
LC–MS/MS Analysis of Ciguatoxins Revealing the Regional and Species Distinction of Fish in the Tropical Western Pacific

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Reference Toxins
2.3. Fish Specimens and Sample Preparation
2.4. LC–MS/MS Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yasumoto, T.; Nakajima, I.; Bagnis, R.; Adachi, R. Finding of a Dinoflagellate as a likely culprit of Ciguatera. Bull. Jap. Soc. Sci. Fish. 1977, 43, 1021–1026. [Google Scholar] [CrossRef] [Green Version]
- Chinain, M.; Gatti, C.M.; Roué, M.; Darius, H.T.; Subba Rao, D. Ciguatera-causing dinoflagellates in the genera Gambierdiscus and Fukuyoa: Distribution, ecophysiology and toxicology. In Dinoflagellates: Classification, Evolution, Physiology and Ecological Significance; Durvasula, S.R.V., Ed.; Nova Science Publishers: New York, NY, USA, 2020; pp. 405–457. [Google Scholar]
- Yasumoto, T. Chemistry, etiology, and food chain dynamics of marine toxins. Proc. Jpn. Acad. Ser. B 2005, 81, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Ikehara, T.; Kuniyoshi, K.; Oshiro, N.; Yasumoto, T. Biooxidation of Ciguatoxins Leads to Species-Specific Toxin Profiles. Toxins 2017, 9, 205. [Google Scholar] [CrossRef] [Green Version]
- Yogi, K.; Oshiro, N.; Inafuku, Y.; Hirama, M.; Yasumoto, T. Detailed LC-MS/MS Analysis of Ciguatoxins revealing distinct regional and species characteristics in fish and Causative Alga from the Pacific. Anal. Chem. 2011, 83, 8886–8891. [Google Scholar] [CrossRef]
- Yogi, K.; Oshiro, N.; Matsuda, S.; Sakugawa, S.; Matsuo, T.; Yasumoto, T. Toxin profiles in fish implicated in Ciguatera fish poisoning in Amami and Kakeroma Islands, Kagoshima Prefecture, Japan. Shokuhin Eiseigaku Zasshi (Food Hyg. Saf. Sci.) 2013, 54, 385–391. [Google Scholar] [CrossRef] [Green Version]
- Yogi, K.; Sakugawa, S.; Oshiro, N.; Ikehara, T.; Sugiyama, K.; Yasumoto, T. Determination of toxins involved in Ciguatera fish poisoning in the Pacific by LC/MS. J. AOAC Int. 2014, 97, 398–402. [Google Scholar] [CrossRef]
- Oshiro, N.; Tomikawa, T.; Kuniyoshi, K.; Kimura, K.; Kojima, T.; Yasumoto, T.; Asakura, H. Detection of ciguatoxins from the fish introduced to a wholesale market in Japan. Shokuhin Eiseigaku Zasshi (Food Hyg. Saf. Sci.) 2021, 62, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Igarashi, T.; Legrand, A.-M.; Cruchet, P.; Chinain, M.; Fujita, T.; Naoki, H. Structural elucidation of ciguatoxin congeners by fast-atom bombardment tandem mass spectroscopy. J. Am. Chem. Soc. 2000, 122, 4988–4989. [Google Scholar] [CrossRef]
- Murata, M.; Legrand, A.M.; Ishibashi, Y.; Yasumoto, T. Structures of ciguatoxin and its congener. J. Am. Chem. Soc. 1989, 111, 8929–8931. [Google Scholar] [CrossRef]
- Murata, M.; Legrand, A.M.; Ishibashi, Y.; Fukui, M.; Yasumoto, T. Structures and configurations of ciguatoxin from the moray eel Gymnothorax javanicus and its likely precursor from the dinoflagellate Gambierdiscus toxicus. J. Am. Chem. Soc. 1990, 112, 4380–4386. [Google Scholar] [CrossRef]
- Satake, M.; Murata, M.; Yasumoto, T. The structure of CTX3C, a ciguatoxin congener isolated from cultured Gambierdiscus toxicus. Tetrahedron Lett. 1993, 34, 1975–1978. [Google Scholar] [CrossRef]
- Satake, M.; Ishibashi, Y.; Legrand, A.-M.; Yasumoto, T. Isolation and structure of Ciguatoxin-4A, a new Ciguatoxin precursor, from cultures of Dinoflagellate Gambierdiscus toxicus and Parrotfish Scarus gibbus. Biosci. Biotechnol. Biochem. 1997, 60, 2103–2105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satake, M.; Fukui, M.; Legrand, A.-M.; Cruchet, P.; Yasumoto, T. Isolation and structures of new ciguatoxin analogs, 2,3-dihydroxyCTX3C and 51-hydroxyCTX3C, accumulated in tropical reef fish. Tetrahedron Lett. 1998, 39, 1197–1198. [Google Scholar] [CrossRef]
- Kato, T.; Yasumoto, T. Quantification of representative Ciguatoxins in the Pacific using quantitative nuclear magnetic resonance spectroscopy. Mar. Drugs 2017, 15, 309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halstead, B.W. Poisonous and Venomous Marine Animals of the World; Darwin Press, Inc.: Princeton, UK, 1988. [Google Scholar]
- EFSA Panel on Contaminants in the Food Chain, Scientific Opinion on marine biotoxins in shellfish–Emerging toxins: Ciguatoxin group. EFSA J. 2010, 8, 1627.
- Skinner, M.P.; Brewer, T.D.; Johnstone, R.; Fleming, L.E.; Lewis, R.J. Ciguatera fish poisoning in the Pacific Islands (1998 to 2008). PLoS Negl. Trop. Dis. 2011, 5, e1416. [Google Scholar] [CrossRef]
- Lewis, N.D. Epidemiology and impact of Ciguatera in the Pacific: A review. Mar. Fish. Rev. 1986, 48, 6–13. [Google Scholar]
- Lin, W.-F.; Lyu, Y.-C.; Wu, Y.-J.; Lu, C.-H.; Hwang, D.-F. Species identification of snapper: A food poisoning incident in Taiwan. Food Control 2012, 25, 511–515. [Google Scholar] [CrossRef]
- Sintunawa, C. Surveillance of marine fish for ciguatera toxin at fish markets in Bangkok, Thailand. Asian BioMed 2014, 8, 263. [Google Scholar] [CrossRef] [Green Version]
- Chan, T.Y. Ciguatera Fish Poisoning in East Asia and Southeast Asia. Mar. Drugs 2015, 13, 3466–3478. [Google Scholar] [CrossRef] [Green Version]
- Friedman, M.; Fernandez, M.; Backer, L.; Dickey, R.; Bernstein, J.; Schrank, K.; Kibler, S.; Stephan, W.; Gribble, M.; Bienfang, P.; et al. An updated review of Ciguatera fish poisoning: Clinical, epidemiological, environmental, and public health management. Mar. Drugs 2017, 15, 72. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, M. Ciguatera poisoning in North-West Viti Levu, Fiji Islands. Hawaii Med. J. 1975, 34, 207–210. [Google Scholar]
- Narayan, Y. Fish Poisoning in Fiji. Fiji Med. J. 1980, 8, 254–258. [Google Scholar]
- Nahar, J.; Sharma, N.A.; Kumar, K.; Prasad, A.; Kumar, A. In Fishermen’s Expert Views on the Causes of Fish Poisoning in Fiji: An Investigation through Data Mining Technique. In Proceedings of the 2017 4th Asia-Pacific World Congress on Computer Science and Engineering (APWC on CSE), Nadi, Fiji, 11–13 December 2017; IEEE: Piscataway, NJ, USA, 2017; pp. 99–105. [Google Scholar]
- Oshiro, N.; Yogi, K.; Asato, S.; Sasaki, T.; Tamanaha, K.; Hirama, M.; Yasumoto, T.; Inafuku, Y. Ciguatera incidence and fish toxicity in Okinawa, Japan. Toxicon 2010, 56, 656–661. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Collection Site | Number of Specimens |
|---|---|
| Keelung, Taiwan | 6 |
| Hat Yai, Thailand | 10 |
| Manila, the Philippines | 7 |
| The Viti Levu Island, Fiji | 19 |
| Total | 42 |
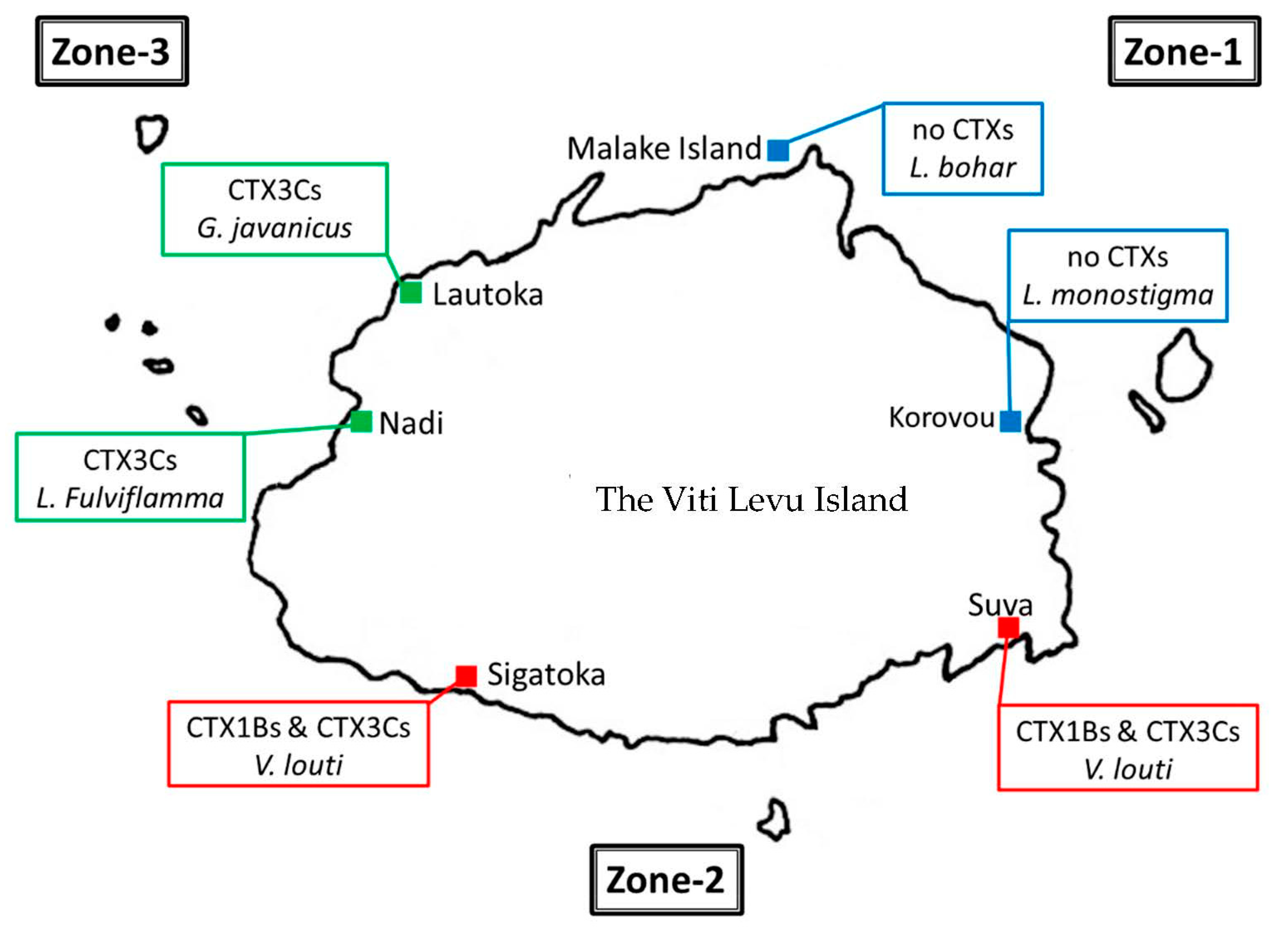
| Species | SL 1 (mm) | BW 2 (g) | Site 3 |
|---|---|---|---|
| Lutjanus argentimaculatus | 41 | 1295 | Korovou |
| Lutjanus monostigma | 27 | 640 | Korovou |
| Lutjanus fulviflamma | 17 | 120 | Nadi |
| 21 | 205 | Nadi | |
| 19 | 195 | Nadi | |
| Lutjanus bohar | 38 | 2150 | Malake |
| Lutjanus sp. | 21 | 635 | Sigatoka |
| Epinephelus fuscoguttatus | - 4 | 1515 | Lautoka |
| Plectropomus leopardus | 31 | 725 | Korovou |
| 27 | 455 | Korovou | |
| 22 | 210 | Korovou | |
| 33 | 750 | Suva | |
| - | 1575 | Suva | |
| Anyperodon leucogrammicus | - | 515 | Sigatoka |
| Variola louti | - | 1780 | Sigatoka |
| 37 | 1465 | Suva | |
| Upeneus sp. | - | 360 | Suva |
| Plectorhynchus chaetodontoides | 22 | 395 | Suva |
| Gymnothorax javanicus | 86 | 1845 | Lautoka |
| Species | Body Weight (g) | Collection Cite | Levels of CTXs in the Specimens (µg/kg) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1B 1 | Epi- Deoxy1b 2 | Deoxy1B 3 | OH-3C 4 | epi3C 5 | 3C 6 | Total | 1B eq 7 | |||
| V. louti | 1780 | Sigatoka | 0.078 | 0.065 | 0.105 | - 8 | + 9 | + | 0.25 | 0.13 |
| 1465 | Suva | 0.033 | 0.022 | 0.028 | - | + | + | 0.083 | 0.048 | |
| L. fulviflamma | 205 | Nadi | - | - | - | - | + | + | + | |
| 195 | Nadi | - | - | - | - | + | + | + | ||
| G. javanicus | 1845 | Lautoka | - | - | - | + | 0.054 | 0.069 | 0.079 | 0.025 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oshiro, N.; Tomikawa, T.; Kuniyoshi, K.; Ishikawa, A.; Toyofuku, H.; Kojima, T.; Asakura, H. LC–MS/MS Analysis of Ciguatoxins Revealing the Regional and Species Distinction of Fish in the Tropical Western Pacific. J. Mar. Sci. Eng. 2021, 9, 299. https://doi.org/10.3390/jmse9030299
Oshiro N, Tomikawa T, Kuniyoshi K, Ishikawa A, Toyofuku H, Kojima T, Asakura H. LC–MS/MS Analysis of Ciguatoxins Revealing the Regional and Species Distinction of Fish in the Tropical Western Pacific. Journal of Marine Science and Engineering. 2021; 9(3):299. https://doi.org/10.3390/jmse9030299
Chicago/Turabian StyleOshiro, Naomasa, Takumi Tomikawa, Kyoko Kuniyoshi, Akira Ishikawa, Hajime Toyofuku, Takashi Kojima, and Hiroshi Asakura. 2021. "LC–MS/MS Analysis of Ciguatoxins Revealing the Regional and Species Distinction of Fish in the Tropical Western Pacific" Journal of Marine Science and Engineering 9, no. 3: 299. https://doi.org/10.3390/jmse9030299