose descendants never fell into bestial relations with nonhuman hominins or at least produced no descendants from such liaisons.
What seems to be driving Craig’s model is a conviction that Adam and Eve were real historical people who could have been our sole genetic progenitors, creating a race of humans untarnished by interbreeding with other hominids, yet sharing genetic properties with apes. His model requires a very ancient Adam and Eve in order to accommodate them being ancestral to other humanlike members of the genus Homo (again, Neanderthals or Denisovans), whom he believes were so morphologically, genetically, and behaviorally advanced that they were probably made in the image of God. Another key reason Craig requires an ancient Adam and Eve is that he desires them to be “our sole genetic progenitors.” He cites studies showing that modern human genetic diversity can be accounted for only if humanity traces to an initial pair that lived at least 500,000 years ago (Craig 2021, p. 353).
2.4.1. Responses
Craig’s model seeks to posture Adam and Eve as an initial pair of progenitors—a bottleneck of two—from whom all subsequent humanity descended, and allows humanlike members of the genus Homo such as the Neanderthals and Denisovans to be descendants of Adam. His model accomplishes this by placing Adam and Eve hundreds of thousands of years in the past, a position which also can account for modern-day human genetic diversity. However, an ancient Adam and Eve who lived hundreds of thousands of years ago could be viewed as problematic by some who would cite biblical genealogies as requiring a more recent Adam and Eve. If these genealogies are incomplete (Kitchen 1966, pp. 37–39; Henry and Dyke 2012; Keathley and Rooker 2014, pp. 169–77; Collins 2018, p. 200), however, then this problem might be avoidable.
Craig’s model is compatible with either the de novo creation of Adam and Eve or an Adam and Eve who evolved from apelike ancestors, sharing a common ancestor with apes. Craig prefers the latter position—human–ape common ancestry—because it explains human–ape genetic similarity and the presence of what he calls “broken pseudogenes” shared in the genomes of humans and apes. He believes this genetic evidence is incompatible with the de novo creation of Adam and Eve. However, if pseudogenes are in fact functional genetic elements and not merely “junk” DNA (see Section 2.1.1), then there could be functional reasons for the fact that humans and apes share them in similar locations. That is, functional genetic similarities between humans and apes would be the result of common design rather than common descent, and would not require an evolutionary origin. Thus, the genetic evidence does not necessarily refute the de novo creation of humans as Craig seems to think it does.
Another issue raised by Craig’s model is that Adam, Eve, and the entire human race are arising hundreds of thousands of years before there is definitive evidence for modern human intelligence in the archaeological record. Many researchers have recognized an “explosion” or “revolution” (Bar-Yosef 2002; Mellars 2004; Nowell 2006) of modern humanlike culture in the archaeological record showing the abrupt appearance of human creativity (White 2003, pp. 11, 231), technology, and art (Rice 2007, pp. 104, 187, 194)—even including paintings (Kelly and Thomas 2010, p. 303)—and the rapid emergence of self-awareness, group identity, and symbolic thought (Bar-Yosef 2002). One review of Paleolithic archaeology dubbed this the “Creative Explosion” (Toth and Schick 2015, p. 2459). However, this explosion of modern humanlike creativity does not appear until about 100,000 years ago—hundreds of thousands of years after Craig places the origin of Adam and Eve.
This is not to say that there is no evidence for high intelligence among earlier hominids such as the Neanderthals. Studies have found that Neanderthals had the morphological capability for human speech (Arensburg et al. 1989; D’Anastasio et al. 2013), and Neanderthal remains have reportedly been found associated with signs of culture including art, burial of their dead, and complex tools (Stringer 1992; Trinkaus and Shipman 1993; Folger and Menon 1997; Chase and Nowell 1998; Alper 2003; Wong 2003)—including musical instruments like the flute (Chase and Nowell 1998; Folger and Menon 1997). Morphological mosaics—skeletons showing a mix of human and Neanderthal traits—suggest “Neandertals and modern humans are members of the same species who interbred freely” (Trinkaus and Duarte 2003). Even among the earlier Homo erectus, there is direct and circumstantial evidence that they may have controlled fire (Wrangham and Carmody 2010; Zhong et al. 2014; Chazan 2017; Hlubik et al. 2017) and traveled by boat (Bednarik 1997; Gibbons 1998)—activities which presumably required high intelligence and even speech (Everett 2018).
However, not all experts agree with these claims and the evidence for the technological, artistic, and cultural abilities of Neanderthals, Homo erectus, and other contemporary hominids is debated (Rana and Ross 2015). Uncontested evidence of high human intelligence and cultural activity in the archaeological record does not exist until long after Craig’s proposed date for Adam and Eve.
2.5. Unique Origins Design Model of Ann Gauger and Other Intelligent Design Advocates
As noted earlier, after the 2011 CT article various intelligent design (ID) proponents collaborated on a project to ask whether human genetic diversity could be explained if we descended from an initial pair. Intelligent design is a scientific theory which holds that some features of the universe and life are best explained by an intelligent cause rather than an undirected cause such as natural selection (Meyer 2009, p. 4). The leaders of this project were design theorist and biologist Ann Gauger and mathematician Ola Hössjer who co-published three papers from 2016 to 2019 showing that modern human genetic diversity could be resolved back to just two individuals. (Hössjer et al. 2016a, 2016b; Hössjer and Gauger 2019). Their population genetics model found that if these two individuals lived at least 500,000 years ago then modern-day human genetic diversity could be accounted for with humanity arising from an initial couple. The assumption in their model is that this initial couple had “designed variants” of genes representing “primordial diversity” built into their genomes and so did not need to descend from previous hominids via natural evolutionary processes to provide the requisite genetic diversity (Hössjer and Gauger 2019). According to this model, the pair did not share common ancestry with apes and would have been the sole genetic progenitors of modern humans—the couple from whom all humanity descended.
If this couple lived hundreds of thousands of years ago, such a model would enjoy some benefits of William Lane Craig’s Homo heidelbergensis model in that it would allow other humanlike members of the genus Homo (e.g., Neanderthals or Denisovans) to also belong to a monophyletic group of hominids made in the image of God. Depending on how far back this initial couple lived (e.g., 1.5 million years), even earlier humanlike members of Homo such as Homo erectus could be viewed as “image bearers,” potentially placing Adam and Eve synchronously with the first appearance of the humanlike body plan in the fossil record.
2.5.1. Responses
The Unique Origins Design model maintains that our human lineage was designed separately from apes at least 500,000 years ago,4 and demonstrates that this can account for modern day human genetic diversity. Some evolution proponents might object that some genetic evidence remains unexplained, such as junk DNA, including what Craig calls “broken pseudogenes that we share with chimps.” As noted in Section 2.1.1, however, there are good reasons to view pseudogenes and other forms of junk DNA as functional, and it is possible to explain their presence in humans and apes with common design rather than common descent and inheritance from a common ancestor with apes.
This model requires an ancient Adam and Eve and is thereby compatible with Craig’s proposal that Adam and Eve could have belonged to the species Homo heidelbergensis or some similar contemporary member of Homo. This allows Neanderthals, Denisovans, and other humanlike members of Homo to be descended from Adam and Eve. It also allows Adam and Eve to have lived even earlier, potentially having been members of Homo erectus, thus aligning the creation of Adam and Eve with the first appearance of the humanlike body plan in the fossil record. Although there is circumstantial evidence that these earlier hominids had high intelligence, such a proposal implies that Adam and Eve and their descendants lived for hundreds of thousands of years before there is definitive evidence of humanlike intelligence and creativity in the archaeological record.
An ancient Adam and Eve could also trouble those who view biblical genealogies as requiring a more recent inception for humanity. It is important to note that the Unique Origins Design model was developed by intelligent design proponents in a series of three technical papers that were primarily asking the scientific question of whether human genetic diversity could be derived strictly from an initial pair, and those papers did not develop a full-fledged theological model of Adam and Eve. One response to concerns over an ancient Adam and Eve could be that the biblical genealogies are incomplete (Kitchen 1966, pp. 37–39; Henry and Dyke 2012; Keathley and Rooker 2014, pp. 169–77; Collins 2018, p. 200). However, even if incomplete, the ability of the genealogies to be stretched to accommodate an Adam and Eve that lived, say, 1.5 million years ago seems to have not yet been adequately investigated. Another possible response is that a much more recent Adam and Eve might still be compatible with human genetic diversity.
Gauger and Hössjer’s model essentially assumed that normal genetic/evolutionary processes occurred after the initial design of the human species, and that these processes work very slowly to allow mutations to accumulate and generate observed human genetic diversity. Hence, Adam and Eve had to live hundreds of thousands of years ago to allow for enough time for modern human genetic diversity to naturally appear. However, if additional evolutionary assumptions are questioned and the genetic history of humanity did not follow standard evolutionary rates, then human genetic diversity could be compatible with a much more recent Adam and Eve (see Section 2.6, Section 2.7 and Section 2.8). A more recent Adam and Eve might not allow other humanlike forms in the hominid record such as Neanderthals, Denisovans, or Homo heidelbergensis, to be descended from Adam and Eve. However, it could bring the timing of Adam and Eve into alignment with the explosion of modern humanlike creativity observed in the archaeological record at about 100,000 years ago.
2.6. Classical Old Earth Creationist Model of Reasons to Believe
Two of the primary expositors of a highly mature Old Earth Creationist (OEC) model of human origins are Fazale Rana and Hugh Ross, both scientists affiliated with the OEC advocacy organization Reasons to Believe (RTB) (Rana and Ross 2005; Rana and Ross 2015). OECs accept the conventional view that the earth and universe are billions of years old; however, they are generally skeptical that unguided evolutionary mechanisms are responsible for life and typically believe that God progressively and specially created various groups of organisms during the history of life. OECs also seek to find harmony between a literal reading of the book of Genesis and conventional chronology and ordering of major events in natural history. Their human origins model maintains that Adam and Eve were historical persons, the sole progenitors of humanity, and specially and miraculously created by God, separately from apes, sometime between 55,000 and 130,000 years ago (Rana and Ross 2015, p. 252; Ross 2016; Rana 2020a, p. 70; Rana 2018; Rana 2021).
An important component of the OEC model is that other members of the genus Homo such as Neanderthals or Denisovans were not descended from Adam and Eve, and were not made in the image of God, and thus they do not need to place Adam and Eve far enough back in time to be ancestral to those groups. This is important to their model, which includes a typical OEC view of a “local biblical flood,” where Adam and Eve’s descendants (apart from Noah’s family) were universally killed off in the flood because they all lived in a localized region in Mesopotamia that was impacted by this small-scale flood. If Neanderthals or other non-human hominids were descended from Adam, their widespread geographic distribution would require a much larger “global” flood to fulfill the apparent biblical requirement that all humanity was wiped out in the flood, and they do not want to postulate a global flood in their model.
Rana and Ross cite archaeological evidence as highly consistent with their model, and have argued that a creation date for Adam and Eve of roughly 70,000–80,000 years ago fits closely with the explosion of humanlike creative activity in the archaeological record around this same time (Rana and Ross 2015, p. 274). Their model is flexible, however, and finds that the “most likely” creation date of Adam and Eve is between 55,000 and 120,000 years but could be “stretched as far back as 230,000 years ago” (Ross 2016). Based upon the more recent evidence, a creation date for Adam and Eve of about 130,000 years has been proposed (Rana 2018; Rana 2021). In light of these modestly varying estimates, a rough date of ~100,000 years for the creation of Adam and Eve will be assigned to this model throughout the rest of this paper.
2.6.1. Responses
Those who desire a more recent Adam and Eve based upon biblical genealogies might find this model more palatable than Craig’s Homo heidelbergensis model or the Unique Origins Design model, which proposes they lived at least 500,000 years ago. However, this model is still unlikely to satisfy those who seek a very recent Adam and Eve based upon strictly complete biblical genealogies. OECs often maintain that the genealogies are incomplete (Henry and Dyke 2012) in attempts to avoid such problems.
On the other hand, proposing that Adam and Eve lived much less than 500,000 years ago could complicate efforts to account for modern human genetic diversity. However, as noted in Section 2.5, if the genetic evolution of humans after their creation proceeded faster than presently observed rates, these problems could be avoided. Ironically, help on this point for this OEC model may come from Young Earth Creationists who believe the earth is only a few thousand years old. Sanford et al. (2018) constructed a model which predicted that if Adam and Eve lived for hundreds of years and had dozens of children, and if Adam and Eve had genetic diversity built into their gametes, then modern human genetic diversity could be accounted for in just 200 generations—i.e., a few thousand years. This suggests that the scale of modern human genetic diversity may not be a problem for a traditional Adam and Eve specially created by God as our sole progenitors regardless of whether one adopts the Unique Origins Design model, the Old Earth Creationist model, the Young Earth Creationist model, or the Old Earth/Recent Humans Hybrid model.
OECs are skeptical on fossil and genetic grounds of evolutionary arguments that humans descend from apelike ancestors (Dykes 2020a, 2020b). Since the OEC model is a non-evolutionary model for the origin of humans, it, like the Unique Origins Design model, must account for shared “junk” DNA between humans and apes. This does not seem to be highly problematic, however, given the abundance of evidence of function for non-coding DNA (Section 2.1.1).
Another challenge to the OEC model is that many would prefer that other humanlike members of the genus Homo such as Neanderthals or Denisovans should be related to modern humans and descended from Adam and Eve. These other groups have body plans which are highly similar to modern humans and also have very high genetic similarity to modern humans. William Lane Craig notes that they share genetic traits which are thought to be required for human cognition and speech, raising the possibility that they were highly intelligent (Craig 2021, pp. 302–29). There is also genetic evidence that Homo sapiens interbred with both Neanderthals and Denisovans (Villanea and Schraiber 2018; Teixeira et al. 2021), a finding suggestive of their being part of a common human family traceable to Adam and Eve. Proponents of the OEC human origins model would reply that it is difficult to make any firm conclusions from genetics, and would cite the paucity of clear-cut archaeological evidence for high intelligence within Neanderthals, Denisovans, or any hominid that predates modern humans (Rana and Ross 2015; Dykes 2020c, 2020d). However, while the OEC model holds that Neanderthals and Denisovans were created separately from humans, it does allow for evidence of interbreeding between humans and these other groups, though this evidence was unexpected (Rana 2020b). They see their placement of Adam and Eve at ~100,000 years ago as closely aligned with the evidence for advanced human creativity appearing in archaeological records, and a major point in favor of their model.
2.7. Classical Young Earth Creationist Model
The classical Young Earth Creationist (YEC) model is perhaps the oldest model in terms of having been developed many decades ago, and it will be familiar to many readers. In general, Young Earth Creationism adopts an interpretation of the book of Genesis where God created the universe, earth, and all “kinds” of life in six 24-hour days. Under this view, on the sixth day God specially and miraculously created Adam and Eve as the sole progenitors of the human race (Morris 1976, 1977). Because all of creation is typically said to be only 6000 to 10,000 years in age, YECs would hold that Adam and Eve lived only a few thousand years ago (Jeanson 2017, pp. 191–92). Under this model, there is no physical death before the fall of Adam and Eve, and physical (and spiritual) death entered the world just a few thousand years ago when Adam and Eve committed the first sin.
After their creation, Adam and Eve (and their descendants) lived for hundreds of years and gave birth to many children, leading to rapid expansion of the human population. The YEC model holds that a few thousand years after the creation of Adam and Eve, the Earth experienced a worldwide “global flood” which killed off all humanity except for Noah and his family—a second genetic bottleneck. Other humanlike members of the genus Homo such as the Neanderthals are typically said to simply be members or subraces of the human species and were fully human.
2.7.1. Responses
Historically, many Protestants, including early reformers such as Martin Luther and John Calvin, believed in a recent creation (Barrett and Caneday 2013); however, at various points old Earth viewpoints have also enjoyed popularity among Evangelicals.5 The YEC model is not only compatible with traditional theological views of Adam and Eve, it essentially defines the traditional theological view. In addition to satisfying interpretations of the Bible which indicate a recent creation of Adam and Eve, many YEC proponents will point out that their model is distinguished because it alone accommodates the traditional view that there was no physical death prior to the creation (and subsequent fall) of Adam and Eve.
Apart from the Hybrid model (Section 2.8), any model which accepts an old Earth would require that there was physical death prior to the creation of Adam and Eve. Thus, the theological view that there was ‘no death before sin’ is a driving force for many who accept the YEC model.6 However, OECs reply that when God warned Adam and Eve not to eat from the Tree of Life, He said “in the day that you eat of it you shall surely die.” (Genesis 2:17, ESV). Yet, Adam and Eve did not “die” that very same day when they ate the fruit—but they did immediately realize they were naked, felt shame, and hid from God. Thus, OECs would argue that the warning of Genesis 2:17 speaks of “spiritual death” rather than “physical death.” (Ross 2001, p. 94; 2014, p. 110). YECs would reply that at the fall Adam and Eve experienced “the commencement of a process of physical dying” (Ham 2006, p. 99)—a plausible supposition, but the rejoinder from OECs has severely blunted the conclusion that only the YEC model can make sense of such passages. Alternatively, some proponents of an old Earth argue that Adam and Eve’s sin did in fact “cause” physical death, but the curse was applied by God retroactively to earlier periods of creation: “effects of the Fall can be retroactive as well as proactive (much as the saving effects of the Cross stretch not only forward in time but also backward, saving, for instance, the Old Testament saints)” (Dembski 2009, p. 19).
Various scientific challenges have also been posed to the YEC model, some of which are more easily answered than others. YECs would certainly see junk DNA and pseudogenes as functional genetic elements, thereby explaining their shared presence in ape and human genomes. The challenge of modern human genetic diversity being unable to arise in just a few thousand years initially seemed formidable to any model of Adam and Eve, especially one where they lived only a few thousand years ago. However, as noted in Section 2.6, work by Sanford et al. (2018) proposes that human genetic diversity could arise very rapidly if Adam and Eve were created with initial diversity not only within their own genome but also in the genomes of their gametes. If they had many offspring, then it is argued that human genetic diversity could increase to modern-day levels in a few hundred generations—within the timespan allowed by the YEC model. Other YECs have offered similar arguments that Adam and Eve were created with initial diversity in their genome, which can account for modern human genetic diversity (Jeanson 2017). They also maintain that mutational degradation of the human genome implies that the human species cannot be more than a few thousand years old (Sanford 2005).
At first blush, the YEC model seems to encounter a chronological problem when it proposes that Neanderthals and Denisovans are fully human members of our species, descended from Adam and Eve. Under conventional dates, these groups lived tens or hundreds of thousands of years ago—long before the date YECs would give for the creation of Adam and Eve. However, YECs reject almost all standard dating methods which otherwise would place Neanderthals and Denisovans as living long before the date YECs give for the creation of Adam. Though there is not universal agreement, many YECs claim that fossil specimens of virtually all humanlike members of the genus Homo—from H. erectus to H. heidelbergensis to Neanderthals to Denisovans to modern humans—represent post-Flood human fossils that probably lived concurrently, just a few thousand years ago. Moreover, YECs admit no discrepancy between the appearance of the modern humanlike body plan in the fossil record (approximately 1.5 million years ago under conventional dates) and the first clear-cut evidence of modern humanlike intelligence in the archaeological record (approximately 100,000 years ago under conventional dates) because they reject the methods used to obtain those dates. Yet, this approach to resolving certain chronological problems by proposing near-wholesale rejection of scientific dating methods thus substitutes one set of problems for another, and generates major hurdles related to rejecting mainstream science which this model must overcome.
2.8. Old Earth/Recent Humans Hybrid Model
As the GAE model shows (Section 2.3), belief in a recent Adam and Eve who lived only a few thousand years ago does not necessarily entail belief in a young Earth. The Old Earth/Recent Humans Hybrid (“Hybrid”) model accepts a conventional age of the universe and earth of billions of years old, but proposes that God specially and miraculously created Adam and Eve very recently, perhaps as recent as 6000 to 10,000 years ago (Daae 2010, 2012). Unlike the GAE model, however, the Hybrid model does not hold that Adam and Eve’s progeny interbred with other pre-existing hominids. Rather, the Hybrid model postulates that there were no humanlike hominids prior to the creation of Adam and Eve, and that Adam and Eve are the sole progenitors of all humanity—which would include the Neanderthals and Denisovans as subraces of normal human beings.
The Hybrid model thus resembles the YEC model in virtually all respects except that it allows that the universe and earth (and potentially also non-human forms of life) predated the creation of Adam and Eve by millions if not billions of years. This model generally accepts conventional dating techniques for natural features other than hominid fossils, but holds that many humanlike hominid fossils are far younger than is conventionally believed. Proponents of this model would emphasize that fossil and archaeological remains of humans and humanlike hominids are so fragmented, sparse, and obscure that it is difficult to date them definitively.
2.8.1. Responses
The main motive underlying this model is to attempt a merging of traditional theological beliefs about Adam and Eve with a conventional view of the age of the universe and earth. Because it proposes a very recent Adam and Eve, the responses to the Hybrid model are thus virtually identical to responses to the YEC model—with the exception that the Hybrid model does not require challenges to the ages of the earth, universe, or other natural features (apart from humanlike hominid fossils).
It should be noted that some versions of the Hybrid model propose that all animal life was created recently (e.g., Gray 1997), but it seems possible to hold to an ancient Earth and ancient creation of non-human life, but then hold that only the human species was created very recently. Either viewpoint would therefore hold that there was no human death before sin. The version which holds that all animal life was created recently can claim that there was no death whatsoever prior to the fall of Adam and Eve. However, the position that non-human life existed long before Adam and Eve must accept that animal death existed prior to Adam and Eve’s sin. Defenses of this position from old Earth proponents are reviewed in Section 2.7.
While some details of this model can therefore vary, this model could allow that some early hominid fossils distinct from humans (e.g., the australopithecines) may in fact be much older than just a few thousand years in age. However, whenever a hominid species is related to humans (e.g., Neanderthals, Denisovans, etc.), this model holds they must descend from Adam and Eve and thereby have lived only within the last few thousand years. This model would therefore propose that the conventional dating of some, if not many, humanlike hominid fossils is inaccurate, as these fossils are said to be far younger than is typically believed. Whether it is feasible to reduce the ages of known humanlike hominid fossils to just a few thousand years is a major scientific hurdle this model must overcome.
3. Discussion
The eight models reviewed here have various strengths and weaknesses with regard to their respective approaches to incorporating traditional theological beliefs about human origins and scientific evidence regarding fossils, genetics, population genetics, archaeology, and chronology. Arguably, four of the models (TE/EC, Homo divinus, GAE, and Homo heidelbergensis) are evolutionary in that they involve humans evolving from apelike ancestors and sharing a common ancestor with apes, while the other four models (Unique Origins Design, OEC, YEC, and Hybrid) reject those evolutionary points.
Comparison of the Models
Table 1 below summarizes how these models interface with important traditional theological beliefs about Adam and Eve. The TE/EC model satisfies none of these beliefs, whereas only the YEC and Hybrid models satisfy all of them. The Homo divinus model, also an evolutionary model, proposes a historical Adam and Eve who lived recently, but because Adam and Eve are detached from any genealogical relationships to subsequent humans in this model, this recent origin is of unclear value. Some of the other models perform considerably better, as the Homo heidelbergensis, Unique Origins Design, and OEC models each satisfy four or five of the seven theological points—all of them missing a recent timing of the origin of Adam and Eve and no physical death before the fall. This latter point is only satisfied by the YEC and Hybrid models. Ironically, a recent origin of Adam and Eve is satisfied by the Homo divinus and GAE models—both evolutionary models. The GAE model is the only model to postulate that Adam and Eve are genealogical ancestors of all humans, but are not their sole initial progenitors.
Table 2 below summarizes how these models interface with mainstream scientific positions on human origins. Here, the TE/EC, Homo divinus, and GAE models perform best and satisfy all of the mainstream scientific points. The Homo heidelbergensis model satisfies at least five of the points, but diverges from mainstream science when it proposes that humanity went through a bottleneck of two individuals at the time of Adam and Eve, as mainstream science recognizes no such event. The Unique Origins Design model scores three points—two related to dating and one for recognizing that humans are related to Neanderthals and Denisovans. The OEC model also scores three points—two related to dating, and one for being the only non-evolutionary model to recognize that the human body plan appears before human intelligence in the fossil record. The YEC model scores only one point, related to humans and Neanderthals/Denisovans being related. The Hybrid model also scores this latter point, as well as a second point for an old age of the universe/earth.
Darwin skeptics have proposed many scientific arguments regarding human origins that might differ from the mainstream scientific consensus. However, they would argue that these points are nonetheless supported by peer-reviewed scientific papers and evidence. As such, these points ought to be considered in any evaluation of these models, even if they diverge from the standard scientific “consensus.” This evaluation is shown in Table 3. Here, the TE/EC, Homo divinus, and GAE models score zero points, underscoring the commitment of these models to never diverging from the scientific consensus. The Homo heidelbergensis model recognizes that population genetics models have found that human genetic diversity could arise from an initial pair, and also recognizes evidence of the full humanity of Neanderthals and Denisovans. The remaining four models (Unique Origins Design, OEC, YEC, and Hybrid models) all recognize the possibility of intelligent design in human history. Indeed, the Unique Origins Design, YEC, and Hybrid models recognize all of these alternative scientific points except for aligning the timing of the origin of humans in the fossil record with the first archaeological appearance of humanlike intelligence. Only the Unique Origins Design model accepts dating methods, yet proposes Adam and Eve could have been created when the humanlike body plan first appears, while only the OEC model accepts dating methods and proposes that Adam and Eve coincided with the first archaeological evidence of humanlike intelligence. These are important points distinguishing the OEC and Unique Origins Design models from other viewpoints.
Table 4 and Figure 1 present the totals from each of these tables. Table 4 also includes a subtotal from only Table 1 and Table 2, such that each models’ performance strictly from the vantage of mainstream theological views (Table 1) and mainstream scientific views (Table 2) can be evaluated.
In the subtotals, the GAE and Homo heidelbergensis models perform best because they incorporate mainstream scientific views about common ancestry and human evolution from apelike species in contrast to the other non-evolutionary models, while also acknowledging Adam and Eve as real historical people. The GAE model also allows for their de novo creation and recent origin, while the Homo heidelbergensis model allows them to be the sole ancestors of humans and denies interbreeding with non-human species. In the grand total analysis (Table 4, Figure 1), however, the Unique Origins and Design and Hybrid models tie for the highest scores, followed closely by the YEC model, which is followed closely by the OEC and Homo heidelbergensis models. The TE/EC model scores lowest, largely because it rejects traditional theological beliefs and other important scientific points worth considering in the conversation.
As noted, four of the models (TE/EC, Homo divinus, GAE, and Homo heidelbergensis) are evolutionary in that they involve common human–ape ancestry, while the other four models (Unique Origins Design, OEC, YEC, and Hybrid) favor or require the de novo/miraculous creation of humans. It is thus useful to compare the performance of evolutionary models vs. non-evolutionary models: Considering only Table 1 and Table 2 (i.e., the Subtotal in Table 4), the evolutionary models outperform, scoring an average of 9.25 points to 8 points for the non-evolutionary models. When Table 3 is incorporated into the totals, the non-evolutionary models outperform, scoring an average of 13.25 points compared with 9.75 points. This reflects the fact that the evolutionary models tend to score many mainstream scientific points but few theological points, whereas the non-evolutionary models tend to score both theological and scientific points.
The value of this scoring method is unclear, as the various theological and scientific points evaluated may not all have equivalent value. The alternative scientific points (Table 3) could also be selected differently. Which models a person prefers will likely depend upon whether they find it more important to align with certain traditional theological beliefs (Table 1), mainstream scientific beliefs (Table 2), other scientific beliefs (Table 3), or a combination of various beliefs from any of those broader categories. For example:
- If one cares very little about traditional theological beliefs regarding human origins and is convinced that mainstream evolutionary biology is correct, then the TE/EC model may be appealing.
- Conversely, if one places the highest priority upon traditional theological beliefs about Adam and Eve, then the YEC or Hybrid models may be most attractive.
For those incorporating both theological and scientific considerations into their positions, some of the newly conceived models incorporate multiple traditional theological views about Adam and Eve and also perform fairly well when compared with the scientific evidence. For example:
- If one finds evolutionary science persuasive but desires a form of historical Adam and Eve, then the Homo heidelbergensis, GAE, or Homo divinus models may be persuasive.
- If one finds evolutionary claims to be scientifically weak but accepts standard dating methods, then the Unique Origins Design or OEC models seem most viable. The Hybrid model is also available to those who accept an old Earth, though it requires rejecting dating methods as regards many hominid fossils.
A key scientific question distinguishing these models is how they resolve a potential discrepancy in the timing of the appearance of the humanlike body plan in the fossil record (conventionally dated to about 1–2 million years ago) versus the appearance of unambiguously modern humanlike intellectual activity in the archaeological record (conventionally dated to about 100,000 years ago). The TE/EC and GAE models accept these dates by adopting the standard evolutionary view that hominids experienced “mosaic” evolution, where the humanlike body plan evolved prior to the evolution of modern human cognitive and intellectual capacities. Under the GAE and Homo divinus models, Adam and Eve might even have lived long after modern human intelligence evolved.
The OEC model provides a simple solution to this conundrum by proposing that Neanderthals, Denisovans, and other hominids are not descended from Adam, allowing them to place the origin of Adam at the exact time of this archaeological “explosion” of humanlike creativity. Another potential resolution is to propose that humanlike creativity actually extends further back into the archaeological record than is commonly believed—a position supported by evidence of high intelligence in Neanderthals and other humanlike hominids, but not explored in detail here.7 Such a conclusion would lend support to both the Homo heidelbergensis and Unique Origins Design models, both of which propose an ancient Adam and Eve—though only the Unique Origins Design model allows Adam and Eve to potentially coincide with Homo erectus, when the humanlike body plan first appears in the fossil record. A final potential avenue for reconciling this apparent discrepancy is to question the dating of various hominid fossils and tools. Such a position would be found in the YEC model or Hybrid model. The YEC model is skeptical of not just evolutionary science, but virtually all dating methods. The Hybrid model questions the dating of very recent hominid fossils, but otherwise accepts the conventional ancient age of the earth.
A key outstanding theological question is whether the biblical genealogies can potentially allow for an Adam and Eve that lived 100,000, 500,000, or 750,000 years ago (or even earlier). This hinges on the incompleteness or extent of incompleteness of the genealogies. Such an ancient Adam and Eve would be required by the OEC, Homo heidelbergensis, and Unique Origins Design models. At present, the primary models which allow for a very recent Adam and Eve are the GAE, YEC, and Hybrid models. The GAE model, however, raises additional theological questions by postulating huge numbers of biologically modern humans that lived throughout much of human history but were not descended from Adam and Eve. The Homo divinus model also allows for a recent Adam and Eve; however, because it is divorced from genetics or genealogies, this is of unclear theological importance.
4. Conclusions
If recent decades are any indication, new models will likely be constructed in the future that adopt and refine the strongest elements that these current models have to offer. Whatever the final resolution to these questions, it seems clear that a variety of options already exist for Protestant Evangelicals and other Christians seeking to understand human origins and harmonize traditional theological beliefs with the scientific evidence. Although some models appear more successful than others at achieving a reconciliation, one need not jettison traditional beliefs about Adam and Eve in light of the findings of science.
Funding
Funding provided by the Discovery Institute.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
Jonathan Witt, Emily Reeves, Michael Keas, Brian Miller, John Bloom, and Steve Dilley are thanked for their helpful edits and feedback during the preparation of this article.
Conflicts of Interest
The author declares no conflict of interest.
| 1 | The intent of this section is not to discuss or critique all aspects of modern evolutionary biology, but simply to provide scientific commentary on specific common arguments from evolutionary creationists on the specific topic of human origins. |
| 2 | Swamidass’s (2019) descriptions of the GAE model contains various other statements which indicate the close association of his GAE model with an attempt to retain a standard evolutionary viewpoint:
|
| 3 | Schaffner (2021) stated: “we eventually learn that his [Craig’s] model allows for an unspecified amount of admixture from other hominin lineages into the descendants of Adam, eliminating the need for such a tight bottleneck. A more clearly stated hypothesis in this section would have saved the reader time and frustration.” |
| 4 | The concept of intelligent design entails an infusion of information that came from an intelligent mind. In the context of this model, intelligent design could inolve de novo origin/special creation, but it could also include taking the body plan of a pre-existing organism and infusing information into the genome such that a new type of organism suddenly emerged. Thus, at the very minimum, intelligent design requires some input of information which changes the genotype and phenotype of an organism. Again, this could mean de novo origin/special creation, but it does not necessarily require this. However, even in the case that design does not involve de novo origin/special creation, something special is still happening—namely the infusion of information into the genome which produces an informational discontinuity that is beyond the abilities of natural mechanisms to produce. |
| 5 | For example, the Scofield Refererence Bible, highly influential in American Evangelicalism and Fundamentalism, advocated the gap theory and day/age theory, which accept an ancient age of the earth. However, the Scofield Bible did not place a date on Adam and Eve, and seemingly allowed or assumed a recent Adam and Eve. Its theology of origins would probably be most compatible with the Hybrid model (Section 2.8). |
| 6 | |
| 7 | This evidence is controversial to some, including proponents of the classical OEC model. See Rana and Ross (2015). |
References
- Alexander, Denis. 2008. Creation or Evolution: Do We Have to Choose. Oxford: Monarch Books. [Google Scholar]
- Alper, Joe. 2003. Rethinking Neanderthals. Smithsonian Magazine. Available online: https://www.smithsonianmag.com/science-nature/rethinking-neanderthals-83341003/ (accessed on 8 March 2023).
- Arensburg, Baruch, Anne-Marie Tillier, Bernard Vandermeersch, Henri Duday, Lynne A. Schepartz, and Yoel Rak. 1989. A Middle Palaeolithic human hyoid bone. Nature 338: 758–60. [Google Scholar] [CrossRef] [PubMed]
- Ashton, John F., ed. 2000. In Six Days: Why Fifty Scientists Choose to Believe in Creation. Green Forest: Master Books. [Google Scholar]
- Badarinarayan, Smitha Srinivasachar, and Daniel Sauter. 2021. Switching Sides: How Endogenous Retroviruses Protect Us from Viral Infections. Journal of Virology 95: e02299-20. [Google Scholar] [CrossRef]
- Barrett, Matthew, and Ardel B. Caneday. 2013. Adam: To Be or Not To Be? In Four Views on The Historical Adam. Edited by Gregory A. Boyd and Philip G. Ryken. Grand Rapids: Zondervan, pp. 13–36. [Google Scholar]
- Barrick, William D. 2013a. Response from the Young-Earth View. In Four Views on The Historical Adam. Edited by Gregory A. Boyd and Philip G. Ryken. Grand Rapids: Zondervan, pp. 80–85. [Google Scholar]
- Barrick, William D. 2013b. A Historical Adam: Young-Earth Creation View. In Four Views on The Historical Adam. Edited by Gregory A. Boyd and Philip G. Ryken. Grand Rapids: Zondervan, pp. 197–227. [Google Scholar]
- Bar-Yosef, Ofer. 2002. The upper paleolithic revolution. Annual Review of Anthropology 31: 363–93. [Google Scholar] [CrossRef]
- Bednarik, Robert G. 1997. The earliest evidence of ocean navigation. The International Journal of Nautical Archaeology 26: 183–91. [Google Scholar] [CrossRef]
- Buggs, Richard. 2017. Comment at Adam, Eve and Population Genetics: A Reply to Dr. Richard Buggs (Part 1). The BioLogos Forum. November 18. Available online: https://discourse.biologos.org/t/adam-eve-and-population-genetics-a-reply-to-dr-richard-buggs-part-1/37039/61 (accessed on 8 March 2023).
- Buggs, Richard. 2018a. Adam and Eve: Lessons Learned. Nature Ecology & Evolution Blog. April 14. Available online: https://ecoevocommunity.nature.com/posts/32171-adam-and-eve-lessons-learned (accessed on 8 March 2023).
- Buggs, Richard. 2018b. Comment at Adam, Eve and Population Genetics: A Reply to Dr. Richard Buggs (Part 1). The BioLogos Forum. April 25. Available online: https://discourse.biologos.org/t/adam-eve-and-population-genetics-a-reply-to-dr-richard-buggs-part-1/37039/1061 (accessed on 8 March 2023).
- Buggs, Richard. 2018c. How Similar Are Human and Chimpanzee Genomes? RichardBuggs.com. July 14. Available online: https://richardbuggs.com/2018/07/14/how-similar-are-human-and-chimpanzee-genomes/ (accessed on 28 May 2023).
- Chase, Philip G., and April Nowell. 1998. Taphonomy of a Suggested Middle Paleolithic Bone Flute from Slovenia. Current Anthropology 39: 549–53. [Google Scholar] [CrossRef]
- Chazan, Michael. 2017. Toward a Long Prehistory of Fire. Current Anthropology 58: S351–S359. [Google Scholar] [CrossRef]
- Cheetham, Seth W., Geoffrey J. Faulkner, and Marcel E. Dinger. 2020. Overcoming challenges and dogmas to understand the functions of pseudogenes. Nature Reviews Genetics 21: 191–201. [Google Scholar] [CrossRef]
- Chuong, Edward B. 2018. The placenta goes viral: Retroviruses control gene expression in pregnancy. PLoS Biology 16: e3000028. [Google Scholar] [CrossRef] [Green Version]
- Cohen, Jon. 2007. Relative Differences: The Myth of 1%. Science 316: 1836. [Google Scholar] [CrossRef]
- Collins, C. John. 2011. Did Adam and Eve Really Exist Who They Were and Why You Should Care. Wheaton: Crossway. [Google Scholar]
- Collins, C. John. 2013. Historical Adam: Old Earth Creation View. In Four Views on The Historical Adam. Edited by Gregory A. Boyd and Philip G. Ryken. Grand Rapids: Zondervan, pp. 143–75. [Google Scholar]
- Collins, C. John. 2018. Reading Genesis Well: Navigating History, Poetry, Science, and Truth in Genesis 1–11. Grand Rapids: Zondervan Academic. [Google Scholar]
- Collins, Francis. 2006. The Language of God: A Scientist Presents Evidence for Belief. New York: Free Press. [Google Scholar]
- Conley, Andrew B., Jittima Piriyapongsa, and I. King Jordan. 2008. Retroviral promoters in the human genome. Bioinformatics 24: 1563–67. [Google Scholar] [CrossRef] [Green Version]
- Craig, William Lane. 2000. The Kalām Cosmological Argument. Eugene: Wipf and Stock. [Google Scholar]
- Craig, William Lane. 2001. Time and the Metaphysics of Relativity. Dordecht: Kluwer Academic. [Google Scholar]
- Craig, William Lane. 2021. In Quest of the Historical Adam: A Biblical and Scientific Exploration. Grand Rapids: Eerdmans. [Google Scholar]
- Craig, William Lane, and James Porter Moreland. 2012. The Blackwell Companion to Natural Theology. Sussex: Wiley-Blackwell. [Google Scholar]
- Craig, William Lane, and Quentin Smith. 1995. Theism, Atheism, and Big Bang Cosmology. Oxford: Clarendon Press. [Google Scholar]
- D’Anastasio, Ruggero, Stephen Wroe, Claudio Tuniz, Lucia Mancini, Deneb T. Cesana, Diego Dreossi, Mayoorendra Ravichandiran, Marie Attard, William C. H. Parr, Anne Agur, and et al. 2013. Micro-Biomechanics of the Kebara 2 Hyoid and Its Implications for Speech in Neanderthals. PLoS ONE 8: e82261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daae, H. Donald. 2010. Bridging the Gap: The 7th Day. Bloomington: Westbow Press. [Google Scholar]
- Daae, H. Donald. 2012. Bridging the Gap: The First 6 Days (Revised Edition). Bloomington: Westbow Press. [Google Scholar]
- Davis, Ted. 2012. Theistic Evolution: History and Beliefs. BioLogos. October 15. Available online: https://biologos.org/series/science-and-the-bible/articles/theistic-evolution-history-and-beliefs (accessed on 9 March 2023).
- Dembski, William A. 2009. The End of Christianity: Finding a Good God in an Evil World. Nashville: B&H Academic. [Google Scholar]
- Donald, Alistair. 2009. Evolution and the Church. In Should Christians Embrace Evolution: Biblical and Scientific Responses. Edited by Norman C. Nevin. Phillipsburg: P&R Publishing, pp. 15–26. [Google Scholar]
- Durrett, Rick, and Deena Schmidt. 2008. Waiting for Two Mutations: With Applications to Regulatory Sequence Evolution and the Limits of Darwinian Evolution. Genetics 180: 1501–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dykes, Sue. 2020a. What about the Genetic Similarity between Humans and Chimps? In Thinking About Evolution: 25 Questions Christians Want Answered. Edited by Anjeanette Roberts, Fazale Rana, Sue Dykes and Mark Perez. Covina: RTB, pp. 167–75. [Google Scholar]
- Dykes, Sue. 2020b. Are the Hominin Fossils Evidence for Human Evolution? In Thinking About Evolution: 25 Questions Christians Want Answered. Edited by Anjeanette Roberts, Fazale Rana, Sue Dykes and Mark Perez. Covina: RTB, pp. 177–87. [Google Scholar]
- Dykes, Sue. 2020c. Did Humans and Neanderthals Interbreed? In Thinking About Evolution: 25 Questions Christians Want Answered. Edited by Anjeanette Roberts, Fazale Rana, Sue Dykes and Mark Perez. Covina: RTB, pp. 189–99. [Google Scholar]
- Dykes, Sue. 2020d. Did Neanderthals Create Art? In Thinking About Evolution: 25 Questions Christians Want Answered. Edited by Anjeanette Roberts, Fazale Rana, Sue Dykes and Mark Perez. Covina: RTB, pp. 201–12. [Google Scholar]
- ENCODE Project Consortium. 2012. An Integrated Encyclopedia of DNA Elements in the Human Genome. Nature 489: 57–74. [Google Scholar] [CrossRef] [Green Version]
- Everett, Daniel. 2018. Did Homo erectus speak? Aeon. February 28. Available online: https://aeon.co/essays/tools-and-voyages-suggest-that-homo-erectus-invented-language (accessed on 24 March 2023).
- Falk, Darrel. 2004. Coming to Peace with Science: Bridging the Worlds Between Faith and Biology. Downers Grove: IVP Academic. [Google Scholar]
- Falk, Darrel. 2017. Hominid Fossils (evolutionary view). In Dictionary of Christianity and Science. Edited by Paul Copan, Tremper Longman III, Christopher L. Reese and Michael G. Strauss. Grand Rapids: Zondervan, pp. 355–58. [Google Scholar]
- Fiddes, Ian T., Gerrald A. Lodewijk, Meghan Mooring, Colleen M. Bosworth, Adam D. Ewing, Gary L. Mantalas, Adam M. Novak, Anouk van den Bout, Alex Bishara, Jimi L. Rosenkrantz, and et al. 2018. Human-Specific NOTCH2NL Genes Affect Notch Signaling and Cortical Neurogenesis. Cell 173: 1356–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folger, Tim, and Shanti Menon. 1997. …Or Much Like Us? Discover Magazine. Available online: https://www.discovermagazine.com/planet-earth/or-much-like-us (accessed on 8 March 2023).
- Gates, Alexander J., Deisy Morselli Gysi, Manolis Kellis, and Albert-László Barabási. 2022. A wealth of discovery built on the Human Genome Project—By the numbers. Nature 590: 212–15. [Google Scholar] [CrossRef]
- Gauger, Ann, Douglas Axe, and Casey Luskin. 2012. Science and Human Origins. Seattle: Discovery Institute Press. [Google Scholar]
- Gibbons, Ann. 1998. Ancient Island Tools Suggest Homo erectus Was a Seafarer. Science 279: 1635–37. [Google Scholar] [CrossRef]
- Gray, Gorman. 1997. The Age of the Universe: What Are the Biblical Limits? Washougal: Morning Star. [Google Scholar]
- Grudem, Wayne. 2017a. Biblical and Theological Introduction: The Incompatibility of Theistic Evolution with the Biblical Account of Creation and with Important Christian Doctrines. In Theistic Evolution: A Scientific, Philosophical, and Theological Critique. Edited by J. P. Moreland, Stephen C. Meyer, Christopher Shaw, Ann K. Gauger and Wayne Grudem. Wheaton: Crossway, pp. 61–77. [Google Scholar]
- Grudem, Wayne. 2017b. Theistic Evolution Undermines Twelve Creation Events and Several Crucial Christian Doctrines. In Theistic Evolution: A Scientific, Philosophical, and Theological Critique. Edited by J. P. Moreland, Stephen C. Meyer, Christopher Shaw, Ann K. Gauger and Wayne Grudem. Wheaton: Crossway, pp. 783–837. [Google Scholar]
- Habib, Adella M., Andrei L. Okorokov, Matthew N. Hill, Jose T. Bras, Man-Cheung Lee, Shengnan Li, Samuel J Gossage, Marie van Drimmelen, Maria Morena, Henry Houlden, and et al. 2019. Microdeletion in a FAAH pseudogene identified in a patient with high anandamide concentrations and pain insensitivity. British Journal of Anaesthesia 123: e249–e253. [Google Scholar] [CrossRef] [Green Version]
- Ham, Ken. 2006. Could God Really Have Created Everything in Six Days? In The New Answers Book 1: Over 25 Questions on Creation/Evolution and the Bible. Edited by Ken Ham. Green Forest: Master Books, pp. 88–112. [Google Scholar]
- Hawks, John, Keith Hunley, Sang-Hee Lee, and Milford Wolpoff. 2000. Population bottlenecks and Pleistocene human evolution. Molecular Biology and Evolution 17: 2–22. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, H., T. Arao, Y. Togashi, H. Kato, Y. Fujita, M. A. De Velasco, H. Kimura, K. Matsumoto, K. Tanaka, I. Okamoto, and et al. 2015. The OCT4 pseudogene POU5F1B is amplified and promotes an aggressive phenotype in gastric cancer. Oncogene 34: 199–208. [Google Scholar] [CrossRef]
- Henry, Hugh, and Daniel Dyke. 2012. From Noah to Abraham to Moses: Evidence of Genealogical Gaps in Mosaic Literature. Reasons to Believe. July 23. Available online: https://reasons.org/explore/publications/articles/from-noah-to-abraham-to-moses-evidence-of-genealogical-gaps-in-mosaic-literature-part-1 (accessed on 9 March 2023).
- Hirotsune, Shinji, Noriyuki Yoshida, Amy Chen, Lisa Garrett, Fumihiro Sugiyama, Satoru Takahashi, Ken-ichi Yagami, Anthony Wynshaw-Boris, and Atsushi Yoshiki. 2003. An expressed pseudogene regulates the messenger-RNA stability of its homologous coding gene. Nature 423: 91–96. [Google Scholar] [CrossRef]
- Hlubik, Sarah. Francesco Berna, Craig Feibel, David Braun, and John W. K. Harris. 2017. Researching the Nature of Fire at 1.5 Mya on the Site of FxJj20 AB, Koobi Fora, Kenya, Using High-Resolution Spatial Analysis and FTIR Spectrometry. Current Anthropology 58: S243–S257. [Google Scholar] [CrossRef]
- Hössjer, Ola, and Ann Gauger. 2019. A Single-Couple Human Origin is Possible. BIO-Complexity 2019: 1–20. [Google Scholar] [CrossRef]
- Hössjer, Ola, Ann Gauger, and Colin Reeves. 2016a. Genetic Modeling of Human History Part 1: Comparison of Common Descent and Unique Origin Approaches. BIO-Complexity 2016: 1–15. [Google Scholar] [CrossRef]
- Hössjer, Ola, Ann Gauger, and Colin Reeves. 2016b. Genetic Modeling of Human History Part 2: A Unique Origin Algorithm. BIO-Complexity 2016: 1–36. [Google Scholar] [CrossRef]
- Hössjer, Ola, Günter Bechly, and Ann Gauger. 2021. On the waiting time until coordinated mutations get fixed in regulatory sequences. Journal of Theoretical Biology 524: 110657. [Google Scholar] [CrossRef]
- Jacques, Pierre-Étienne, Justin Jeyakani, and Guillaume Bourque. 2013. The Majority of Primate-Specific Regulatory Sequences Are Derived from Transposable Elements. PLoS Genetics 9: e1003504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeanson, Nathanial T. 2017. Replacing Darwin: The New Origin of Species. Green Forest: Master Books. [Google Scholar]
- Ji, Zhe, Ruisheng Song, Aviv Regev, and Kevin Struhl. 2015. Many lncRNAs, 5′UTRs, and Pseudogenes Are Translated and Some Are Likely to Express Functional Proteins. eLife 4: e08890. [Google Scholar] [CrossRef]
- Jönsson, Marie E., Raquel Garza, Yogita Sharma, Rebecca Petri, Erik Södersten, Jenny G. Johansson, Pia A. Johansson, Diahann A. M. Atacho, Karolina Pircs, Sofia Madsen, and et al. 2021. Activation of endogenous retroviruses during brain development causes an inflammatory response. The EMBO Journal 40: e106423. [Google Scholar] [CrossRef] [PubMed]
- Keathley, Kenneth, and Mark Rooker. 2014. 40 Questions about Creation and Evolution. Grand Rapids: Kregel Academic. [Google Scholar]
- Kelly, Robert L., and David Hurst Thomas. 2010. Archaeology, 5th ed. Belmont: Wadsworth Cengage Learning. [Google Scholar]
- Kidner, Derek. 1967. Genesis: An Introduction and Commentary. Downers Grove: InterVarsity Press. [Google Scholar]
- Kim, Min-Sik, Sneha M. Pinto, Derese Getnet, Raja Sekhar Nirujogi, Srikanth S. Manda, Raghothama Chaerkady, Anil K. Madugundu, Dhanashree S. Kelkar, Ruth Isserlin, Shobhit Jain, and et al. 2014. A Draft Map of the Human Proteome. Nature 509: 575–81. [Google Scholar] [CrossRef] [Green Version]
- Kitchen, Kenneth Anderson. 1966. Ancient Orient and Old Testament. Downers Grove: InterVarsity Press. [Google Scholar]
- Kovalenko, T. F., and L. I. Patrushev. 2018. Pseudogenes as Functionally Significant Elements of the Genome. Biochemistry (Moscow) 83: 1332–49. [Google Scholar] [CrossRef]
- Lamoureux, Denis O. 2009. I Love Jesus & Accept Evolution. Eugene: Wipf and Stock. [Google Scholar]
- Lieberman, Daniel E., David R. Pilbeam, and Richard W. Wrangham. 2009. The Transition from Australopithecus to Homo. In Transitions in prehistory: Essays in Honor of Ofer Bar-Yosef. Edited by John J. Shea and Daniel E. Lieberman. Cambridge: Oxbow Books, pp. 1–22. [Google Scholar]
- Luskin, Casey. 2011. BioLogos Blog Author Helps Darwin’s Universal Acid Burn God Right Out of Religion. Evolution News. November 3. Available online: https://evolutionnews.org/2011/11/biologos_blog_author_helps_dar/ (accessed on 24 March 2023).
- Luskin, Casey. 2014. The New Theistic Evolutionists: BioLogos and the Rush to Embrace the ‘Consensus’. Christian Research Journal 37: 32–41. Available online: https://www.equip.org/article/new-theistic-evolutionists-biologos-rush-embrace-consensus/ (accessed on 8 March 2023).
- Luskin, Casey. 2022. Evolutionary models of palaeoanthropology, genetics and psychology fail to account for human origins: A review. In Science and Faith in Dialogue. Edited by Frikkie Van Niekerk and Nico Vorster. Cape Town: AOSIS, pp. 243–81. [Google Scholar] [CrossRef]
- Ma, Yanni, Siqi Liu, Jie Gao, Chunyan Chen, Xin Zhang, Hao Yuan, Zhongyang Chen, Xiaolin Yin, Chenguang Sun, Yanan Mao, and et al. 2021. Genome-wide Analysis of Pseudogenes Reveals HBBP1’s Human-specific Essentiality in Erythropoiesis and Implication in β-Thalassemia. Developmental Cell 56: 478–93. [Google Scholar] [CrossRef]
- Madueme, Hans. 2020. Evolution and Historical Adam? A Provocative but Unconvincing Attempt. The Gospel Coalition. March 2. Available online: https://www.thegospelcoalition.org/reviews/genealogical-adam-eve-swamidass/ (accessed on 8 March 2023).
- Mattick, John, and Paulo Amaral. 2023. RNA: The Epicenter of Genetic Information. Boca Raton: CRC/Taylor & Francis Press. [Google Scholar]
- Mayr, Ernst. 2004. What Makes Biology Unique?: Considerations on the Autonomy of a Scientific Discipline. Cambridge: Cambridge University Press. [Google Scholar]
- McLatchie, Jonathan. 2020. The Search for Adam and Eve: Human Origins According to Scripture and Science. JonathanMclatchi.com. December 8. Available online: https://jonathanmclatchie.com/the-search-for-adam-and-eve-human-origins-according-to-scripture-and-science/ (accessed on 8 March 2023).
- Mellars, Paul. 2004. Neanderthals and the modern human colonization of Europe. Nature 432: 461–65. [Google Scholar] [CrossRef] [PubMed]
- Meyer, Stephen C. 2009. Signature in the Cell: DNA and the Evidence for Intelligent Design. New York: Harper One. [Google Scholar]
- Miller, Kenneth. 2005. Day 1 AM testimony, Kitzmiller v. Dover trial. Harrisburg, PA: Middle District Court of Pennsylvania, September 26. Available online: https://web.archive.org/web/20220219142105/https://ncse.ngo/files/pub/legal/kitzmiller/trial_transcripts/2005_0926_day1_am.pdf (accessed on 3 June 2023).
- Moleirinho, Ana, Susana Seixas, Alexandra M. Lopes, Celeste Bento, Maria J. Prata, and António Amorim. 2013. Evolutionary Constraints in the β-Globin Cluster: The Signature of Purifying Selection at the δ-Globin (HBD) Locus and Its Role in Developmental Gene Regulation. Genome Biology and Evolution 5: 559–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, Henry M. 1976. The Genesis Record: A Scientific and Devotional Commentary on the Book of Beginnings. Grand Rapids: Baker Book House. [Google Scholar]
- Morris, Henry M. 1977. The Beginning of the World. Denver: Ascent Books. [Google Scholar]
- Muro, Enrique M., Nancy Mah, and Miguel A. Andrade-Navarro. 2011. Functional evidence of post-transcriptional regulation by pseudogenes. Biochimie 93: 1916–21. [Google Scholar] [CrossRef] [PubMed]
- Nowell, April. 2006. From a Paleolithic art to Pleistocene visual cultures (Introduction to two special issues on “Advances in the study of Pleistocene imagery and symbol use’). Journal of Archaeological Method and Theory 13: 239–49. [Google Scholar] [CrossRef]
- Ostling, Richard N. 2011. The Search for the Historical Adam. Christianity Today. June 3. Available online: https://www.christianitytoday.com/ct/2011/june/historicaladam.html (accessed on 8 March 2023).
- Pain, Debleena, Gung-Wei Chirn, Christopher Strassel, and Daniel M. Kemp. 2005. Multiple Retropseudogenes from Pluripotent Cell-specific Gene Expression Indicates a Potential Signature for Novel Gene Identification. The Journal of Biological Chemistry 280: 6265–68. [Google Scholar] [CrossRef] [Green Version]
- Pennisi, Elizabeth. 2012. ENCODE Project Writes Eulogy for Junk DNA. Science 377: 1160–61. [Google Scholar] [CrossRef]
- Piehler, Armin P., Marit Hellum, Jürgen J. Wenzel, Ellen Kaminski, Kari Bente Foss Haug, Peter Kierulf, and Wolfgang E. Kaminski. 2008. The human ABC transporter pseudogene family: Evidence for transcription and gene-pseudogene interference. BMC Genomics 9: 165. [Google Scholar] [CrossRef] [Green Version]
- Pink, Ryan Charles, Kate Wicks, Daniel Paul Caley, Emma Kathleen Punch, Laura Jacobs, and David Raul Francisco Carter. 2011. Pseudogenes: Pseudo-functional or key regulators in health and disease? RNA 17: 792–98. [Google Scholar] [CrossRef] [Green Version]
- Poliseno, Laura. 2012. Pseudogenes: Newly Discovered Players in Human Cancer. Science Signaling 5: re5. [Google Scholar] [CrossRef]
- Poliseno, Laura, Leonardo Salmena, Jiangwen Zhang, Brett Carver, William J. Haveman, and Pier Paolo Pandolfi. 2010. A coding-independent function of gene and pseudogene mRNAs regulates tumour biology. Nature 465: 1033–38. [Google Scholar] [CrossRef] [Green Version]
- Prieto-Godino, Lucia L., Raphael Rytz, Benoîte Bargeton, Liliane Abuin, J. Roman Arguello, Matteo Dal Peraro, and Richard Benton. 2016. Olfactory receptor pseudo-pseudogenes. Nature 539: 93–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rana, Fazale. 2018. When Did Modern Human Brains—and the Image of God—Appear? Reasons to Believe. November 14. Available online: https://reasons.org/explore/blogs/the-cells-design/when-did-modern-human-brains-and-the-image-of-god-appear (accessed on 3 June 2023).
- Rana, Fazale. 2020a. Is Evolution Really a Problem for the Christian Faith? In Thinking About Evolution: 25 Questions Christians Want Answered. Edited by Anjeanette Roberts, Fazale Rana, Sue Dykes and Mark Perez. Covina: RTB, pp. 65–73. [Google Scholar]
- Rana, Fazale. 2020b. Answering Scientific Questions on Neanderthal-Human Interbreeding, Part 1. Reasons.org Posted August 5. Available online: https://reasons.org/explore/blogs/the-cells-design/answering-scientific-questions-on-neanderthal-human-interbreeding-part-1 (accessed on 11 May 2023).
- Rana, Fazale. 2021. Ancient Cave Art Strengthens Evidence for the Image of God. Reasons to Believe. March 10. Available online: https://reasons.org/explore/blogs/the-cells-design/ancient-cave-art-strengthens-evidence-for-the-image-of-god (accessed on 3 June 2023).
- Rana, Fazale, and Hugh Ross. 2005. Who Was Adam: A Creation Model Approach to the Origin of Man. Colorado Springs: Navpress. [Google Scholar]
- Rana, Fazale, and Hugh Ross. 2015. Who Was Adam: A Creation Model Approach to the Origin of Man (Second Expanded Edition). Covina: RTB Press. [Google Scholar]
- Rapicavoli, Nicole A., Kun Qu, Jiajing Zhang, Megan Mikhail, Remi-Martin Laberge, and Howard Y. Chang. 2013. A mammalian pseudogene lncRNA at the interface of inflammation and anti-inflammatory therapeutics. eLife 2: e00762. [Google Scholar] [CrossRef] [PubMed]
- Reeves, Michael. 2009. Adam and Eve. In Should Christians Embrace Evolution: Biblical and Scientific Responses. Edited by Norman C. Nevin. Phillipsburg: P&R Publishing, pp. 43–56. [Google Scholar]
- Rice, Stanley A. 2007. Encyclopedia of Evolution. New York: Checkmark Books. [Google Scholar]
- Robson, Michael I., and Stefan Mundlos. 2019. Jumping retroviruses nudge TADs apart. Nature Genetics 51: 1304–5. [Google Scholar] [CrossRef] [PubMed]
- Ross, Hugh. 2001. The Genesis Question: Scientific Advances and the Accuracy of Genesis. Colorado Springs: NavPress. [Google Scholar]
- Ross, Hugh. 2014. Navigating Genesis: A Scientist’s Journey through Genesis 1–11. Covina: RTB Press. [Google Scholar]
- Ross, Hugh. 2016. When Did God Create Adam and Eve? Reasons.org Posted October 19. Available online: https://reasons.org/explore/blogs/todays-new-reason-to-believe-es/when-did-god-create-adam-and-eve (accessed on 29 May 2023).
- Rossano, Matt J. 2010. Supernatural Selection: How Religion Evolved. Oxford: Oxford University Press. [Google Scholar]
- Sakashita, Akihiko, Tomohiro Kitano, Hirotsugu Ishizu, Youjia Guo, Harumi Masuda, Masaru Ariura, Kensaku Murano, and Haruhiko Siomi. 2023. Transcription of MERVL retrotransposons is required for preimplantation embryo development. Nature Genetics 55: 484–95. [Google Scholar] [CrossRef]
- Salmena, Leonardo, Laura Poliseno, Yvonne Tay, Lev Kats, and Pier Paolo Pandolfi. 2011. A ceRNA Hypothesis: The Rosetta Stone of a Hidden RNA Language? Cell 146: 353–58. [Google Scholar] [CrossRef] [Green Version]
- Sanford, John. 2005. Genetic Entropy & The Mystery of the Genome. Lima: Elim. [Google Scholar]
- Sanford, John, Rob Carter, Wes Brewer, John Baumgardner, Bruce Potter, and John Potter. 2018. Adam and Eve, Designed Diversity, and Allele Frequencies. In Proceedings of the Eighth International Conference on Creationism. Edited by J.H. Whitmore. Pittsburgh: Creation Science Fellowship, pp. 200–16. [Google Scholar]
- Schaffner, Stephen. 2021. Adam, Eve, and the evolution of humankind. Science 374: 162. [Google Scholar] [CrossRef]
- Scott, Eugenie. 2007. What Do Creationists Believe about Human Evolution?|Dr. Eugenie Scott. The Leakey Foundation, YouTube Posted November 29, 2021. Available online: https://www.youtube.com/watch?v=iZhO7_GhDH0 (accessed on 8 March 2023).
- Seaman, Josiah, and Richard Buggs. 2020. FluentDNA: Nucleotide visualization of whole genomes, annotations, and alignments. Frontiers in Genetics 11: a292. [Google Scholar] [CrossRef]
- Stitz, Maria, Cristian Chaparro, Zhigang Lu, V. Janett Olzog, Christina E. Weinberg, Jochen Blom, Alexander Goesmann, Christoph Grunau, and Christoph G. Grevelding. 2021. Satellite-Like W-Elements: Repetitive, Transcribed, and Putative Mobile Genetic Factors with Potential Roles for Biology and Evolution of Schistosoma mansoni. Genome Biology and Evolution 13: evab204. [Google Scholar] [CrossRef]
- Stott, John R.W. 1999. Understanding the Bible: Expanded Edition. Grand Rapids: Zondervan. First published 1972. [Google Scholar]
- Stringer, Christopher B. 1992. Evolution of Early Humans. In Cambridge Encyclopedia of Human Evolution. Edited by Steve Jones, Robert Martin and David Pilbeam. Cambridge: Cambridge University Press, pp. 241–51. [Google Scholar]
- Suzuki, Ikuo K., David Gacquer, Roxane Van Heurck, Devesh Kumar, Marta Wojno, Angéline Bilheu, Adèle Herpoel, Nelle Lambert, Julian Cheron, Franck Polleux, and et al. 2018. Human-Specific NOTCH2NL Genes Expand Cortical Neurogenesis through Delta/Notch Regulation. Cell 173: 1370–84. [Google Scholar] [CrossRef] [Green Version]
- Swamidass, S. Joshua. 2017. Heliocentric Certainty Against a Bottleneck of Two? Peaceful Science. Available online: https://discourse.peacefulscience.org/t/heliocentric-certainty-against-a-bottleneck-of-two/61 (accessed on 8 March 2023).
- Swamidass, S. Joshua. 2019. The Genealogical Adam and Eve: The Surprising Science of Universal Ancestry. Downers Grove: IVP Academic. [Google Scholar]
- Tam, Oliver H., Alexei A. Aravin, Paula Stein, Angelique Girard, Elizabeth P. Murchison, Sihem Cheloufi, Emily Hodges, Martin Anger, Ravi Sachidanandam, Richard M. Schultz, and et al. 2008. Pseudogene-derived small interfering RNAs regulate gene expression in mouse oocytes. Nature 453: 534–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, João C., Guy S. Jacobs, Chris Stringer, Jonathan Tuke, Georgi Hudjashov, Gludhug A. Purnomo, Herawati Sudoyo, Murray P. Cox, Raymond Tobler, Chris S. M. Turney, and et al. 2021. Widespread Denisovan ancestry in Island Southeast Asia but no evidence of substantial super-archaic hominin admixture. Nature Ecology & Evolution 5: 616–24. [Google Scholar] [CrossRef]
- Thorvaldsen, Steinar, and Ola Hössjer. 2020. Using statistical methods to model the fine-tuning of molecular machines and systems. Journal of Theoretical Biology 501: 110352. [Google Scholar] [CrossRef]
- Toth, Nicholas, and Kathy Schick. 2015. Overview of paleolithic archaeology. In Handbook of Paleoanthropology: Principles, Methods, and Approaches, 2nd ed. Edited by Winfried Henke and Ian Tattersall. Heidelberg: Springer, pp. 2441–64. [Google Scholar] [CrossRef]
- Travis, Melissa Cain. 2021. William Lane Craig Explores the Headwaters of the Human Race. Christianity Today. September 20. Available online: https://www.christianitytoday.com/ct/2021/october/william-lane-craig-quest-historical-adam-genesis-origins.html (accessed on 8 March 2023).
- Trinkaus, Erik, and Cidália Duarte. 2003. The Hybrid Child from Portugal. Scientific American 288: 32. [Google Scholar]
- Trinkaus, Erik, and Pat Shipman. 1993. Neandertals: Images of Ourselves. Evolutionary Anthropology 1: 194–201. [Google Scholar] [CrossRef]
- Troskie, Robin-Lee, Geoffrey J. Faulkner, and Seth W. Cheetham. 2021. Processed pseudogenes: A substrate for evolutionary innovation. BioEssays 43: 2100186. [Google Scholar] [CrossRef] [PubMed]
- University of Michigan News Service. 2000. New Study Suggests Big Bang Theory of Human Evolution. January 10. Available online: https://news.umich.edu/new-study-suggests-big-bang-theory-of-human-evolution/ (accessed on 8 March 2023).
- Venema, Dennis. 2018. Comment at Adam, Eve and Population Genetics: A Reply to Dr. Richard Buggs (Part 1). The BioLogos Forum. April 25. Available online: https://discourse.biologos.org/t/adam-eve-and-population-genetics-a-reply-to-dr-richard-buggs-part-1/37039/1063 (accessed on 8 March 2023).
- Venema, Dennis, and Darrel Falk. 2010. Signature in the Pseudogenes. BioLogos. April 19. Available online: https://biologos.org/articles/signature-in-the-pseudogenes (accessed on 8 March 2023).
- Venema, Dennis, and Scot McKnight. 2017. Adam and the Genome: Reading Scripture after Genetic Science. Grand Rapids: Brazos Press. [Google Scholar]
- Villanea, Fernando A., and Joshua G. Schraiber. 2018. Multiple episodes of interbreeding between Neanderthal and modern humans. Nature Ecology & Evolution 3: 39–44. [Google Scholar] [CrossRef]
- Walton, John H. 2013. A Historical Adam: Archetypal Creation View. In Four Views on The Historical Adam. Edited by Gregory A. Boyd and Philip G. Ryken. Grand Rapids: Zondervan, pp. 89–118. [Google Scholar]
- Waters, Guy Prentiss. 2017. Theistic Evolution Is Incompatible with the Teachings of the New Testament. In Theistic Evolution: A Scientific, Philosophical, and Theological Critique. Edited by J. P. Moreland, Stephen C. Meyer, Christopher Shaw, Ann K. Gauger and Wayne Grudem. Wheaton: Crossway, pp. 879–926. [Google Scholar]
- Wen, Yan-Zi, Ling-Ling Zheng, Liang-Hu Qu, Francisco J. Ayala, and Zhao-Rong Lun. 2012. Pseudogenes Are Not Pseudo Any More. RNA Biology 9: 27–32. [Google Scholar] [CrossRef] [Green Version]
- White, Randall. 2003. Prehistoric Art: The Symbolic Journey of Humankind. New York: Harry N Abrams. [Google Scholar]
- Wong, Kate. 2003. Who were the Neandertals? Scientific American 288: 28–37. [Google Scholar] [CrossRef]
- Wood, Bernard, and Terry Harrison. 2011. The evolutionary context of the first hominins. Nature 470: 347–52. [Google Scholar] [CrossRef]
- Wrangham, Richard, and Rachel Carmody. 2010. Human Adaptation to the Control of Fire. Evolutionary Anthropology 19: 187–99. [Google Scholar] [CrossRef] [Green Version]
- 2012. ENCODE: The Rough Guide to the Human Genome. Discover. September 5. Available online: https://www.discovermagazine.com/the-sciences/encode-the-rough-guide-to-the-human-genome (accessed on 8 March 2023).
- Zhang, Jingyu, Xia Wang, Meixiang Li, Jin Han, Bing Chen, Bin Wang, and Jianwu Dai. 2006. NANOGP8 is a retrogene expressed in cancers. FEBS Journal 273: 1723–30. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Maohua, Congling Shi, Xing Gao, Xinzhi Wu, Fuyou Chen, Shuangquan Zhang, Xingkai Zhang, and John W. Olsen. 2014. On the possible use of fire by Homo erectus at Zhoukoudian, China. Chinese Science Bulletin 59: 335–43. [Google Scholar] [CrossRef]
Figure 1.
Comparison of scores and point totals of the models.


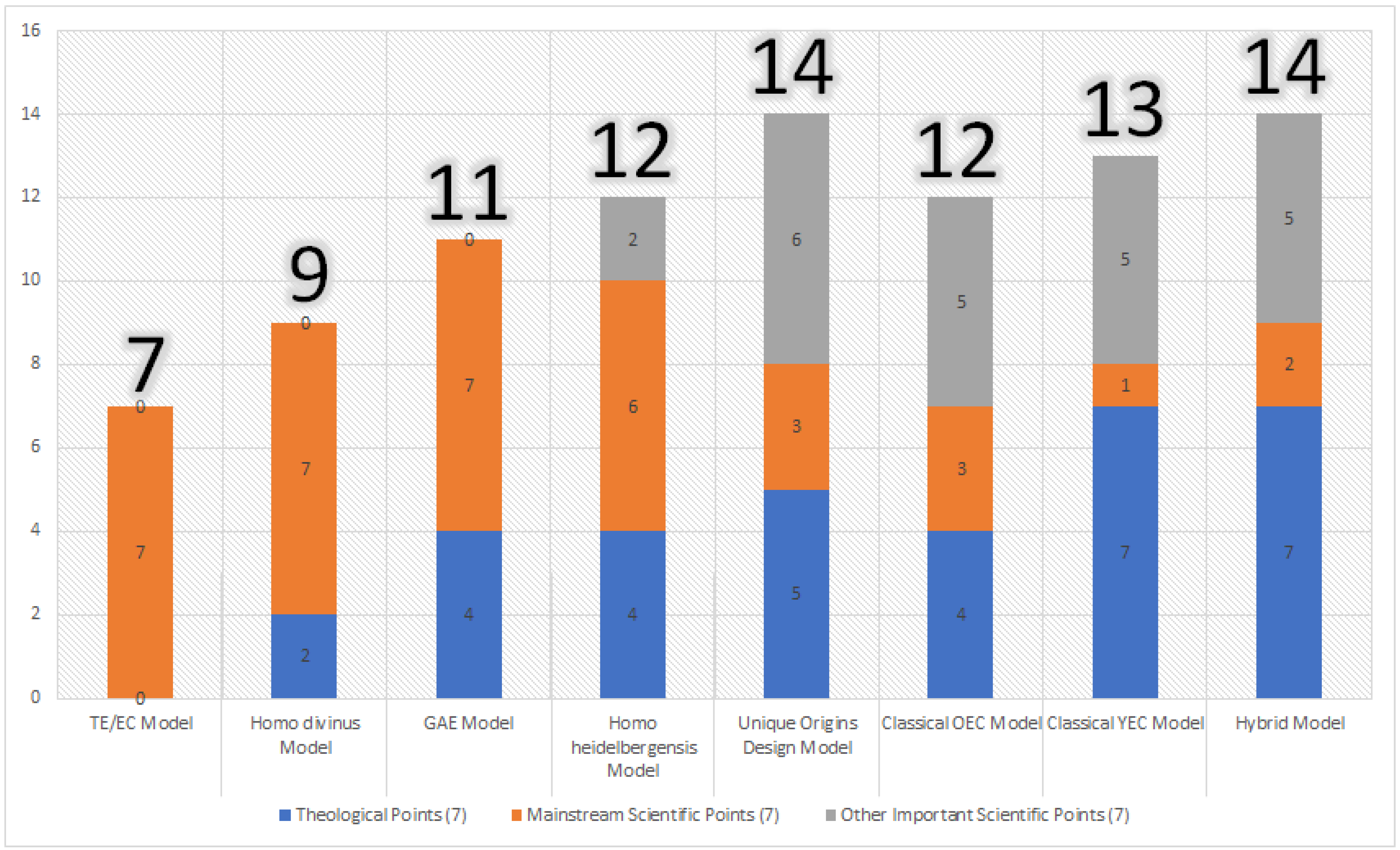{kind=link}
Table 1.
Comparison of model interaction with traditional theological beliefs about Adam and Eve.
| Model | Adam and Eve Were Real Historical People Who Fell into Sin. | Adam and Eve Were Ancestral to All Living Humans. | Adam and Eve Were the Sole Initial Progenitors of All Humans. | Adam and Eve Were Specially and Miraculously Created “de novo.” | Adam and Eve Lived Recently (<10,000 Years) as Genealogies Typically Suggest. | No Physical Death before the Fall of Adam and Eve | Humans Only Interbreed with Imago Dei Humans (i.e., Bestiality Not Required). |
|---|---|---|---|---|---|---|---|
| Classical Theistic Evolution/Evolutionary Creation Model | |||||||
| Homo divinus Model | X | X | |||||
| Genealogical Adam and Eve Model | X | X | Possible, if not favored, but not required. | Quite possible, but not required. | |||
| Homo heidelbergensis Model | X | X | Strongly preferred, but not required. | Probably not. | Strongly preferred, but not required. | ||
| Unique Origins Design Model | X | X | X | Allowed, if not favored. | X | ||
| Classical Old Earth Creationist Model | X | X | X | X | This was initially strongly preferred, but was rejected after evidence of human–Neanderthal interbreeding. | ||
| Classical Young Earth Creationist Model | X | X | X | X | X | X | X |
| Old Earth/Recent Origin Hybrid Model | X | X | X | X | X | Possible, but not required. | X |
Table 2.
Comparison of model interaction with mainstream scientific positions on human origins.
| Model | Humans Evolved via Standard/Unguided Evolutionary Mechanisms from Apelike Species. | Humans Share a Common Ancestor with Apes. | Humans Had Thousands of Primordial Ancestors and Never a Bottleneck of Two. | Humans (Homo sapiens) Are Related to Neanderthals and Denisovans. | Human Body Plan Appears before Human Intelligence. | Dating (Old Age) of Hominid Fossils Is Accurate. | Dating (Old Age) of Earth and Universe Is Accurate. |
|---|---|---|---|---|---|---|---|
| Classical Theistic Evolution/Evolutionary Creation Model | X | X | X | X | X | X | X |
| Homo divinus Model | X | X | X | X | X | X | X |
| Genealogical Adam and Eve Model | X | X | X | X | X | X | X |
| Homo heidelbergensis Model | Not clear, but seems to be possible. | X | X | X | X | X | |
| Unique Origins Design Model | X | X | X | ||||
| Classical Old Earth Creationist Model | X | X | X | ||||
| Classical Young Earth Creationist Model | X | ||||||
| Old Earth/Recent Origin Hybrid Model | X | X |
Table 3.
Other important scientific points.
| Model | Acknowledges Waiting Times Problem for Evolving Complex Traits and the Possibility of Design. | Incorporates Population Genetics Modeling Showing Humans Can Come from an Initial Pair. | Incorporates Evidence of Pseudogene Function. | Accepts Evidence of Full Humanity of Neanderthals/Denisovans. | Acknowledges Fossil Gap between Human and Apelike Species. | Adam and Eve Aligned with First Appearance of Fossil Evidence of Humanlike Body Plan. | Adam and Eve Aligned with First Appearance of Archaeological Evidence of Humanlike Intelligence. |
|---|---|---|---|---|---|---|---|
| Classical Theistic Evolution/Evolutionary Creation Model | |||||||
| Homo divinus Model | |||||||
| Genealogical Adam and Eve Model | |||||||
| Homo heidelbergensis Model | X | X | |||||
| Unique Origins Design Model | X | X | X | X | X | Possible. | |
| Classical Old Earth Creationist Model | X | X | X | X | X | ||
| Classical Young Earth Creationist Model | X | X | X | X | X | Not if standard dating methods are adopted. | Not if standard dating methods are adopted. |
| Old Earth/Recent Origin Hybrid Model | X | X | X | X | X | Not if standard dating methods are adopted. | Not if standard dating methods are adopted. |
Table 4.
Point totals for the models.
| Model | Theological Points (7) | Mainstream Scientific Points (7) | Subtotal (14) of Table 1 and Table 2 | Other Scientific Points (7) | Grand Total (21) |
|---|---|---|---|---|---|
| Classical Theistic Evolution/Evolutionary Creation Model | 0 | 7 | 7 | 0 | 7 |
| Homo divinus Model | 2 | 7 | 9 | 0 | 9 |
| Genealogical Adam and Eve Model | 4 | 7 | 11 | 0 | 11 |
| Homo heidelbergensis model | 4 | 6 | 10 | 2 | 12 |
| Unique Origins Design Model | 5 | 3 | 8 | 6 | 14 |
| Classical Old Earth Creationist Model | 4 | 3 | 7 | 5 | 12 |
| Classical Young Earth Creationist Model | 7 | 1 | 8 | 5 | 13 |
| Old Earth/Recent Origin Hybrid Model | 7 | 2 | 9 | 5 | 14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Luskin, C. Comparing Contemporary Evangelical Models Regarding Human Origins. Religions 2023, 14, 748. https://doi.org/10.3390/rel14060748
AMA Style
Luskin C. Comparing Contemporary Evangelical Models Regarding Human Origins. Religions. 2023; 14(6):748. https://doi.org/10.3390/rel14060748
Chicago/Turabian StyleLuskin, Casey. 2023. "Comparing Contemporary Evangelical Models Regarding Human Origins" Religions 14, no. 6: 748. https://doi.org/10.3390/rel14060748
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.