Tool Use by Four Species of Indo-Pacific Sea Urchins

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
Statistical Analysis
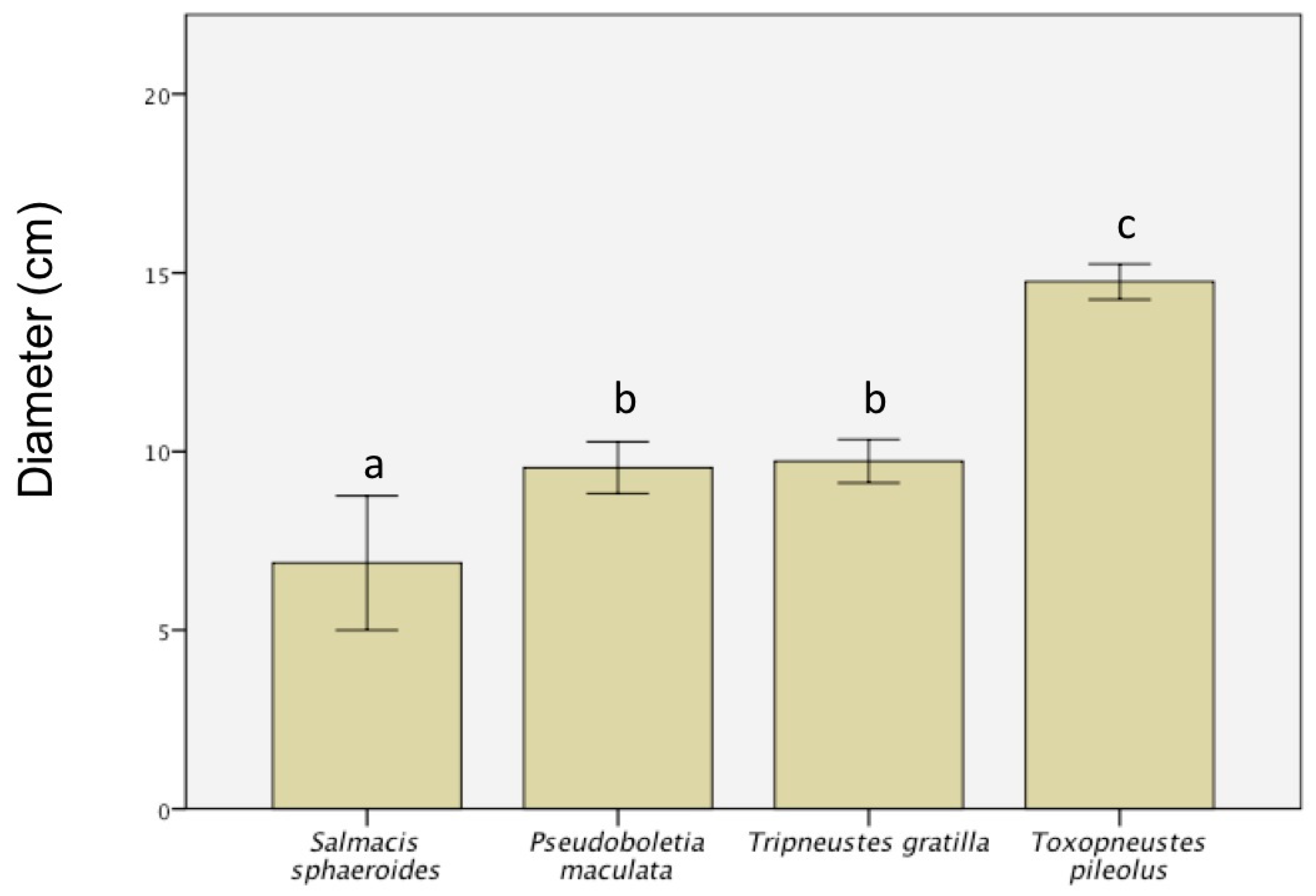
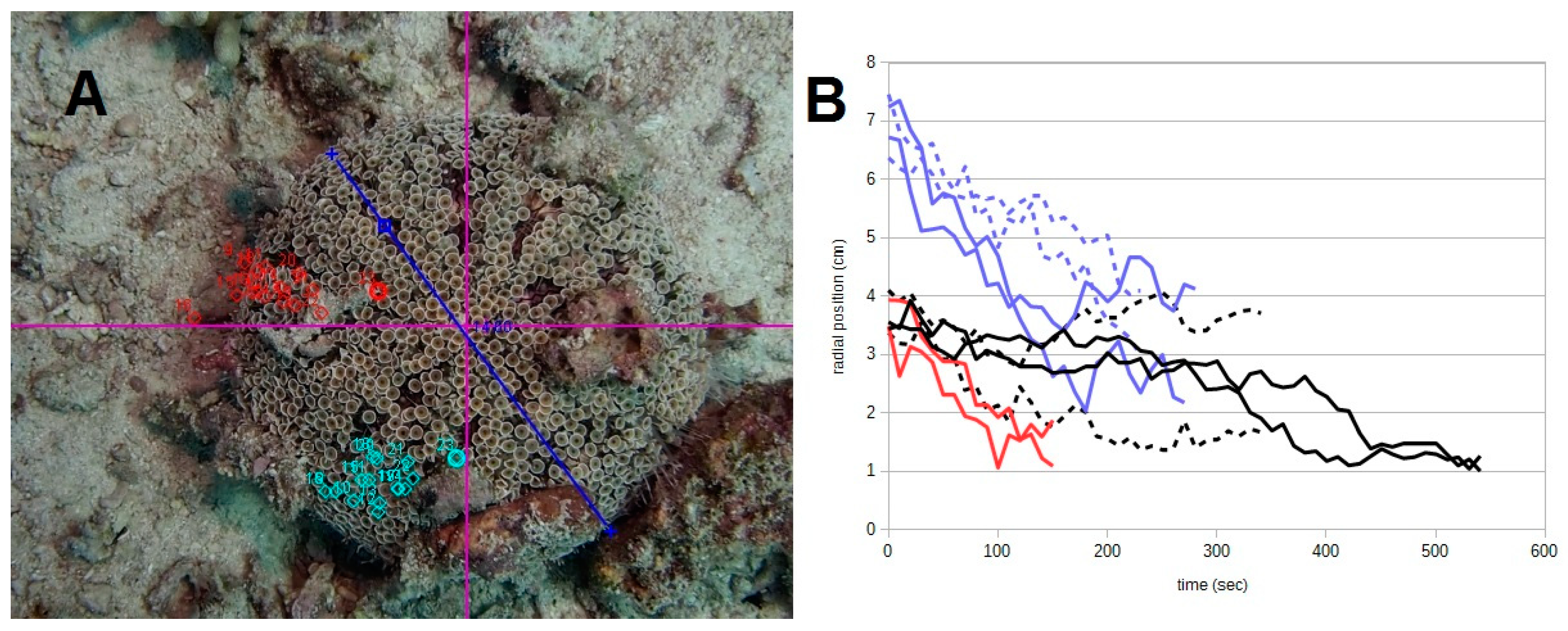
3. Results
4. Discussion
4.1. Echinoid Covering Behavior Evolution
4.2. Echinoid Covering Behavior as Tool Use
- -
- The use of selected pieces of debris as a cover by sea urchins (as shown in this and other studies [5,13]). By no means is the covering material simply debris that gets stuck on the sea urchin, but it is there through the active selection and choice of the animal, as a consequence of the coordinated action of tube-feet and spines in response to the environment.
- -
- The adjustment of debris cover according to need (increased coverage in response to increased UV radiation [2], and the increased coverage at shallower depth as demonstrated in this study) also point to goal-directed tool-use behavior.
- -
- The difference in debris type between echinoid species points toward a specific form of tool use tailored to the needs and capabilities of the different sea urchin species.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Agatsuma, Y. Effect of the covering behavior of the juvenile sea urchin Strongylocentrotus intermedius on predation by the spider crab Pugettia quadridens. Fish Sci. 2001, 67, 1181–1183. [Google Scholar] [CrossRef]
- Dumont, C.P.; Drolet, D.; Deschênes, I.; Himmelman, J.H. Multiple factors explain the covering behaviour in the green sea urchin, Strongylocentrotus droebachiensis. Anim. Behav. 2007, 73, 979–986. [Google Scholar] [CrossRef]
- Zhao, C.; Zhou, H.; Tian, X.; Feng, W.; Chang, Y. The effects of prolonged food deprivation on the covering behavior of the sea urchins Glyptocidaris crenularis and Strongylocentrotus intermedius. Mar. Freshw. Behav. Physiol. 2014, 47, 11–18. [Google Scholar] [CrossRef]
- Wei, J.; Zhang, L.; Zhao, C.; Feng, W.; Sun, P.; Chang, Y. Correlation analyses of covering and righting behaviors to fitness related traits of the sea urchin Glyptocidaris crenularis in different environmental conditions. Chin. J. Oceanol. Limnol. 2016, 34, 1183–1190. [Google Scholar] [CrossRef]
- Ziegenhorn, M.A. Best dressed test: A study of the covering behavior of the collector urchin Tripneustes gratilla. PLoS ONE 2016, 11, e0153581. [Google Scholar] [CrossRef]
- Belleza, D.; Abao, R.; Taguba, C.; Dy, D. Effects of UV-C on the masking behavior of the green urchin Salmacis sphaeroides (Linnaeus, 1758). Philipp. Sci. 2013, 49, 34–43. [Google Scholar]
- David, B.; Magniez, F.; Villier, L. Conveying behaviour of the deep sea pourtalesiid Cystocrepis setigera off Peru. Echinoderm Res. 2001, 1, 253–257. [Google Scholar]
- Brothers, C.; Smith, K.; Amsler, M.; Aronson, R.; Singh, H.; McClintock, J. Covering behavior of deep-water echinoids in Antarctica: Possible response to predatory king crabs. Mar. Ecol. Prog. Ser. 2016, 553, 155–162. [Google Scholar] [CrossRef]
- Richner, H.; Milinski, M. On the functional significance of masking behaviour in sea urchins-an experiment with Paracentrotus lividus. Mar. Ecol. Prog. Ser. 2000, 205, 307–308. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Ji, N.; Zhang, B.; Sun, P.; Feng, W.; Wei, J.; Chang, Y. Effects of covering behavior and exposure to a predatory crab Charybdis japonica on survival and HSP70 expression of juvenile sea urchins Strongylocentrotus intermedius. PLoS ONE 2014, 9, e97840. [Google Scholar] [CrossRef]
- Dix, T.G. Covering response of the Echinoid Evechinus chloroticus (Val.). Pac. Sci. 1970, 24, 187–194. [Google Scholar]
- Douglas, C.A. Availability of drift materials and the covering response of the sea urchin Strongylocentrotus purpuratus (Stimpson). Pac. Sci. 1976, 30, 83–89. [Google Scholar]
- Amato, K.R.; Emel, S.L.; Lindgren, C.A.; Sullan, K.M.; Wright, P.R.; Gilbert, J.J. Covering behavior of two co-occurring Jamaican sea urchins: Differences in the amount of covering and selection of covering material. Bull. Mar. Sci. 2008, 82, 255–261. [Google Scholar]
- Claereboudt, E.J.S.; Claereboudt, M.R. Quantifying sea urchins covering behavior by image analysis. Cah. Biol. Mar. 2013, 54, 587–592. [Google Scholar]
- Sigg, J.E.; Lloyd-Knight, K.M.; Boal, J.G. UV radiation influences covering behaviour in the urchin Lytechinus variegatus. J. Mar. Biol. Assoc. UK 2007, 87, 1257–1261. [Google Scholar] [CrossRef]
- Crook, A.C. Individual Variation in the Covering Behaviour of the Shallow Water Sea Urchin Paracentrotus lividus. Mar. Ecol. 2003, 24, 275–287. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.A.; Yusoff, F.M.; Arshad, A.; Ara, R. Growth and survival of the tropical sea urchin, Salmacis sphaeroides fed with different macroalgae in captive rearing condition. J. Environ. Biol. 2016, 37, 855–862. [Google Scholar]
- Lees, D.C.; Carter, G.A. The covering response to surge, sunlight and ultraviolet light in Lytechinus anamesus. Ecology 1972, 53, 1127–1133. [Google Scholar] [CrossRef]
- Pawson, D.L.; Pawson, D.J. Bathyal sea urchins of the Bahamas, with notes on covering behavior in deep sea echinoids (Echinodermata: Echinoidea). Deep Sea Res. Part II Top. Stud. Oceanogr. 2013, 92, 207–213. [Google Scholar] [CrossRef]
- James, D.W. Diet, movement, and covering behavior of the sea urchin Toxopneustes roseus in rhodolith beds in the Gulf of California, México. Mar. Biol. 2000, 137, 913–923. [Google Scholar] [CrossRef]
- Adams, N.L. UV Radiation Evokes Negative Phototaxis and Covering Behavior in the Sea Urchin Strongylocentrotus droebachiensis; Marine Ecology Progress Series; Inter-Research Science Center: Luhe, Germany, 2001; pp. 87–95. [Google Scholar]
- Zhao, C.; Bao, Z.; Chang, Y. Fitness-related consequences shed light on the mechanisms of covering and sheltering behaviors in the sea urchin Glyptocidaris crenularis. Mar. Ecol. 2016, 37, 998–1007. [Google Scholar] [CrossRef]
- Levin, L.A.; Gooday, A.J.; James, D.W. Dressing up for the deep: Agglutinated protists adorn an irregular urchin. J. Mar. Biol. Assoc. UK 2001, 81, 881. [Google Scholar] [CrossRef]
- Ferber, I.; Lawrence, J.M. Distribution, substratum preference and burrowing behaviour of Lovenia elongata (Gray) (Echinoidea: Spatangoida) in the Gulf of Elat (Aqaba), Red Sea. J. Exp. Mar. Biol. Ecol. 1976, 22, 207–225. [Google Scholar] [CrossRef]
- Kroh, A.; Smith, A.B. The phylogeny and classification of post-Palaeozoic echinoids. J. Syst. Palaeontol. 2010, 8, 147–212. [Google Scholar] [CrossRef] [Green Version]
- McGhee, G.R. Convergent Evolution: Limited forms Most Beautiful; MIT Press: Cambridge, MA, USA, 2011. [Google Scholar]
- Smith, E.; Bentley-Condit, V. Animal tool use: Current definitions and an updated comprehensive catalogue. Behaviour 2010, 147, 185–32A. [Google Scholar]
- Milikan, G.; Bowman, R.L. Observations in Galapagos tool-using finches in captivity. Living Bird 1967, 6, 23–42. [Google Scholar]
- Van Lawick-Goodall, J. Tool-using in primates and other vertebrates. Adv. Stud. Behav. 1971, 3, 195–249. [Google Scholar]
- St Amant, R.; Horton, T.E. Revisiting the definition of animal tool use. Anim. Behav. 2008, 75, 1199–1208. [Google Scholar] [CrossRef]
- Shumaker, R.W.; Walkup, K.R.; Beck, B.B. Animal Tool Behavior: The Use and Manufacture of Tools by Animals; Johns Hopkins University Press: Baltimore, MD, USA, 2011. [Google Scholar]
- Lawrence, J.M. Covering response in sea urchins. Nature 1976, 262, 490–491. [Google Scholar] [CrossRef]
- Mayr, E. Cause and effect in biology. Science 1961, 134, 1501–1506. [Google Scholar] [CrossRef]
- Laland, K.N.; Sterelny, K.; Odling-Smee, J.; Hoppitt, W.; Uller, T. Cause and Effect in Biology Revisited: Is Mayr’s Proximate-Ultimate Dichotomy Still Useful? Science 2011, 334, 1512–1516. [Google Scholar] [CrossRef]
- Seed, A.; Byrne, R. Animal Tool-Use. Curr. Biol. 2010, 20, R1032–R1039. [Google Scholar] [CrossRef]
- Mann, J.; Patterson, E.M. Tool use by aquatic animals. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20120424. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order | Family | Genus | Species | Distribution | Reference |
|---|---|---|---|---|---|
| Camarodonta | Parechinidae | Paracentrotus | lividus | Mediterranean Sea and eastern Atlantic Ocean | [16] |
| Camarodonta | Temnopleuridae | Salmacis | sphaeroides | Tropical Indo-Pacific | [24] |
| Camarodonta | Toxopneustidae | Lytechinus | anamesus | Caribbean | [25] |
| Camarodonta | Toxopneustidae | Lytechinus | euerces | Deep sea, Caribbean | [26] |
| Camarodonta | Toxopneustidae | Lytechinus | variegatus | Caribbean | [13] |
| Camarodonta | Toxopneustidae | Toxopneustes | roseus | Gulf of California | [27] |
| Camarodonta | Toxopneustidae | Tripneustes | gratilla | Tropical Indo-Pacific | This study |
| Camarodonta | Toxopneustidae | Tripneustes | ventricosus | Caribbean | [13] |
| Camarodonta | Toxopneustidae | Pseudoboletia | maculata | Tropical Indo-Pacific | This study |
| Camarodonta | Toxopneustidae | Toxopneustes | pileolus | Tropical Indo-Pacific | This study |
| Echinoida | Strongylocentrotidae | Strongylocentrotus | droebachiensis | Temperate Atlantic | [18] |
| Echinoida | Strongylocentrotidae | Strongylocentrotus | intermedius | Temperate and cold northern hemisphere | [1] |
| Cassiduloida | Echinolampadidae | Conolampas | sigsbei | Deep sea, Caribbean | [26] |
| Spatangoida | - | Paleobrissus | hilgardi | Deep sea, Caribbean | [26] |
| Stomopneustoida | Glyptocidaridae | Glyptocidaris | Crenularis | Temperate and cold northern hemisphere | [3] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barrett, G.A.; Revell, D.; Harding, L.; Mills, I.; Jorcin, A.; Stiefel, K.M. Tool Use by Four Species of Indo-Pacific Sea Urchins. J. Mar. Sci. Eng. 2019, 7, 69. https://doi.org/10.3390/jmse7030069
Barrett GA, Revell D, Harding L, Mills I, Jorcin A, Stiefel KM. Tool Use by Four Species of Indo-Pacific Sea Urchins. Journal of Marine Science and Engineering. 2019; 7(3):69. https://doi.org/10.3390/jmse7030069
Chicago/Turabian StyleBarrett, Glyn A., Dominic Revell, Lucy Harding, Ian Mills, Axelle Jorcin, and Klaus M. Stiefel. 2019. "Tool Use by Four Species of Indo-Pacific Sea Urchins" Journal of Marine Science and Engineering 7, no. 3: 69. https://doi.org/10.3390/jmse7030069