Copepod Community Structure in Pre- and Post- Winter Conditions in the Southern Adriatic Sea (NE Mediterranean)


Abstract
:1. Introduction
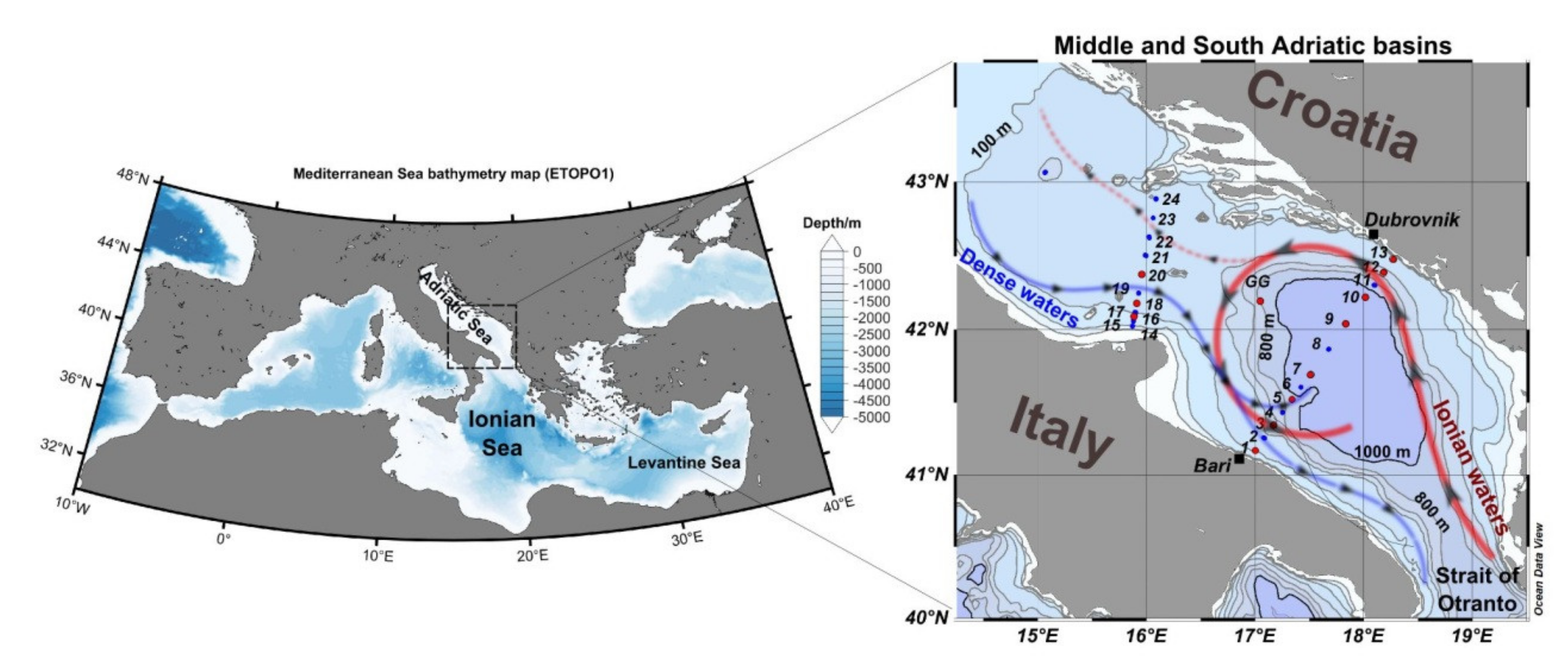
2. Materials and Methods
3. Results
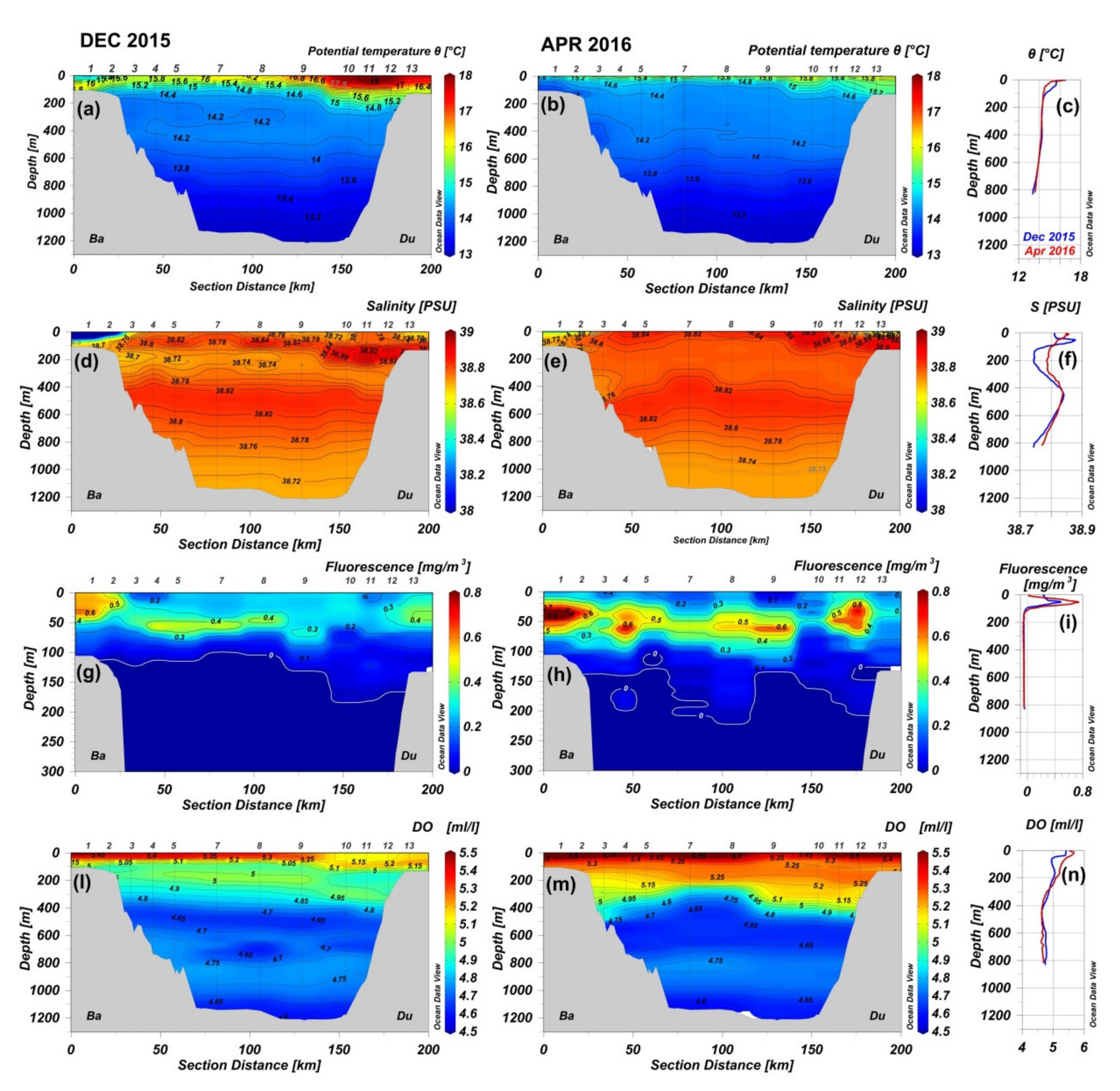
3.1. Environmental Conditions
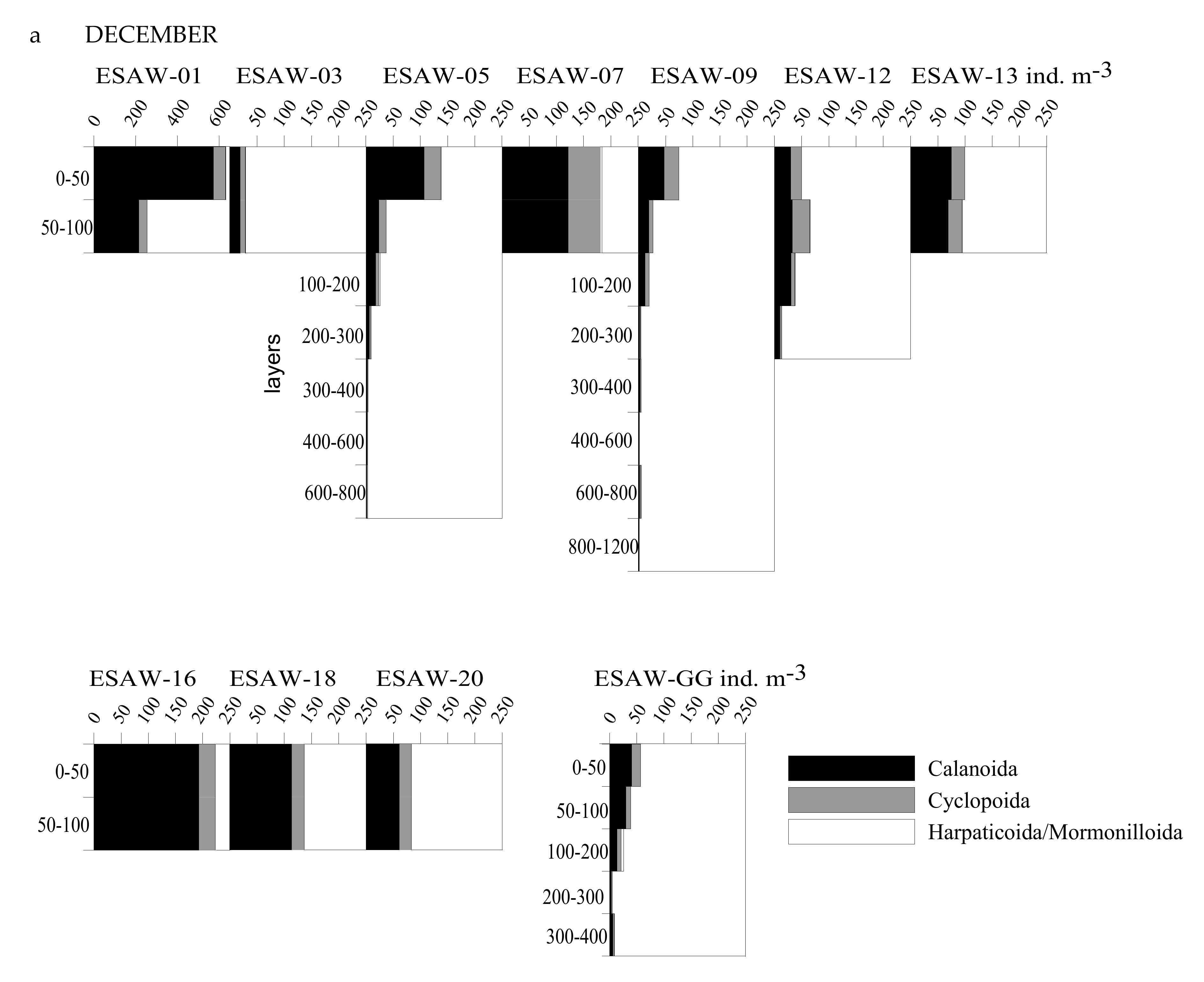
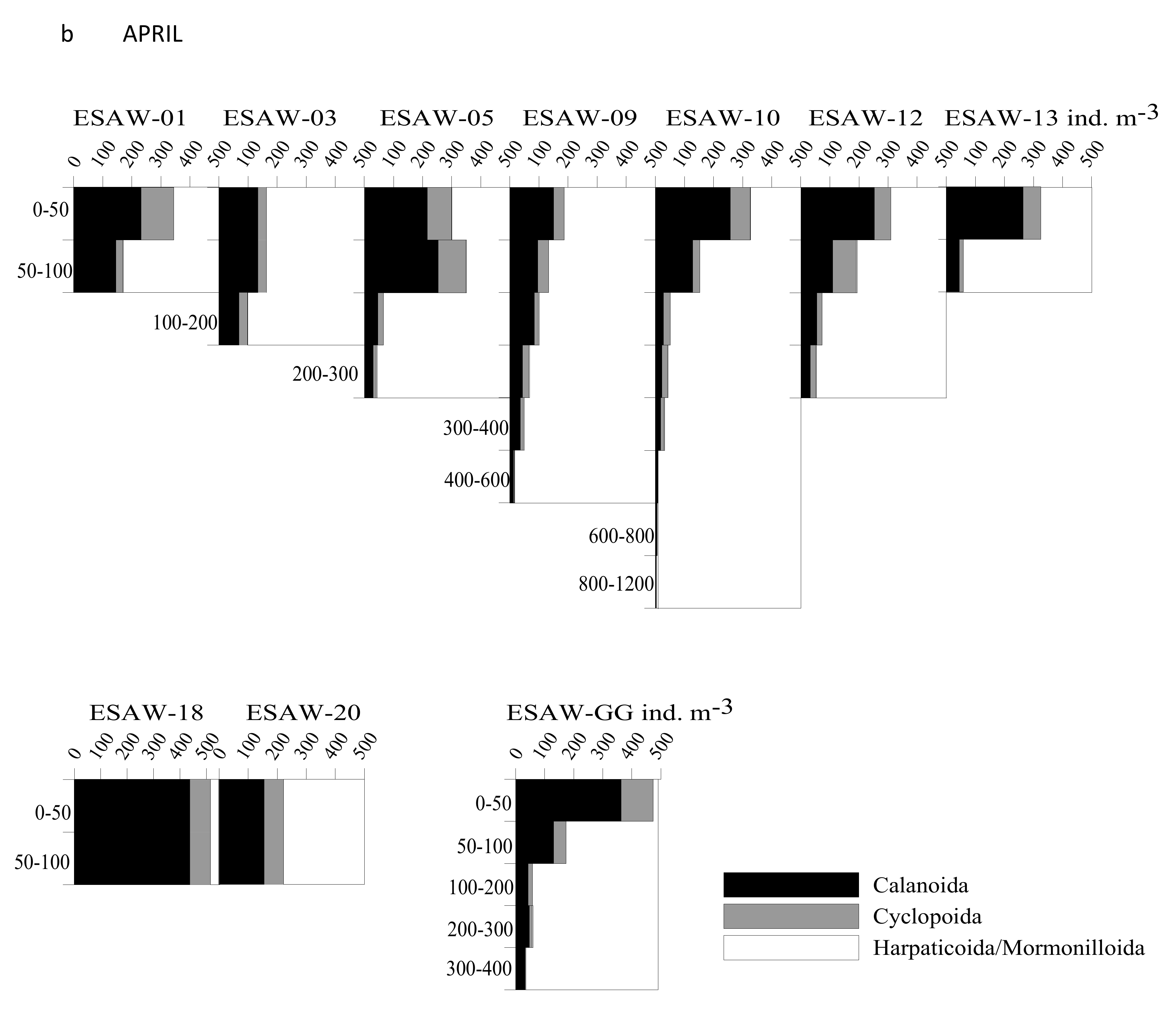
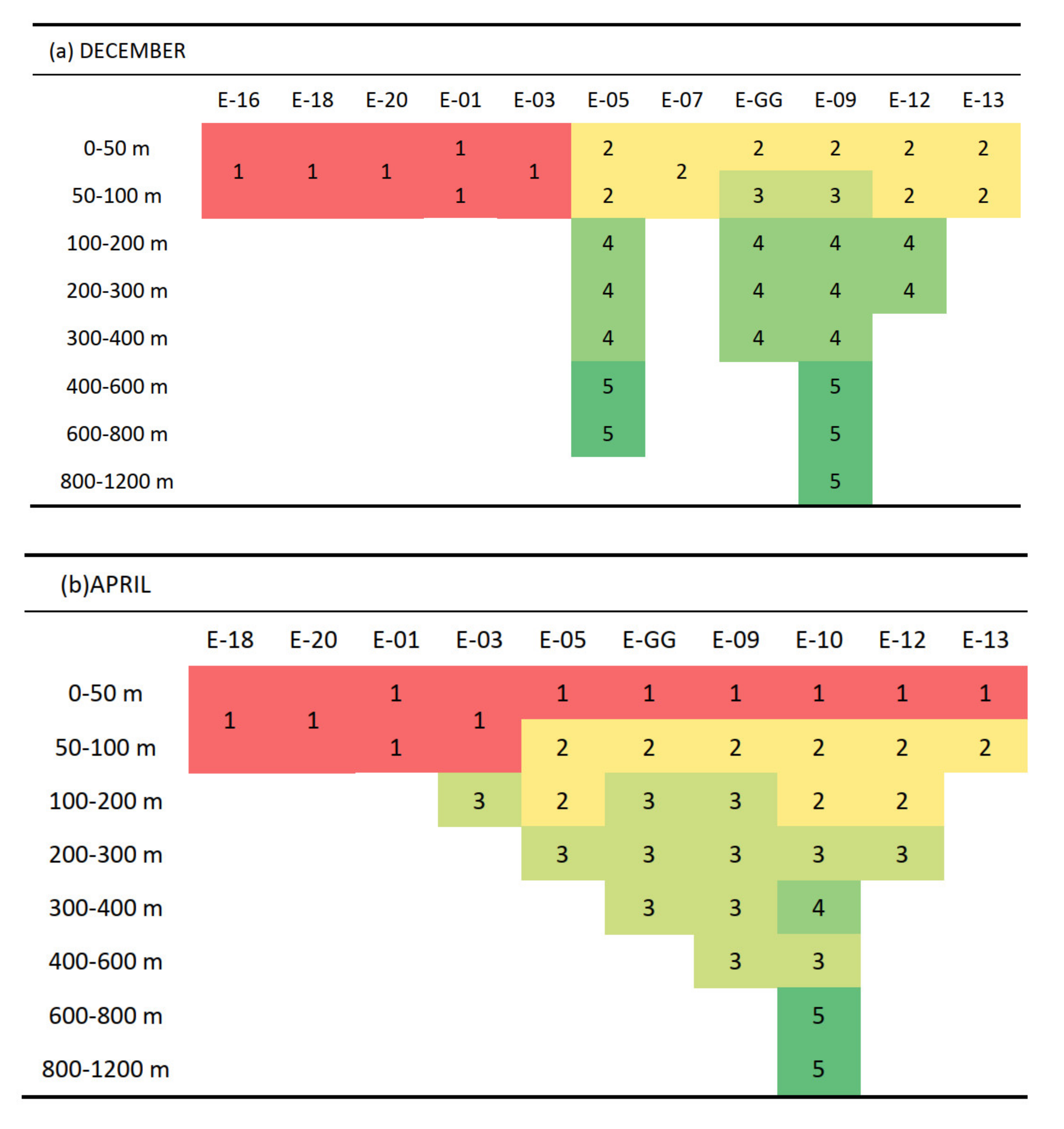
3.2. Vertical and Horizontal Distribution of Copepod Abundance
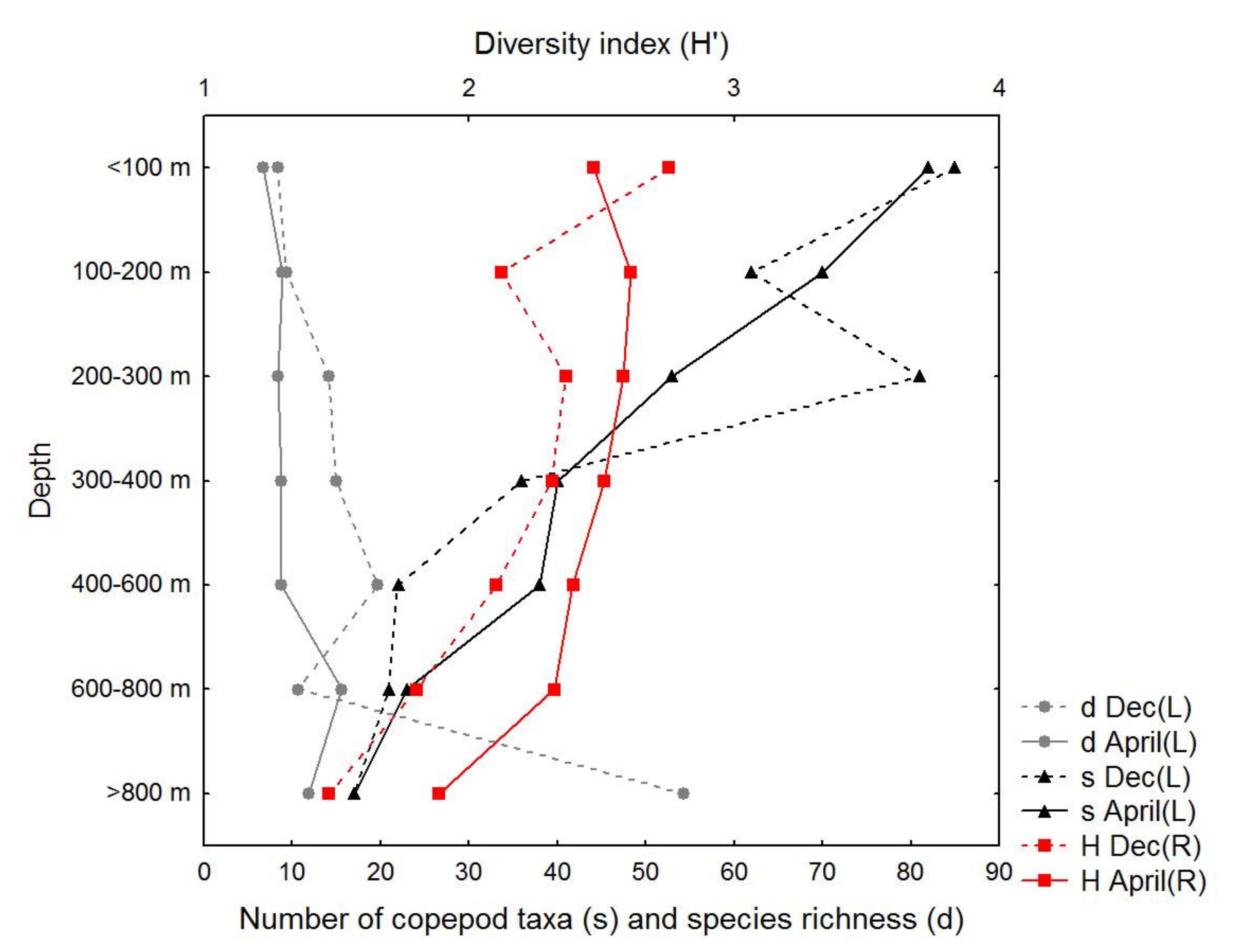
3.3. Copepod Composition and Diversity
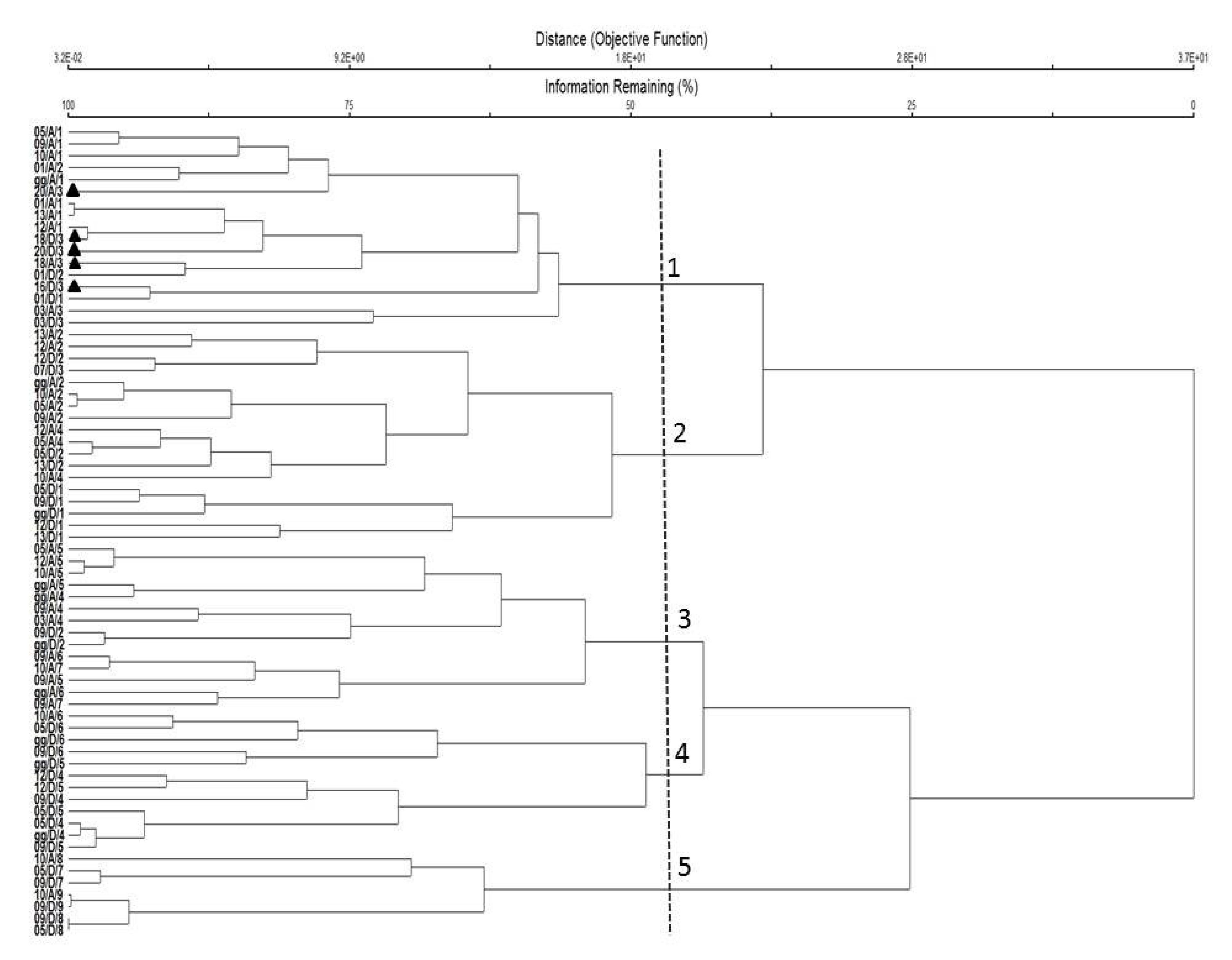
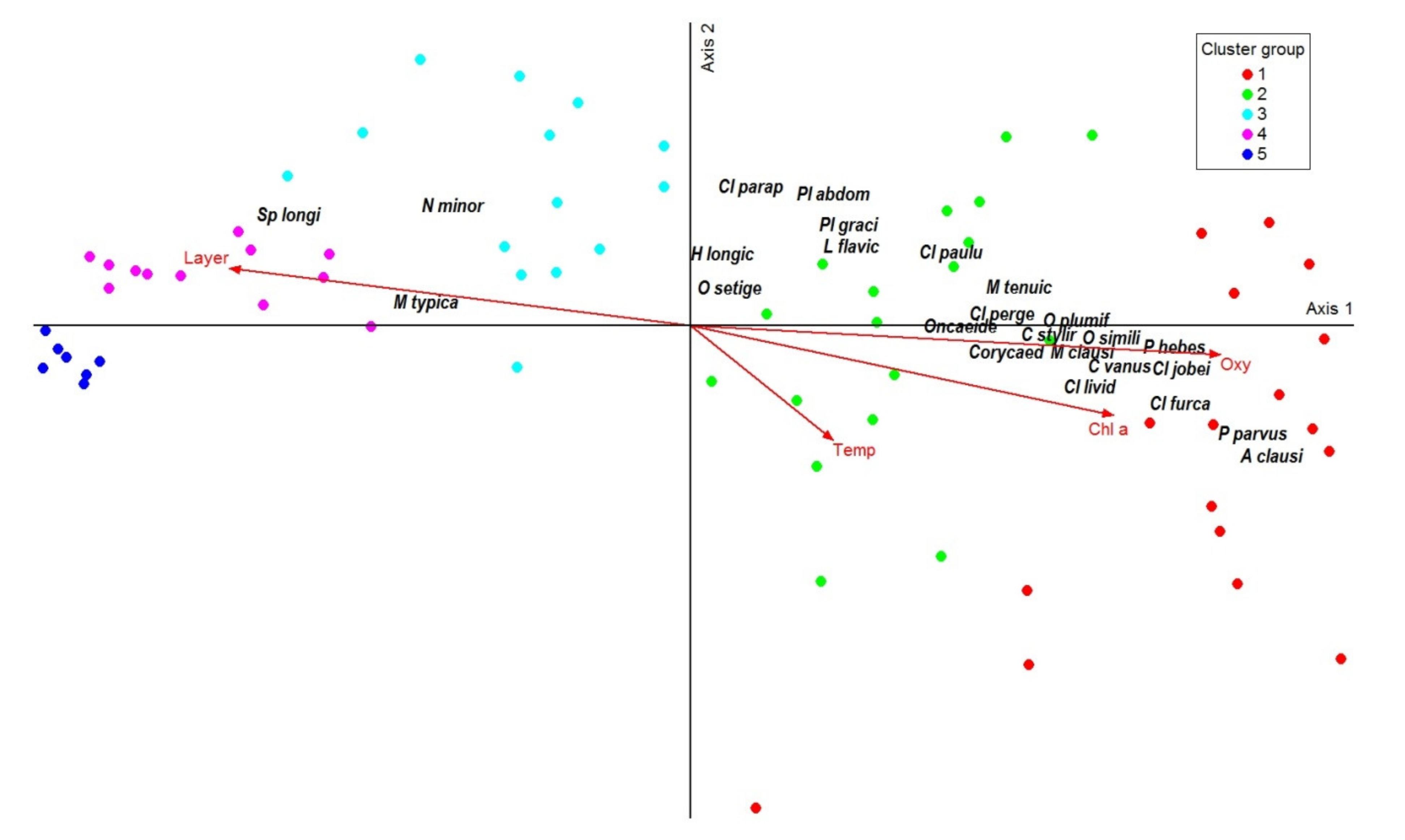
3.4. Copepod Community Structure
4. Discussion
4.1. Copepod Abundance, Species Composition, and Diversity
4.2. Pattern of Copepod Assemblages in Relation to Environmental Parameters
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zore, M. On the gradient currents in the Adriatic Sea. Acta Adriat. 1956, 8, 1–38. [Google Scholar]
- Artegiani, A.; Bregant, D.; Paschini, E.; Pinardi, N.; Raicich, F.; Russo, A. The Adriatic Sea General Circulation. Part II: Baroclinic Circulation Structure. J. Phys. Ocean 1997, 27, 1515–1532. [Google Scholar] [CrossRef]
- Gačić, M.; Marullo, S.; Santoleri, R.; Bergamasco, A. Analysis of the seasonal and interannual variability of the sea surface temperature field in the Adriatic Sea from AVHRR data (1984-1992). J. Geophys. Res. Oceans 1997, 102, 22937–22946. [Google Scholar] [CrossRef]
- Cushman-Roisin, B.; Malačić, V.; Gačić, M. Tides, Seiches and Low-Frequency Oscillations. In Physical Oceanography of the Adriatic Sea; Springer: Dordrecht, The Netherlands, 2001. [Google Scholar]
- Grbec, B.; Dulcic, J.; Morovic, M. Long-term changes in landings of small pelagic fish in the eastern Adriatic-possible influence of climate oscillations over the Northern Hemisphere. Clim. Res. 2002, 20, 241–252. [Google Scholar] [CrossRef]
- Cozzi, S.; Giani, M. River water and nutrient discharges in the Northern Adriatic Sea: Current importance and long term changes. Cont. Shelf Res. 2011, 31, 1881–1893. [Google Scholar] [CrossRef]
- Zoppini, A.; Ademollo, N.; Bensi, M.; Berto, D.; Bongiorni, L.; Campanelli, A.; Casentini, B.; Patrolecco, L.; Amalfitano, S. Impact of a river flood on marine water quality and planktonic microbial communities. Estuar. Coast. Shelf Sci. 2019, 224, 62–72. [Google Scholar] [CrossRef]
- Buljan, M.; Zore-Armanda, M. Hydrographic properties of the Adriatic Sea in the period from 1965 through 1970. Acta Adriat. 1979, 20, 1–368. [Google Scholar]
- Faganeli, J.; Gačić, M.; Malej, A.; Smodlaka, N. Pelagic organic matter in the Adriatic Sea in relation to winter hydrographic conditions. J. Plankton Res. 1989, 11, 1129–1141. [Google Scholar] [CrossRef]
- Gačić, M.; Borzelli, G.L.E.; Civitarese, G.; Cardin, V.; Yari, S. Can internal processes sustain reversals of the ocean upper circulation? The Ionian Sea example: INTERNAL PROCESSES AND UPPER CIRCULATION. Geophys. Res. Lett. 2010, 37. [Google Scholar] [CrossRef]
- Rubino, A.; Gačić, M.; Bensi, M.; Kovačević, V.; Malačič, V.; Menna, M.; Negretti, M.E.; Sommeria, J.; Zanchettin, D.; Barreto, R.V.; et al. Experimental evidence of long-term oceanic circulation reversals without wind influence in the North Ionian Sea. Sci. Rep. 2020, 10, 1905. [Google Scholar] [CrossRef]
- Gačić, M.; Civitarese, G.; Miserocchi, S.; Cardin, V.; Crise, A.; Mauri, E. The open-ocean convection in the Southern Adriatic: A controlling mechanism of the spring phytoplankton bloom. Cont. Shelf Res. 2002, 22, 1897–1908. [Google Scholar] [CrossRef]
- Vilibić, I.; Matijević, S.; Šepić, J.; Kušpilić, G. Changes in the Adriatic oceanographic properties induced by the Eastern Mediterranean Transient. Biogeosciences 2012, 9, 2085–2097. [Google Scholar] [CrossRef] [Green Version]
- Batistić, M.; Viličić, D.; Kovačević, V.; Jasprica, N.; Garić, R.; Lavigne, H.; Carić, M. Occurrence of winter phytoplankton bloom in the open southern Adriatic: Relationship with hydroclimatic events in the Eastern Mediterranean. Cont. Shelf Res. 2019, 174, 12–25. [Google Scholar] [CrossRef]
- Batistić, M.; Jasprica, N.; Carić, M.; Čalić, M.; Kovačević, V.; Garić, R.; Njire, J.; Mikuš, J.; Bobanović-Ćolić, S. Biological evidence of a winter convection event in the South Adriatic: A phytoplankton maximum in the aphotic zone. Cont. Shelf Res. 2012, 44, 57–71. [Google Scholar] [CrossRef]
- Batistić, M.; Garić, R.; Molinero, J. Interannual variations in Adriatic Sea zooplankton mirror shifts in circulation regimes in the Ionian Sea. Clim. Res. 2014, 61, 231–240. [Google Scholar] [CrossRef]
- Civitarese, G.; Gačić, M.; Lipizer, M.; Eusebi Borzelli, G.L. On the impact of the Bimodal Oscillating System (BiOS) on the biogeochemistry and biology of the Adriatic and Ionian Seas (Eastern Mediterranean). Biogeosciences 2010, 7, 3987–3997. [Google Scholar] [CrossRef] [Green Version]
- Čalić, M.; Ljubimir, S.; Bosak, S.; Car, A. First records of two planktonic Indo-Pacific diatoms: Chaetoceros bacteriastroides and C. pseudosymmetricus in the Adriatic Sea. Oceanologia 2018, 60, 101–105. [Google Scholar] [CrossRef]
- Hure, M.; Mihanović, H.; Lučić, D.; Ljubešić, Z.; Kružić, P. Mesozooplankton spatial distribution and community structure in the South Adriatic Sea during two winters (2015, 2016). Mar. Ecol. 2018, 39, e12488. [Google Scholar] [CrossRef]
- Njire, J.; Batistić, M.; Kovačević, V.; Garić, R.; Bensi, M. Tintinnid Ciliate Communities in Pre- and Post-Winter Conditions in the Southern Adriatic Sea (NE Mediterranean). Water 2019, 11, 2329. [Google Scholar] [CrossRef] [Green Version]
- Kiørboe, T. What makes pelagic copepods so successful? J. Plankton Res. 2011, 33, 677–685. [Google Scholar] [CrossRef] [Green Version]
- Siokou-Frangou, I.; Christou, E.D.; Fragopoulu, N.; Mazzocchi, M.G. Mesozooplankton distribution from Sicily to Cyprus (Eastern Mediterranean): II. Copepod assemblages. Oceanol. Acta 1997, 20, 537–548. [Google Scholar]
- Siokou-Frangou, I.; Christaki, U.; Mazzocchi, M.G.; Montresor, M.; Apos, M.R.; Alcalá, M.; Vaqué, D.; Zingone, A. Plankton in the open Mediterranean Sea: A review. Biogeosciences 2010, 7, 1543–1586. [Google Scholar] [CrossRef] [Green Version]
- Siokou, I.; Frangoulis, C.; Grigoratou, Μ.; Pantazi, M. Zooplankton community dynamics in the N. Aegean front (E. Mediterranean) in the winter spring period. Mediterr. Mar. Sci. 2014, 15, 706. [Google Scholar] [CrossRef] [Green Version]
- Siokou, I.; Zervoudaki, S.; Velaoras, D.; Theocharis, A.; Christou, E.D.; Protopapa, M.; Pantazi, M. Mesozooplankton vertical patterns along an east-west transect in the oligotrophic Mediterranean sea during early summer. Deep Sea Res. Part II Top. Stud. Oceanogr. 2019, 164, 170–189. [Google Scholar] [CrossRef]
- Mazzocchi, M.G. Spring mesozooplankton communities in the epipelagic Ionian Sea in relation to the Eastern Mediterranean Transient. J. Geophys. Res. 2003, 108, 8114. [Google Scholar] [CrossRef] [Green Version]
- Costalago, D.; Garrido, S.; Palomera, I. Comparison of the feeding apparatus and diet of European sardines Sardina pilchardus of Atlantic and Mediterranean waters: Ecological implications. J. Fish Biol. 2015, 86, 1348–1362. [Google Scholar] [CrossRef] [PubMed]
- Hunter, J.R. Feeding ecology and predation of marine fish larvae. In Marine Fish Larvae. Morphology, Ecology and Relation to Fisheries; University Washington Press: Seattle, WA, USA, 1981; pp. 33–77. [Google Scholar]
- Turner, J.T. The Importance of Small Planktonic Copepods and Their Roles in Pelagic Marine Food. Zool. Stud. 2004, 43, 255–266. [Google Scholar]
- Jónasdóttir, S.H.; Visser, A.W.; Richardson, K.; Heath, M.R. Seasonal copepod lipid pump promotes carbon sequestration in the deep North Atlantic. Proc. Natl. Acad. Sci. USA 2015, 112, 12122–12126. [Google Scholar] [CrossRef] [Green Version]
- Turner, J.T. Zooplankton fecal pellets, marine snow, phytodetritus and the ocean’s biological pump. Prog. Oceanogr. 2015, 130, 205–248. [Google Scholar] [CrossRef]
- Di Carlo, B.S.; Ianora, A.; Fresi, E.; Hure, J. Vertical zonation patterns for Mediterranean copepods from the surface to 3000 m at a fixed station in the Tyrrhenian Sea. J. Plankton Res. 1984, 6, 1031–1056. [Google Scholar] [CrossRef]
- Gaard, E. The zooplankton community structure in relation to its biological and physical environment on the Faroe Shelf, 1989–1997. J. Plankton Res. 1999, 21, 1133–1152. [Google Scholar] [CrossRef]
- Beaugrand, G.; Ibanez, F. Monitoring marine plankton ecosystems. II: Long-term changes in North Sea calanoid copepods in relation to hydro-climatic variability. Mar. Ecol. Prog. Ser. 2004, 284, 35–47. [Google Scholar] [CrossRef]
- Hure, J. Distribution annuelle vertical du zooplankton sur une station de l’Adriatique méridionale. Acta Adriat. 1955, 7, 1–72. [Google Scholar]
- Hure, J. Dnevna migracija i sezonska vertikalna raspodjela zooplanktona dubljeg mora. Acta Adriat. 1961, 9, 1–59. [Google Scholar]
- Hure, J.; Scotto di Carlo, B. Comparazione tra lo zooplankton del Golfo di Napoli a dell Adriatico meridionale presso Dubrovnik I. Copepoda. Pubblicatione Stn. Zool. Napoli 1968, 36, 21–102. [Google Scholar]
- Hure, J.; Scotto di Carlo, B. Copepodi pelagici dell Adriatico settentrionale nel periodo gennaio—Dicembre 1965. Pubblicatione Stn. Zool. Napoli 1969, 37, 173–195. [Google Scholar]
- Hure, J.; Scotto di Carlo, B. Ripartizione quantitative e distribuzione verticale dei Copepodi pelagici di profondita su una stazione nel Mar Tirreno ed una nell’Adriatico Meridionale. Pubblicatione Stn. Zool. Napoli 1969, 37, 51–83. [Google Scholar]
- Hure, J.; Scotto di Carlo, B. Diurnal vertical migration of some deep water copepods in the Southern Adriatic (East Mediterranean). Pubblicatione Stn. Zool. Napoli 1969, 37, 581–598. [Google Scholar]
- Hure, J.; Scotto di Carlo, B. New patterns of diurnal vertical migration of some deep-water copepods in the Tyrrhenian and Adriatic Seas. Mar. Biol. 1974, 28, 179–184. [Google Scholar]
- Vučetić, T. Les principals masses d’ eau en Adriatique et leur influence sur les communautés pélagiques. Jounées Etud. Planctonol. C.I.E.S.M. 1970, 20, 105–114. [Google Scholar]
- Vučetić, T. Zooplankton and circulation pattern of the water masses in the Adriatic. Neth. J. Sea Res. 1973, 7, 112–121. [Google Scholar]
- Hure, J.; Kršinić, F. Planktonic copepods of the Adriatic sea. Nat. Croat. 1998, 7, 1–135. [Google Scholar]
- Hure, J.; Ianora, A.; di Carlo, B.S. Spatial and temporal distribution of copepod communities in the Adriatic Sea. J. Plankton Res. 1980, 2, 295–316. [Google Scholar] [CrossRef]
- Miloslavić, M.; Lučić, D.; Njire, J.; Gangai, B.; Onofri, I.; Garić, R.; Žarić, M.; Miri Osmani, F.; Pestorić, B.; Nikleka, E.; et al. Zooplankton composition and distribution across coastal and offshore waters off Albania (Southern Adriatic) in late spring. Acta Adriat. 2012, 53, 155–320. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. Primer v5: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2001. [Google Scholar]
- Margalef, R. Perspectives in Ecology Theory; University Chicago Press: Chicago, IL, USA, 1968. [Google Scholar]
- Shannon, C.E.; Wiener, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1963. [Google Scholar]
- McCune, B.; Mefford, M.J. PC-ORD Multivariate Analysis of Ecological Data; MjM Software: Gleneden Beach, OR, USA, 2006. [Google Scholar]
- McCune, B.; Grace, J.B. Analysis of Ecological Communities; MjM Software: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology, Developments in Environmental Modelling; Elsevier: Amsterdam, The Netherlands, 1998; Volume 20. [Google Scholar]
- Dufrene, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar]
- Mather, P.M. Computational Methods of Multivariate Analysis in Physical Geography; J. Wiley and Sons: London, UK, 1976. [Google Scholar]
- Kokkini, Z.; Mauri, E.; Gerin, R.; Poulain, P.M.; Simoncelli, S.; Notarstefano, G. On the salinity structure in the South Adriatic as derived from float and glider observations in 2013–2016. Deep Sea Res. Part II Top. Stud. Oceanogr. 2020, 171, 104625. [Google Scholar] [CrossRef]
- Calbet, A. Annual Zooplankton Succession in Coastal NW Mediterranean Waters: The Importance of the Smaller Size Fractions. J. Plankton Res. 2001, 23, 319–331. [Google Scholar] [CrossRef]
- Zervoudaki, S.; Nielsen, T.G.; Christou, E.D.; Siokou-Frangou, I. Zooplankton distribution and diversity in a frontal area of the Aegean Sea. Mar. Biol. Res. 2006, 2, 149–168. [Google Scholar] [CrossRef]
- Webber, M.K.; Roff, J.C. Annual biomass and production of the oceanic copepod community off Discovery Bay, Jamaica. Mar. Biol. 1995, 123, 481–495. [Google Scholar] [CrossRef]
- Ribera d’Alcalà, M.; Conversano, F.; Corato, F.; Licandro, P.; Mangoni, O.; Marino, D.; Mazzocchi, M.G.; Modigh, M.; Montresor, M.; Nardella, M.; et al. Seasonal patterns in plankton communities in a pluriannual time series at a coastal Mediterranean site (Gulf of Naples): An attempt to discern recurrences and trends. Sci. Mar. 2004, 68, 65–83. [Google Scholar] [CrossRef] [Green Version]
- Weikert, H.; Trinkaus, S. Vertical mesozooplankton abundance and distribution in the deep Eastern Mediterranean Sea SE of Crete. J. Plankton Res. 1990, 12, 601–628. [Google Scholar] [CrossRef]
- Brugnano, C.; Bergamasco, A.; Granata, A.; Guglielmo, L.; Zagami, G. Spatial distribution and community structure of copepods in a central Mediterranean key region (Egadi Islands—Sicily Channel). J. Mar. Syst. 2010, 81, 312–322. [Google Scholar] [CrossRef]
- Fernández de Puelles, M.L.; Alemany, F.; Jansá, J. Zooplankton time-series in the Balearic Sea (Western Mediterranean): Variability during the decade 1994–2003. Prog. Oceanogr. 2007, 74, 329–354. [Google Scholar] [CrossRef]
- Siokou, I.; Zervoudaki, S.; Christou, E.D. Mesozooplankton community distribution down to 1000 m along a gradient of oligotrophy in the Eastern Mediterranean Sea (Aegean Sea). J. Plankton Res. 2013, 35, 1313–1330. [Google Scholar] [CrossRef] [Green Version]
- Brugnano, C.; Granata, A.; Guglielmo, L.; Zagami, G. Spring diel vertical distribution of copepod abundances and diversity in the open Central Tyrrhenian Sea (Western Mediterranean). J. Mar. Syst. 2012, 105, 207–220. [Google Scholar] [CrossRef]
- Gaudy, R.; Youssara, F.; Diaz, F.; Raimbault, P. Biomass, metabolism and nutrition of zooplankton in the Gulf of Lions (NW Mediterranean). Oceanol. Acta 2003, 26, 357–372. [Google Scholar] [CrossRef] [Green Version]
- Peralba, À.; Mazzocchi, M.G. Vertical and seasonal distribution of eight Clausocalanus species (Copepoda: Calanoida) in oligotrophic waters. ICES J. Mar. Sci. 2004, 61, 645–653. [Google Scholar] [CrossRef]
- Nowaczyk, A.; Carlotti, F.; Thibault-Botha, D.; Pagano, M. Distribution of epipelagic metazooplankton across the Mediterranean Sea during the summer BOUM cruise. Biogeosciences 2011, 8, 2159–2177. [Google Scholar] [CrossRef] [Green Version]
- Pasternak, A.; Wassmann, P.; Riser, C.W. Does mesozooplankton respond to episodic P inputs in the Eastern Mediterranean? Deep Sea Res. Part II Top. Stud. Oceanogr. 2005, 52, 2975–2989. [Google Scholar] [CrossRef]
- Riandey, V.; Champalbert, G.; Carlotti, F.; Taupier-Letage, I.; Thibault-Botha, D. Zooplankton distribution related to the hydrodynamic features in the Algerian Basin (western Mediterranean Sea) in summer 1997. Deep Sea Res. Part I Oceanogr. Res. Pap. 2005, 52, 2029–2048. [Google Scholar] [CrossRef]
- Pancuccipapadopoulou, M.; Siokoufrangou, I.; Theocharis, A.; Georgopoulos, D. Zooplankton vertical-distribution in relation to the hydrology in the nw levantine and the se aegean seas (spring 1986). Oceanol. Acta 1992, 15, 365–381. [Google Scholar]
- Koppelmann, R.; Weikert, H. Spatial and temporal distribution patterns of deep-sea mesozooplankton in the eastern Mediterranean? indications of a climatically induced shift? Mar. Ecol. 2007, 28, 259–275. [Google Scholar] [CrossRef]
- Manca, B.B. Evolution of dynamics in the eastern Mediterranean affecting water mass structures and properties in the Ionian and Adriatic Seas. J. Geophys. Res. 2003, 108, 8102. [Google Scholar] [CrossRef] [Green Version]
- Mihanović, H.; Vilibić, I.; Dunić, N.; Šepić, J. Mapping of decadal middle Adriatic oceanographic variability and its relation to the BiOS regime: MAPPING OF DECADAL ADRIATIC VARIABILITY. J. Geophys. Res. Oceans 2015, 120, 5615–5630. [Google Scholar] [CrossRef]
- Mikuš, J. Population structure of calanoid copepods in the South Adriatic. Ph.D. Thesis, University of Zagreb, Zagreb, Croatia, 2011. [Google Scholar]
- De Olazabal, A.; Tirelli, V. First record of the egg-carrying calanoid copepod Pseudodiaptomus marinus in the Adriatic Sea. Mar. Biodivers. Rec. 2011, 4, e85. [Google Scholar] [CrossRef]
- Vidjak, O.; Bojanić, N.; de Olazabal, A.; Benzi, M.; Brautović, I.; Camatti, E.; Hure, M.; Lipej, L.; Lučić, D.; Pansera, M.; et al. Zooplankton in Adriatic port environments: Indigenous communities and non-indigenous species. Mar. Pollut. Bull. 2019, 147, 133–149. [Google Scholar] [CrossRef]
- Mazzocchi, M.G.; Christou, E.D.; Fragopoulu, N.; Siokou-Frangou, I. Mesozooplankton distribution from Sicily to Cyprus (Eastern Mediterranean): I. General aspects. Oceanol. Acta 1997, 20, 521–535. [Google Scholar]
- Licandro, P.; Icardi, P. Basin scale distribution of zooplankton in the Ligurian Sea (north-western Mediterranean) in late autumn. Hydrobiologia 2009, 617, 17–40. [Google Scholar] [CrossRef]
- Zunini Sertorio, T.C.; Licandro, P. Diel variation of mesozooplankton in epipelagic waters of the Ligurian Sea. In Atti del 11 Congresso dell’Associazione Italiana di Oceanologia e Limnologia (Sorrento, 26–28 Ottobre 1996); A.I.O.L: Genova, Italy, 1996; pp. 177–192. [Google Scholar]
- Cummings, J.A. Habitat dimensions of calanoid copepods in the western Gulf of Mexico. J. Mar. Res. 1984, 42, 163–188. [Google Scholar] [CrossRef]
- Fernández de Puelles; Gazá; Cabanellas-Reboredo; Santandreu; Irigoien; González-Gordillo; Duarte Zooplankton Abundance and Diversity in the Tropical and Subtropical Ocean. Diversity 2019, 11, 203. [CrossRef] [Green Version]
- Poulain, P.M.; Kourafalou, V.H.; Cushman-Roisin, B. Northern Adriatic Sea. In Physical Oceanography of the Adriatic Sea; Cushman-Roisin, B., Gačić, M., Poulain, P.M., Artegiani, A., Eds.; Springer: Dordrecht, The Netherlands, 2001; pp. 143–165. ISBN 978-90-481-5921-5. [Google Scholar]
- Gačić, M.; Civitarese, G.; Eusebi Borzelli, G.L.; Kovačević, V.; Poulain, P.M.; Theocharis, A.; Menna, M.; Catucci, A.; Zarokanellos, N. On the relationship between the decadal oscillations of the northern Ionian Sea and the salinity distributions in the eastern Mediterranean. J. Geophys. Res. 2011, 116. [Google Scholar] [CrossRef]
- Andersen, V. Zooplankton Community during the Transition from Spring Bloom to Oligotrophy in the Open NW Mediterranean and Effects of Wind Events. 1. Abundance and Specific Composition. J. Plankton Res. 2001, 23, 227–242. [Google Scholar] [CrossRef] [Green Version]
- Di Carlo, B.S.; Ianora, A.; Mazzocchi, M.G.; Scardi, M. Atlantis II Cruise: Uniformity of deep copepod assemblages in the Mediterranean Sea. J. Plankton Res. 1991, 13, 263–277. [Google Scholar] [CrossRef]
- Mauri, E.; Gerin, R.; Poulain, P.M. Measurements of water-mass properties with a glider in the South-western Adriatic Sea. J. Oper. Oceanogr. 2016, 9, s3–s9. [Google Scholar] [CrossRef]
- Dolan, J. Tintinnid ciliate diversity in the Mediterranean Sea: Longitudinal patterns related to water column structure in late spring-early summer. Aquat. Microb. Ecol. 2000, 22, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Kršinić, F.; Grbec, B. Some distributional characteristics of small zooplankton at two stations in the Otranto Strait (Eastern Mediterranean). Hydrobiologia 2002, 482, 119–136. [Google Scholar] [CrossRef]
- Krsinic, F. Tintinnids (Tintinnida, Choreotrichia, Ciliata) in the Adriatic Sea, Mediterranean. Part Taxon. Part II Ecol. Inst. Oceanogr. Fish. 2010, 113. [Google Scholar]
- Krsinic, F.; Grbec, B. Horizontal distribution of tintinnids in the open waters of the South Adriatic (Eastern Mediterranean). Sci. Mar. 2006, 70, 77–88. [Google Scholar] [CrossRef]
- Böttger-Schnack, R. Community structure and vertical distribution of cyclopoid copepods in the Red Sea: II. Aspects of seasonal and regional differences. Mar. Biol. 1990, 106, 487–501. [Google Scholar] [CrossRef]
- Böttger-Schnack, R. Vertical structure of small metazoan plankton, especially non-calanoid copepods. I. Deep Arabian Sea. J. Plankton Res. 1996, 18, 1073–1101. [Google Scholar]
- Böttger-Schnack, R. Vertical structure of small metazoan plankton, especially non-calanoid copepods. 2. Deep Eastern Mediterranean (Levantine sea). Oceanol. Acta 1997, 20, 399–419. [Google Scholar]
- Nishibe, Y.; Ikeda, T. Vertical distribution, abundance and community structure of oncaeid copepods in the Oyashio region, western subarctic Pacific. Mar. Biol. 2004, 145, 931–941. [Google Scholar] [CrossRef]
- Richter, C. Seasonal changes in the vertical distribution of mesozooplankton in the Greenland Sea Gyre (75°N): Distribution strategies of calanoid copepods. ICES J. Mar. Sci. 1995, 52, 533–539. [Google Scholar] [CrossRef]
- Alldredge, A.L. Abandoned Larvacean Houses: A Unique Food Source in the Pelagic Environment. Science 1972, 177, 885–887. [Google Scholar] [CrossRef]
- Lampitt, R.S.; Wishner, K.F.; Turley, C.M.; Angel, M.V. Marine snow studies in the Northeast Atlantic Ocean: Distribution, composition and role as a food source for migrating plankton. Mar. Biol. 1993, 116, 689–702. [Google Scholar] [CrossRef]
- Ohtsuka, S.; Kubo, N.; Okada, M.; Gushima, K. Attachment and feeding of pelagic copepods on larvacean houses. J. Oceanogr. 1993, 49, 115–120. [Google Scholar] [CrossRef]
- Paffenhöfer, G.A. On the ecology of marine cyclopoid copepods (Crustacea, Copepoda). J. Plankton Res. 1993, 15, 37–55. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stations | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Depth layers | 1 | 3 | 5 | 7 | 9 | 10 | 12 | 13 | 16 | 18 | 20 | GG |
| 0–50 | x | x | x | x ** | x | x | x | |||||
| 50–100 | x | x | x | x ** | x | x | x | |||||
| 0–100 | x | x * | x * | x | x | |||||||
| 100–200 | x ** | x | x | x ** | x | x | ||||||
| 200–300 | x | x | x ** | x | x | |||||||
| 300–400 | x * | x | x ** | x | ||||||||
| 400–600 | x * | x | x ** | |||||||||
| 600–800 | x * | x * | x ** | |||||||||
| 800–1200 | x * | x ** | ||||||||||
| Taxa | Mean (%) | a | b |
|---|---|---|---|
| Clausocalanus pergens | 7.96 | 5.0 ± 8.4 | 10.6 ± 17.4 |
| Oithona setigera-group | 7.28 | 2.0 ± 2.4 | 3.8 ± 6.2 |
| Oithona similis | 6.79 | 4.8 ± 3.4 | 12.2 ± 13.1 |
| Haloptilus longicornis | 6.61 | 2.6 ± 4.0 | 3.4 ± 2.7 |
| Oncaea spp. | 6.24 | 1.4 ± 2.0 | 3.3 ± 3.8 |
| Lucicutia flavicornis | 5.52 | 2.1 ± 0.2 | 4.6 ± 6.9 |
| Corycaeidae | 5.28 | 2.2 ± 3.8 | 3.6 ± 3.4 |
| Ctenocalanus vanus | 5.27 | 3.6 ± 4.1 | 13.8 ± 23.8 |
| Neomormonilla minor | 4.0 | 0.9 ± 1.0 | 1.9 ± 0.9 |
| Pleuromamma gracilis | 2.75 | 0.8 ± 1.0 | 2.5 ± 3.7 |
| Mecynocera clausi | 2.66 | 2.5 ± 3.0 | 3.8 ± 4.0 |
| Paracalanus parvus | 2.31 | 4.6 ± 7.1 | 19.5 ± 28.9 |
| Clausocalanus paululus | 1.87 | 0.9 ± 1.4 | 2.8 ± 3.5 |
| Clausocalanus jobei | 1.63 | 4.2 ± 7.4 | 5.2 ± 7.3 |
| Clausocalanus furcatus | 1.57 | 1.8 ± 1.8 | 5.5 ± 5.7 |
| Paraeuchaeta hebes | 1.41 | 1.2 ± 1.5 | 5.9 ± 7.2 |
| Monacilla typica | 1.38 | 0.2 ± 0.1 | 0.4 ± 0.4 |
| Calocalanus styliremis | 1.35 | 2.0 ± 3.2 | 2.6 ± 1.9 |
| Spinocalanus longicornis | 1.32 | 0.2 ± 0.1 | 0.4 ± 0.4 |
| Clausocalanus lividus | 1.27 | 1.6 ± 2.5 | 3.2 ± 4.1 |
| Acartia (Acartiura) clausi | 1.17 | 10.8 ± 21.2 | 3.2 ± 5.1 |
| Mesocalanus teniucornis | 1.09 | 0.6 ± 0.7 | 1.6 ± 2.1 |
| Pleuromamma abdominalis | 1.08 | 0.2 ± 0.3 | 0.9 ± 1.3 |
| Clausocalanus parapergens | 1.05 | 0.4 ± 0.2 | 1.9 ± 1.3 |
| Cluster 1 | IV | Ab | C | ||
|---|---|---|---|---|---|
| Acartia (Acartiura) clausi | 85.1 | 6.8 | 3.6 | Av. Abundance | 264 ± 99 ind. m−3 |
| Paracalanus parvus | 76.7 | 15.4 | 7.8 | d | 7.19 |
| Clausocalanus furcatus | 70.9 | 6.0 | 4.2 | H’ | 2.62 |
| Paraeuchaeta hebes | 67.4 | 6.3 | 3.7 | ||
| Ctenocalanus vanus | 62.3 | 24.3 | 13.6 | ||
| Candacia giesbrechti | 58.9 | 0.9 | 0.6 | ||
| Calanus helgolandicus | 57.2 | 1.1 | 0.8 | ||
| Clausocalanus arcuicornis | 57.0 | 2.7 | 1.4 | ||
| Oithona similis | 50.5 | 14.9 | 8.2 | ||
| Centropages typicus | 46.6 | 1.5 | 0.7 | ||
| Diaixis pygmaea | 45.3 | 0.57 | 0.5 | ||
| Clausocalanus jobei | 45.5 | 6.9 | 3.5 | ||
| Clausocalanus pergens | 44.5 | 19.8 | 11.5 | ||
| Oithona plumifera | 43.7 | 12.3 | 6.8 | ||
| Neocalanus gracilis | 43.0 | 0.6 | 0.5 | ||
| Pareucalanus attenuatus | 42.2 | 0.1 | 0.1 | ||
| Temora stylifera | 40.6 | 1.3 | 0.7 | ||
| Clausocalanus lividus | 40.5 | 3.3 | 3.1 | ||
| Paracalanus nanus | 39.1 | 1.1 | 0.6 | ||
| Oncaeidae | 36.0 | 4.3 | 2.9 | ||
| Clausocalanus mastigophorus | 35.8 | 2.0 | 1.1 | ||
| Mesocalanus tenuicornis | 34.7 | 1.0 | 0.7 | ||
| Oithona nana | 30.5 | 0.6 | 0.4 | ||
| Calocalanus pavo | 30.2 | 0.8 | 0.8 | ||
| Nannocalanus minor | 29.2 | 0.6 | 0.3 | ||
| Clausocalanus juv. | 25.7 | 5.4 | 2.7 | ||
| Cluster 2 | |||||
| Sapphirina spp. | 76.8 | 0.3 | 0.4 | Av. Abundance | 111 ± 49 ind. m−3 |
| Mecynocera clausi | 50.3 | 4.7 | 6.9 | d | 8.35 |
| Scolecitrix bradyi | 47.2 | 0.2 | 0.4 | H’ | 2.75 |
| Calocalanus styliremis | 46.9 | 2.5 | 3.0 | ||
| Calocalanus contractus | 46.1 | 0.9 | 1.5 | ||
| Lucicutia flavicornis | 41.9 | 7.0 | 8.5 | ||
| Aetidaeus giesbrechti | 41.9 | 0.2 | 0.3 | ||
| Corycaeidae | 40.5 | 4.8 | 7.1 | ||
| Aetidaeus armatus | 39.7 | 0.3 | 0.4 | ||
| Scolecithricella dentata | 36.0 | 0.5 | 0.6 | ||
| Pleuromamma abdominalis | 33.6 | 0.8 | 0.9 | ||
| Calocalanus juv. | 25.4 | 2.8 | 3.7 | ||
| Cluster 3 | |||||
| Chiridius poppei | 48.3 | 0.1 | 0.4 | Av. Abundance | 50 ± 14.9 ind. m−3 |
| Neomormonilla minor | 38.0 | 1.7 | 5.7 | d | 8.93 |
| Euchaeta acuta | 37.3 | 0.3 | 1.1 | H’ | 2.54 |
| Oithona setigera-group | 35.0 | 4.6 | 14.6 | ||
| Heterorhabdus spinifrons | 34.7 | 0.2 | 0.4 | ||
| Haloptilus longicornis | 34.5 | 3.7 | 11.0 | ||
| Clausocalanus parapergens | 33.2 | 1.1 | 4.1 | ||
| Pleuromamma gracilis | 33.5 | 2.0 | 6.4 | ||
| Lucicutia clausi | 25.1 | 0.4 | 1.0 | ||
| Cluster 4 | |||||
| Spinocalanus longicornis | 27.5 | 0.2 | 3.2 | Av. Abundance | 16 ± 9 ind. m−3 |
| d | 12.16 | ||||
| H’ | 2.27 | ||||
| Cluster 5 | |||||
| Temoropia mayumbaensis | 67.5 | 0.2 | 5.7 | Av. Abundance | 4 ± 1.2 ind. m−3 |
| Monacilla typica | 44.0 | 0.3 | 14.0 | d | 22.1 |
| Spinocalanus oligospinosus | 43.9 | 0.1 | 2.9 | H’ | 1.96 |
| Candacia elongata | 28.6 | 0.004 | 0.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hure, M.; Batistić, M.; Kovačević, V.; Bensi, M.; Garić, R. Copepod Community Structure in Pre- and Post- Winter Conditions in the Southern Adriatic Sea (NE Mediterranean). J. Mar. Sci. Eng. 2020, 8, 567. https://doi.org/10.3390/jmse8080567
Hure M, Batistić M, Kovačević V, Bensi M, Garić R. Copepod Community Structure in Pre- and Post- Winter Conditions in the Southern Adriatic Sea (NE Mediterranean). Journal of Marine Science and Engineering. 2020; 8(8):567. https://doi.org/10.3390/jmse8080567
Chicago/Turabian StyleHure, Marijana, Mirna Batistić, Vedrana Kovačević, Manuel Bensi, and Rade Garić. 2020. "Copepod Community Structure in Pre- and Post- Winter Conditions in the Southern Adriatic Sea (NE Mediterranean)" Journal of Marine Science and Engineering 8, no. 8: 567. https://doi.org/10.3390/jmse8080567