
Bidenichthys okamotoi, a New Species of the Bythitidae (Ophidiiformes, Teleostei) from the Koko Seamount, Central North Pacific †


Abstract
:1. Introduction
2. Materials and Methods
Study Area
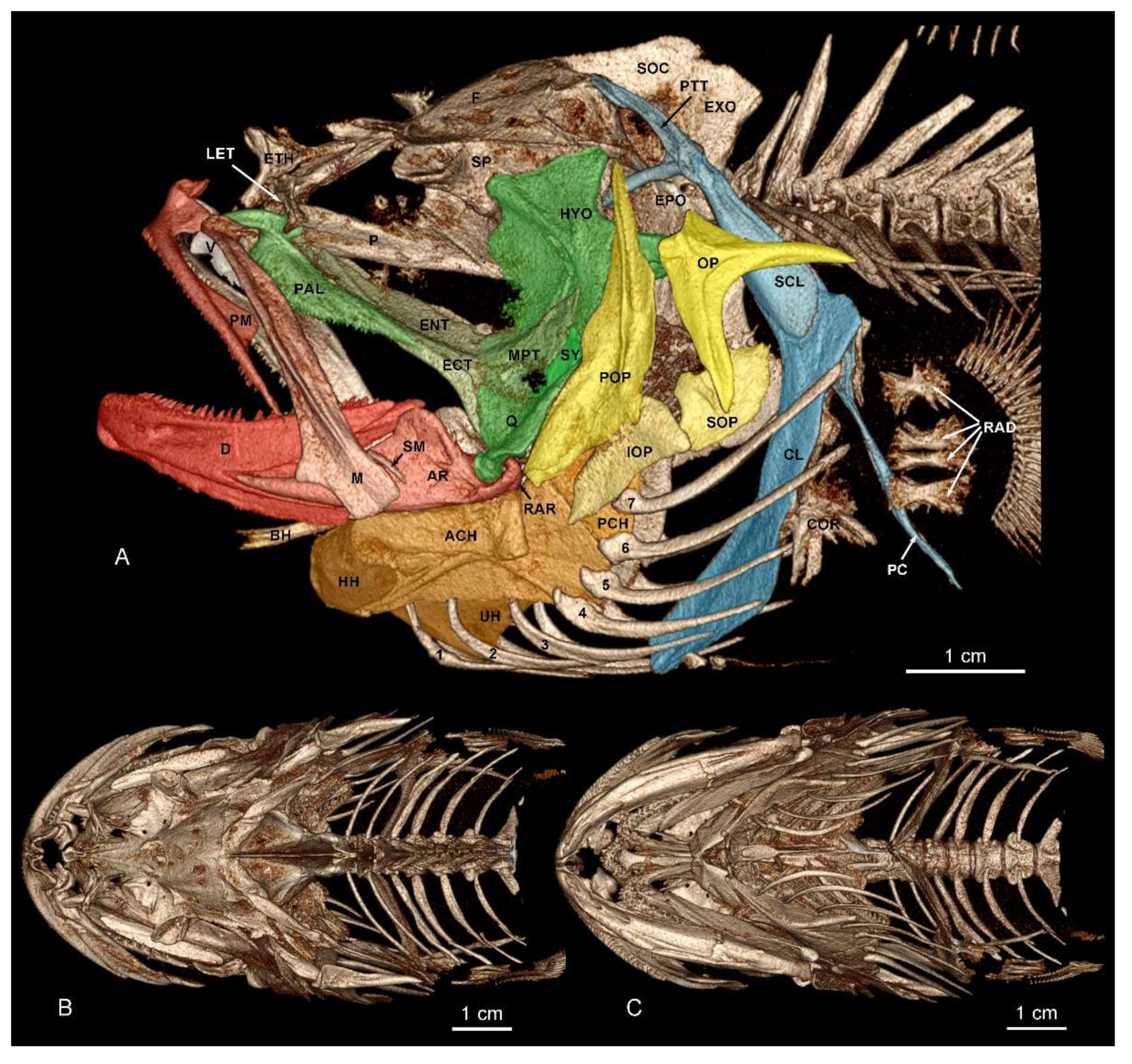
3. Results
Systematic Part
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Barnard, K.H. New records and descriptions of two new species of South African marine fishes. Ann. Mag. Nat. Hist. 1934, 10, 228–234. [Google Scholar] [CrossRef]
- Møller, P.R.; Knudsen, S.W.; Schwarzhans, W.; Nielsen, J.G. A new classification of viviparous brotulas (Bythitidae)—With family status for Dinematichthyidae—Based on molecular, morphological and fossil data. Mol. Phylogenetics Evol. 2016, 100, 391–408. [Google Scholar] [CrossRef] [PubMed]
- Hutton, F.W. Contributions to the ichthyology of New Zealand. Trans. N. Z. Inst. 1876, 8, 209–218. [Google Scholar]
- Hector, J. Notes on New Zealand ichthyology. Trans. N. Z. Inst. 1877, 9, 465–469. [Google Scholar]
- Nielsen, J.G.; Cohen, D.M. Melodichthys, a new genus with two new species of upper bathyal bythitids (Pisces, Ophidiiform). Cybium 1986, 10, 381–387. [Google Scholar]
- Paulin, C.D. Description of a new genus and two new species of bythitid fishes, and a redescription of Bidenichthys consobrinus (Hutton) from New Zealand. J. Nat. Hist. 1995, 29, 249–258. [Google Scholar] [CrossRef]
- Møller, P.R.; Nielsen, J.G. 98 Family Bythitidae. In The Fishes of New Zealand; Roberts, C.D., Stewart, A.L., Struthers, C.D., Eds.; Te Papa Press: Wellington, New Zealand, 2015; pp. 720–735. [Google Scholar]
- Nielsen, J.G.; Cohen, D.M.; Markle, D.F.; Robins, C.R. An Annotated and Illustrated Catalogue of Pearlfishes, Cusk-Eels, Brotulas and Other Ophidiiform Fishes Known to Date; FAO Species Catalogue. Ophidiiform Fishes of the World (Order Ophidiiformes). In FAO Fisheries Synopsis; FAO: Rome, Italy, 1999; p. 178. [Google Scholar]
- Møller, P.R.; Schwarzhans, W.; Nielsen, J.G. Review of the American Dinematichthyini (Teleostei, Bythitidae). Part I. Dinematichthys, Gunterichthys, Typhliasina and two new genera. Aqua J. Icht. Aquat. Biol. 2004, 8, 141–192. [Google Scholar]
- Sabaj, M.H. Standard Symbolic Codes for Institutional Resource Collections in Herpetology and Ichthyology; Version 7.1; American Society of Ichthyologists and Herpetologists: Washington, DC, USA, 2019; Available online: http://www.asih.org (accessed on 21 March 2019).
- Clouard, V.; Bonneville, A. Ages of seamounts, islands, and plateaus on the Pacific plate. In Plates, Plumes and Paradigms; Foulger, G.R., Natland, J.H., Presnall, D.C., Anderson, D.L., Eds.; Geological Society of America: Boulder, CO, USA, 2005; pp. 71–90. [Google Scholar]
- Clague, D.A.; Braga, J.C.; Bassi, D.; Fullagar, P.D.; Renema, W.; Webster, J.M. The maximum age of hawaiian terrestrial lineages: Geological constraints from Koko Seamount. J. Biogeogr. 2010, 37, 1022–1033. [Google Scholar] [CrossRef]
- Sharp, W.D.; Clague, D.A. 50-Ma initiation of Hawaiian-Emperor bend records major change in Pacific plate motion. Science 2006, 313, 1281–1284. [Google Scholar] [CrossRef] [Green Version]
- Clague, D.A.; Dalrymple, G.B.; Greene, H.G.; Wald, D.; Kono, M.; Kroenke, L.W. Bathymtery of the Emperor Seamounts. Init. Rep. Deep Sea Drill. Pro. 1980, 55, 845–849. [Google Scholar]
- Schwarzhans, W. Reconstruction of the fossil marine bony fish fauna (Teleostei) from the Eocene to Pleistocene of New Zealand by means of otoliths. Memorie della Società Italiana di Scienze Naturali e del Museo di Storia Naturale di Milano 2019, 46, 3–326. [Google Scholar]
- Howes, G.J. Notes on the anatomy and classification of ophidiiform fishes with particular reference to the abyssal genus Acanthonus Günther, 1878. Bull. Br. Mus. Nat. Hist. (Zool.) 1992, 58, 95–131. [Google Scholar]
- Carnevale, G.; Johnson, D. A Cretaceous Cusk-Eel (Teleostei, Ophidiiformes) from Italy and the Mesozoic Diversification of Percomorph Fishes. Copeia 2015, 103, 771–791. [Google Scholar] [CrossRef]
- Nolf, D. Etude monographique des otolithes des Ophidiiformes actuels et révision des espèces fossiles (Pisces, Teleostei). Mededelingen van de Werkgroep voor Tertiaire en Kwartaire Geologie 1980, 17, 71–195. [Google Scholar]
- Schwarzhans, W. Vergleichende morphologische Untersuchungen an rezenten und fossilen Otolithen der Ordnung Ophidiiformes. Berliner Geowissenschaftliche Abhandlungen (A) 1981, 32, 63–122. [Google Scholar]
- Nielsen, J.G.; Schwarzhans, W.; Cohen, D. 2012 Revision of Hastatobythites and Saccogaster (Teleostei, Bythitidae) with three new species and a new genus. Zootaxa 2012, 3579, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Gaemers, P.A.M. Fish otoliths from the Bassevelde Sand (Late Tongrian) of Ruisbroek, Belgium, and the stratigraphy of the Early Oligocene of Belgium. Meded. Werkgr. Tert. Kwart. Geol. 1984, 21, 13–57. [Google Scholar]
- Voigt, E. Über ein bemerkenswertes Vorkommen neuer Fischotolithen in einem Senongeschiebe von C€othen in Anhalt. Z. Geschiebeforschung 1926, 2, 172–187. [Google Scholar]
- Nolf, D. Les otolithes des téléostéens des Formations de Landen et de Heers (Paléocène de la Belgique). Geol. Palaeontol. 1978, 12, 223–234. [Google Scholar]
- Schwarzhans, W. Fish Otoliths from the New Zealand Tertiary; New Zealand Geological Survey: Wellington, New Zealand, 1984. [Google Scholar] [CrossRef]
- Nolf, D.; Dockery, D.T. Fish otoliths from the Matthews Landing Marl Member (Porters Creek Formation), Paleocene of Alabama. Miss. Geol. 1993, 14, 24–39. [Google Scholar]
- Lanckneus, J.; Nolf, D. Les otolithes des téléostéens redoniens de Bretagne (Néogène de l’Ouest de la France). Bulletin de l’Institut de Géologie du Bassin d’Aquitaine. Bulletin de l’Institut de Géologie du Bassin d’Aquitaine 1979, 25, 83–109. [Google Scholar]
- Eschmeyer, W.N.; Abe, T.; Nakano, S. Adelosebastes latens, a new genus and species of scorpionfish from the North Pacific Ocean (Pisces, Scorpaenidae). Jpn. Soc. Ichthyol. 1979, 30, 77–84. [Google Scholar]
- Iwamoto, T.; Okamoto, M. A new grenadier fish of the genus Lucigadus (Macrouridae, Gadiformes, Teleostei) from the Emperor Seamounts, Northwestern Pacific. Proc. Calif. Acad. Sci. 2015, 62, 369–380. [Google Scholar]
- Wilson, R.R.; Kaufmann, R.S. Seamount biota and biogeography. Geophys. Monogr. Ser. 1987, 43, 355–377. [Google Scholar]
- Stocks, K.I.; Hart, P.J.B. Biogeography and biodiversity of seamounts. In Seamounts: Ecology, Fisheries and Conservation; Pitcher, T.J., Morato, T., Hart, P.J.B., Clark, M.R., Haggan, N., Santos, R.S., Eds.; Fish and Aquatic Resources Series; Blackwell: Oxford, UK, 2007; pp. 255–281. [Google Scholar]
- Shank, T.M. Seamounts: Deep-ocean laboratories of faunal connectivity, evolution, and endemism. Oceanography 2010, 23, 109–122. [Google Scholar] [CrossRef] [Green Version]
- Hart, P.J.B.; Pearson, E. An application of the theory of island biogeography to fish speciation on seamounts. Mar. Ecol. Prog. Ser. 2011, 430, 281–288. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| B. okamotoi n. sp. | B. paxtoni | B. consobrinus | B. slartibartfasti | B. capensis | |||||
|---|---|---|---|---|---|---|---|---|---|
| HT | PT | HT AMS | HT and 2 Nontypes | 147 Nontypes | HT | HT, P T+ 5 Nontypes | ST | ST and 3 Nontypes | |
| ZMUC P2397-717, Male | ZMUC P2397-390, Female | E. 5264 | Mean and Range | Mean and Range | NMNZ P.30323 | Mean and Range | SAM 17995 | Mean and Range | |
| Ls (mm) | 187 | 169 | 74 | 112.7.3 (74–149) | 58.8 (19–90) | 93 | 87.6 (32–127) | 70 | 53.0 (47–70) |
| Meristic characters | |||||||||
| Dorsal fin rays | 69 | 70 | 69 | 72.7 (69–77) | 82.3 (78–87) | 70 | 69.5 (68–71) | 67 | 69.0 (66–72) |
| Caudal fin rays | 14 | 14 | 14 | 14 | 14 | 14 | 14 | 14 | 14 |
| Anal fin rays | 48 | 47 | 42 | 44.7 (42–46) | 46.3 (44–50) | 42 | 43.3 (42–45) | 36 | 38.8 (36–41) |
| Pectoral fin rays | 21 | 21 | 26 | 24.3 (23–26) | 26.5 (24–30) | 22 | 22.2 (21–23) | - | 21.0 (20–22) |
| Precaudal vertebrae | 12 | 12 | 13 | 13 | 14.2 (13–15) | 13 | 14.5 (13–16) | 13 | 13 |
| Caudal vertebrae | 33 | 34 | 33 | 33.7 (33–34) | 31.5 (30–33) | 33 | 31.2 (29–33) | 28 | 28.3 (27–29) |
| Total vertebrae | 45 | 46 | 46 | 46.7 (46–47) | 45.7 (45–47) | 46 | 45.7 (45–46) | 41 | 41.3 (40–42) |
| Rakers on anterior gill arch | 27 | 22 | 20 | 19.3 (18–20) | 18.6 (18–20) | 20 | 21.7 (20–26) | 13 | 15.5 (13–17) |
| Long rakers on anterior gill arch | 17 | 13 | 11 | 11.0 (10–12) | 6.2 (5–8) | 15 | 14.9 (13–17) | 4 | 4.8 (4–5) |
| Pseudobranchial filaments | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 0 | 0 |
| Anterior dorsal fin ray above vertebra no. (D/V) | 5 | 5 | 6 | 6 | 5.1 (5–6) | 6 | 5.7 (5–6) | 6 | 5.3 (5–6) |
| Anterior anal fin ray below dorsal fin ray no. (D/A) | 29 | 27 | 35 | 34.3 (33–35) | 38.4 (34–42) | 28 | 28.7 (26–30) | 33 | 34.5 (33–36) |
| Anterior anal fin ray below vertebra no. (V/A) | 19 | 18 | 21 | 21 | 21.0 (20–22) | 19 | 19.5 (19–20) | 20 | 20.3 (20–21) |
| Palatine teeth rows | 6 | 4 | 3 | 2.7 (2–3) | 2.7 (2–3) | 2 | 2.1 (2–3) | - | 2 |
| Vomer teeth rows | 4 | 4 | - | 3 | 2.5 (2–3) | 3 | 2.6 (2–4) | - | 2.3 (2–3) |
| Premaxillary teeth rows | 7 | 4 | - | 4–7 | 3.2 (3–4) | 4 | 3.6 (3–5) | 2 | 3 |
| Dentary teeth rows | 5 | 4 | - | 4–6 | 3.4 (3–4) | 4 | 3.7 (3–4) | 2 | 3 |
| Scale rows above anal fin origin | 68 | 67 | - | 39–41 | 28.8 (27–31) | 38 | 36 (33–40) | - | 19.3 (17–23) |
| Anterior mandibular head pores | 2 | 2 | 3 | 3 | 2 | 3 | 2.9 (2–3) | - | 2 |
| Posterior mandibular head pores | 0 | 0 | 3 | 3 | 0 | 3 | 3 | - | 0 |
| Supraorbital head pores | 1 | 1 | 1 | 1 | 1 | 1 | 1 | - | 1 |
| First lateral line head pore | 1 | 1 | 1 | 1 | 0.7 (0–1) | 0 | 0 | - | 1 |
| Lower preopercular head pores | 0 | 0 | 2 | 2 | 0 | 2 | 2 | - | 0 |
| Anterior infraorbital head pores | 3 | 3 | 2 | 2.7 (2–3) | 3 | 3 | 3 | - | 2.3 (2–3) |
| Posterior infraorbital head pores | 0 | 0 | 0 | 0 | 0 | 0 | 0 | - | 2 |
| Morphometric characters in % of Ls | |||||||||
| Head length | 30.1 | 29.6 | 31.7 | 30.3 (28.5–31.7) | 28.5 (26.7–30.3) | 29.9 | 30.5 (28.7–33.7) | - | 30.4 (29.4–32.1) |
| Head width | 20.9 | 19.2 | - | 19.1 (18.6–19.6) | 15.4 (13.3–17.7) | 17.6 | 18.1 (15.3–21.3) | - | 16.1 (15.1–17.3) |
| Head height | 25.6 | 24.8 | - | 22.6 (20.6–24.6) | 19.5 (17.9–20.9) | 21.8 | 22.3 (19.1–27.1) | - | 19.6 (18.5–18.0) |
| Snout length | 7.6 | 7.1 | 6.1 | 6.9 (6.1–7.5) | 6.2 (5.8–6.9) | 6.7 | 6.6 (5.8–7.2) | - | 7.3 (7.2–7.4) |
| Upper jaw length | 17.5 | 17.9 | 15.4 | 15.8 (15.3–16.6) | 13.5 (12.9–14.7) | 15.2 | 16.1 (14.9–19.0) | - | 15.7 (15.1–16.8) |
| Maxillary height | 3.7 | 3.7 | - | 4.2 (3.9–4.4) | 3.5 (3.3–3.6) | 3.7 | 3.9 (2.9–4.5) | - | 4.6 (4.3–5.0) |
| Diameter of pigmented eye | 4.3 | 4.1 | 5.0 | 4.8 (4.4–5.0) | 3.9 (3.6–4.4) | 4.7 | 4.7 (4.0–5.4) | - | 4.4 (4.3–4.5) |
| Interorbital width | 8.5 | 8.6 | 7.2 | 7.8 (7.2–8.8) | 5.2 (4.6–5.8) | 6.6 | 7.0 (6.6–7.4) | - | 5.9 (5.8–6.2) |
| Postorbital length | 19.6 | 19.6 | 20.7 | 19.3 (18.0–20.7) | 19.3 (18.3–20.4) | 20.9 | 20.5 (18.9–23.5) | - | 20.1 (19.3–20.6) |
| Preanal length | 62.0 | 60.9 | 64.0 | 65.5 (64.0–66.2) | 59.7 (56.0–62.3) | 57.7 | 59.8 (56.6–64.0) | - | 64.0 (62.3–65.1) |
| Predorsal length | 32.0 | 31.8 | 33.8 | 33.3 (31.5–34.6) | 30.9 (29.8–33.1) | 31.9 | 33.1 (31.1–36.7) | - | 31.2 (30.2–32.5) |
| Body depth at origin of anal fin | 20.0 | 18.3 | 18.0 | 20.1 (17.9–20.1) | 17.7 (16.8–19.9) | 20.4 | 19.0 (17.0–20.8) | - | 15.6 (15.2–16.4) |
| Pectoral fin length | 15.2 | 16.1 | 19.8 | 18.9 (17.4–19.8) | 18.1 (16.1–19.8) | 18.1 | 18.4 (17.0–20.0) | - | 19.0 (18.5–19.4) |
| Pelvic fin length | 12.1 | 13.4 | 17.9 | 16.9 (15.7–17.9) | 19.4 (17.6–20.7) | 16.9 | 17.5 (14.4–19.5) | - | 20.5 (19.9–21.0) |
| Base of ventral fin to anal fin origin | 39.6 | 37.7 | 42.0 | 44.4 (42.0–46.9) | 38.1 (35.7–40.8) | 34.0 | 36.4 (34.0–38.6) | - | 41.6 (40.1–42.4) |
| Caudal fin length | 9.0 | 7.1 | - | - | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Møller, P.R.; Schwarzhans, W.W.; Lauridsen, H.; Nielsen, J.G. Bidenichthys okamotoi, a New Species of the Bythitidae (Ophidiiformes, Teleostei) from the Koko Seamount, Central North Pacific. J. Mar. Sci. Eng. 2021, 9, 1399. https://doi.org/10.3390/jmse9121399
Møller PR, Schwarzhans WW, Lauridsen H, Nielsen JG. Bidenichthys okamotoi, a New Species of the Bythitidae (Ophidiiformes, Teleostei) from the Koko Seamount, Central North Pacific. Journal of Marine Science and Engineering. 2021; 9(12):1399. https://doi.org/10.3390/jmse9121399
Chicago/Turabian StyleMøller, Peter R., Werner W. Schwarzhans, Henrik Lauridsen, and Jørgen G. Nielsen. 2021. "Bidenichthys okamotoi, a New Species of the Bythitidae (Ophidiiformes, Teleostei) from the Koko Seamount, Central North Pacific" Journal of Marine Science and Engineering 9, no. 12: 1399. https://doi.org/10.3390/jmse9121399