In Vitro Inhibition of Acetylcholinesterase, Alphaglucosidase, and Xanthine Oxidase by Bacteria Extracts from Coral Reef in Hainan, South China Sea


Abstract
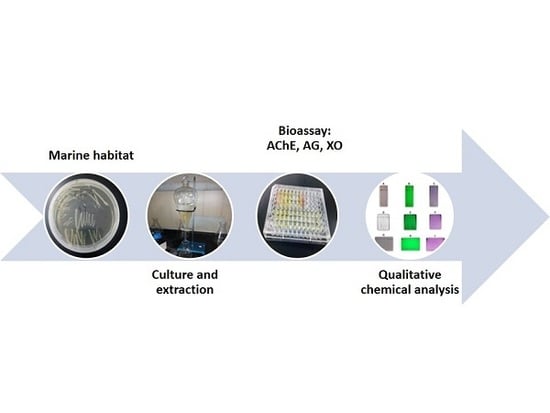
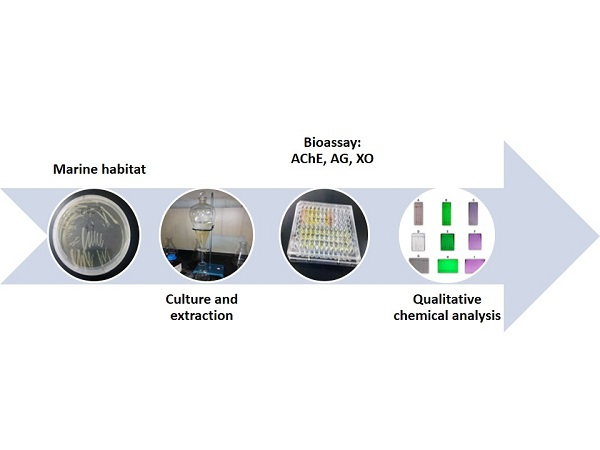
:
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Marine Bacterial Strains
2.3. Extraction Process
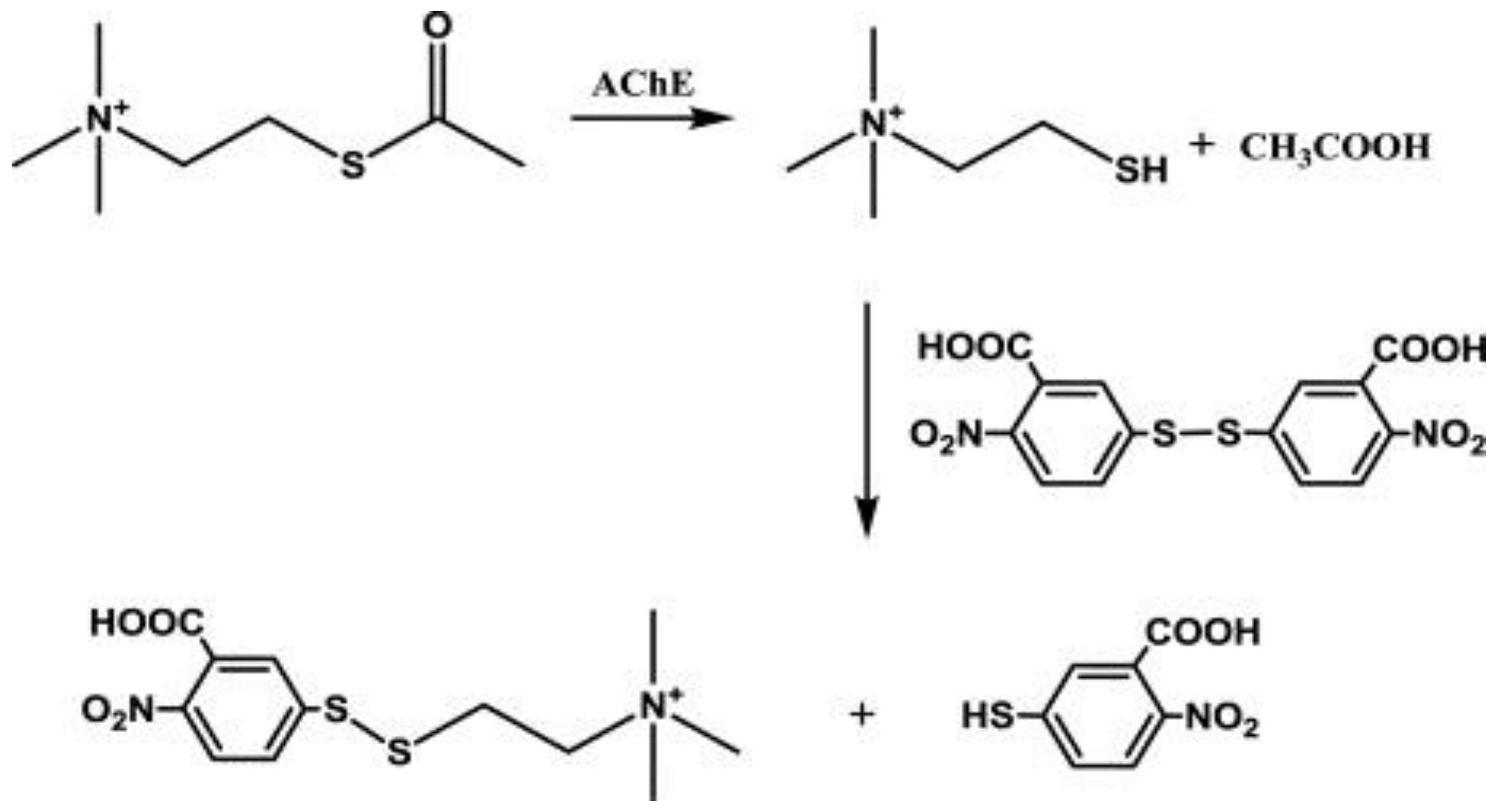
2.4. Acetylcholinesterase Inhibition Assay
2.5. Alphaglucosidase Inhibition Assay
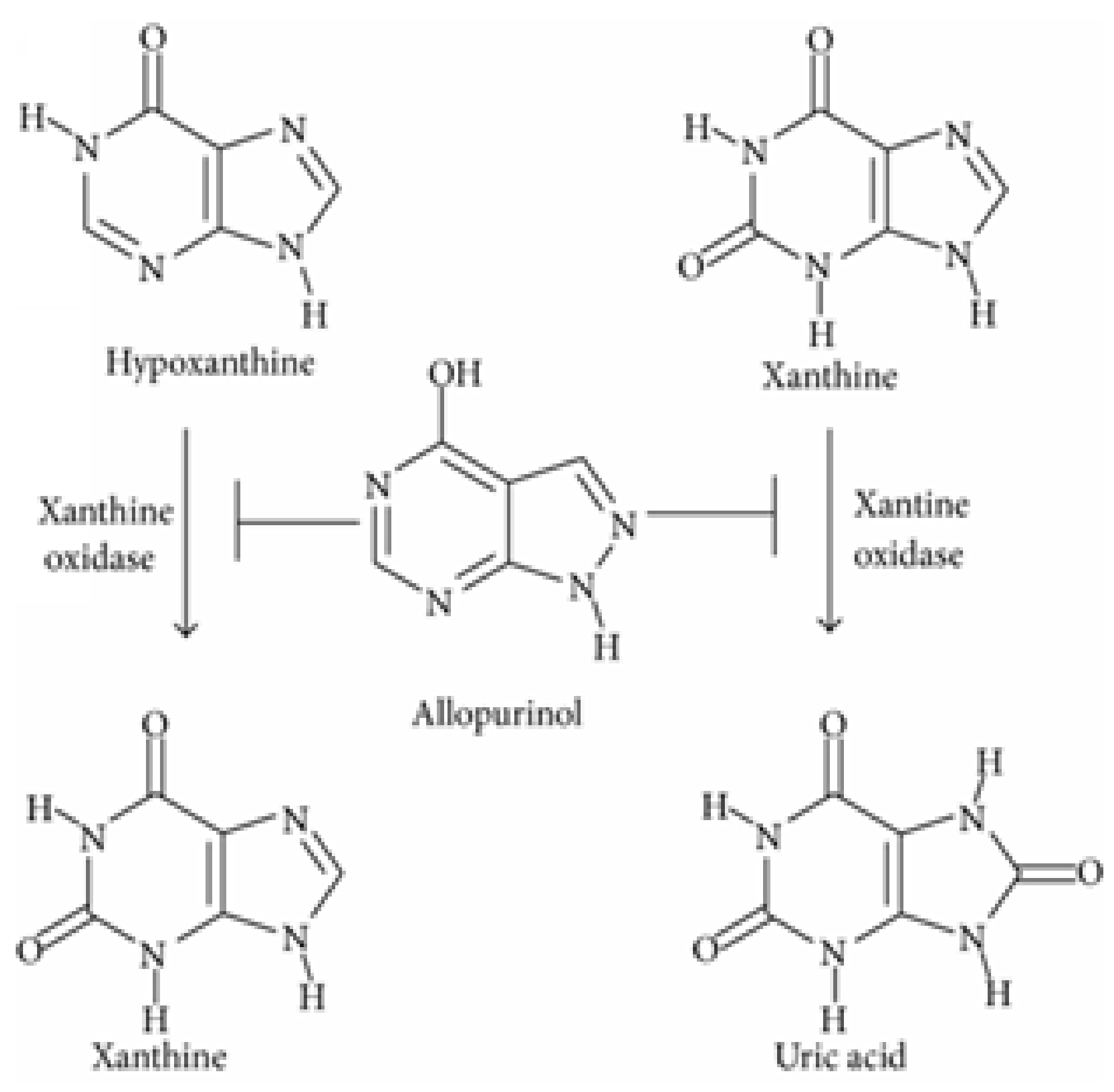
2.6. Xanthine Oxidase Inhibition Assay
2.7. Qualitative Chemical Analysis
2.8. Screening of Antagonistic Bacteria
2.9. Activity: Screening of Antagonistic Bacteria
3. Results
3.1. Acetylcholinesterase and Alphaglucosidase Inhibition Assay
3.2. Xanthine Oxidase Inhibition Assay
3.3. Qualitative Chemical Analysis
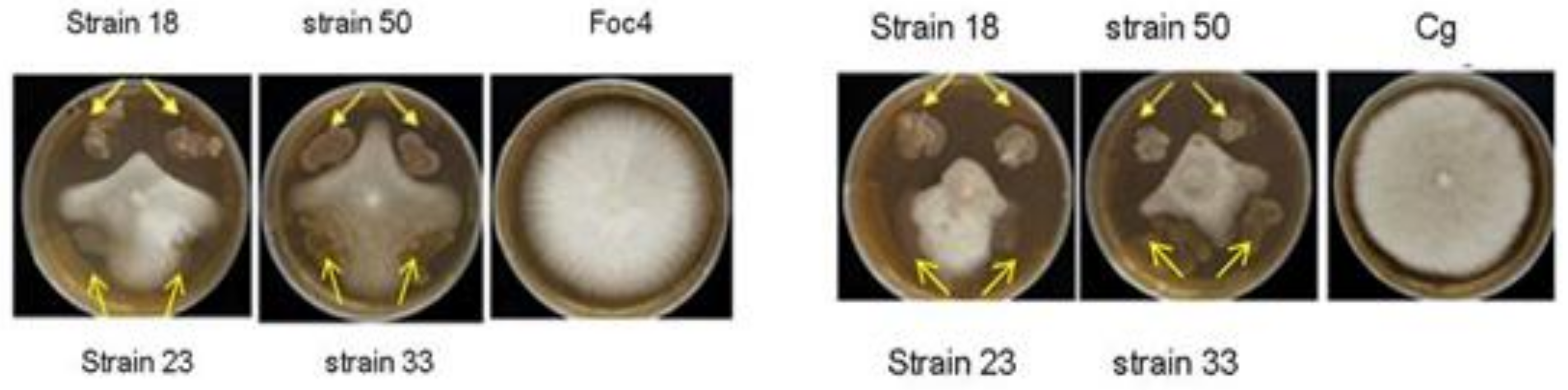
3.4. Screening of Antagonistic Bacteria
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wiesner, J.; Kriz, Z.; Kuca, K.; Jun, D.; Koca, J. Acetylcholinesterases—The structural similarities and differences. J. Enzym. Inhib. Med. Chem. 2007, 22, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.T. Molecular interactions of cholinesterases inhibitors using in silico methods: Current status and future prospects. New Biotechnol. 2009, 25, 331–346. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, M.; Lee, T. Galanthamine from snowdrop—The development of a modern drug against Alzheimer’s disease from local Caucasian knowledge. J. Ethnopharmacol. 2004, 92, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Yan, H.; Tang, X.C. Progress in studies of huperzine A, a natural cholinesterase inhibitor from Chinese herbal medicine. Acta Pharmacol. Sin. 2006, 27, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Bonnard, I.; Jhaumeer-Laulloo, S.B.; Bontemps, N.; Banaigs, B.; Aknin, M. New Lobane and Cembrane Diterpenes from Two Comorian Soft Corals. Mar. Drugs 2010, 8, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Langjae, R.; Bussarawit, S.; Yuenyongsawad, S.; Ingkaninan, K.; Plubrukarn, A. Acetylcholinesterase inhibiting steroidal alkaloid from the sponge Corticium sp. Steroids 2007, 72, 682–685. [Google Scholar] [CrossRef] [PubMed]
- Yoon, N.Y.; Chung, H.Y.; Kim, H.R.; Choi, J.S. Acetyl and butyryl cholinesterase inhibitory activities of sterols and phlorotannins from Ecklonia stolonifera. Fish. Sci. 2008, 74, 200–207. [Google Scholar] [CrossRef]
- Nukoolkarn, V.S.; Saen-oon, S.; Rungrotmongkol, T.; Hannongbua, S.; Ingkaninan, K.; Suwanborirux, K. Petrosamine, a potent anticholinesterase pyridoacridine alkaloid from a Thai marine sponge Petrosia n. sp. Bioorg. Med. Chem. 2008, 16, 6560–6567. [Google Scholar] [CrossRef] [PubMed]
- Kalauni, S.K.; Choudhary, M.I.; Khalid, A.; Manandhar, M.D.; Shaheen, F.; Gewali, M.B. New cholinesterase inhibiting steroidal alkaloids from the leaves of Sarcococca coriacea of Nepalese origin. Chem. Pharm. Bull. 2002, 50, 1423–1426. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Wahab, A.T.; Nawas, S.A.; Choudhary, M.I. New cholinesterase inhibiting bisbenzylisoquinoline alkaloids from Cocculus pendulus. Chem. Pharm. Bull. 2004, 52, 802–806. [Google Scholar] [CrossRef]
- Ahmad, W.; Ahmad, B.; Ahmad, M.; Iqbal, Z.; Nisar, M.; Ahmad, M. In vitro inhibition of acetylcholinesterase, butyrylcholinesterase and lipoxygenase by crude extract of Myricaria elegans Royle. J. Biol. Sci. 2003, 11, 1046–1049. [Google Scholar]
- Sung, S.Y.; Kang, S.Y.; Lee, K.Y.; Park, M.J.; Kim, J.H.; Park, J.H.; Kim, Y.C.; Kim, J.; Kim, Y.C. (+)-α-Viniferin, a stilbene trimer from Caranga chamlague inhibits acetylcholinesterase. Biol. Pharm. Bull. 2002, 25, 125–127. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association (ADA). Diagnosis and classification of diabetes mellitus. Diabetes Care 2009, 101, 274. [Google Scholar]
- Fauci, A.S.; Braunwald, E.; Kasper, D.L. Diabetes Mellitus. In Principles of Internal Medicine; Mc-Graw Hill: New York, NY, USA, 2009. [Google Scholar]
- Abesundara, K.J.; Matsui, T.; Matsumoto, K. Alfa-glucosidase inhibitory activity of some Sri Lanka plant extracts, one of which, Cassia auriculata, exerts a strong antihyperglycemic effect in rats comparable to therapeutic drug acarbose. J. Agric. Food Chem. 2004, 52, 2541. [Google Scholar] [CrossRef] [PubMed]
- Önal, S.; Timmur, S.; Okuttucu, B.; Zihnioglu, A. Inhibition of α-glucosidase by aqueous extracts of some potent antidiabetic medicinal herbs. Prep. Biochem. Biotechnol. 2005, 35, 29. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wen, S.; Kota, B.P.; Peng, G.; Li, G.Q.; Yamahara, J.; Roufogalis, B.D. Punica granatum flower extract, a potent α-glucosidase inhibitor, improves postprandial hyperglycemia in Zucker diabetic fatty rats. J. Ethnopharmacol. 2005, 99, 239. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.; Park, M.; Chul, H.L.; Kang, Y.; Seok-Kang, E.; Ki-Ki, S. Antidiabetic agents from medicinal plants. Curr. Med. Chem. 2006, 13, 1. [Google Scholar] [CrossRef]
- Copeland, R.A. Why Enzymes as Drug Targets? Evaluation of Enzyme Inhibitors in Drug Discovery; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 1–23. [Google Scholar]
- Choma, I.M.; Grzelak, E.M. Bioautography detection in thin layer chromatography. J. Chromatogr. A 2011, 1218, 2684–2691. [Google Scholar] [CrossRef] [PubMed]
- Montaser, R.; Luesch, H. Marine natural products: A new wave of drugs? Future. Med. Chem. 2011, 3, 1475–1489. [Google Scholar] [CrossRef] [PubMed]
- Reid, P.C.; Gorick, G.; Edwards, M. Climate Change and Marine Ecosystem Research; Sir Alister Hardy Foundation for Ocean Science (SAHFOS): Citadel Hill, UK, 2011. [Google Scholar]
- Huehn, S.; Eichhorn, C.; Urmersbach, S.; Breidenbach, J.; Bechlars, S.; Bier, N.; Alter, T.; Bartelt, E.; Frank, C.; Appel, B.; et al. Pathogenic vibrios in environmental, seafood and clinical sources in Germany. Int. J. Med. Microbiol. 2014, 304, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Le Roux, F.; Wegner, K.M.; Baker-Austin, C.; Vezzulli, L.; Osorio, C.R.; Amaro, C. The emergence of Vibrio pathogens in Europe: Ecology, evolution, and pathogenesis, Paris, 11–12th March 2015. Front. Microbiol. 2015, 6, 830. [Google Scholar] [PubMed]
- Vaishampayan, P.; Probst, A.; Krishnamurthi, S.; Ghosh, S.; Osman, S.; McDowall, A.; Ruckmani, A.; Mayilraj, S.; Venkateswaran, K. Bacillus horneckiae sp. nov., isolated from a spacecraft-assembly clean room. Int. J. Syst. Evol. Microbiol. 2010, 60, 1031–1037. [Google Scholar] [CrossRef] [PubMed]
- Seiler, H.; Wenning, M.; Scherer, S. Domibacillus robiginosus gen. nov., sp. nov., isolated from a pharmaceutical clean room. Int. J. Syst. Evol. Microbiol. 2013, 63, 2054–2061. [Google Scholar] [CrossRef] [PubMed]
- Bottone, E.J. Bacillus cereus, a volatile human pathogen. Clin. Microbiol. Rev. 2010, 23, 382–398. [Google Scholar] [CrossRef] [PubMed]
- Marquez, M.C.; Sanchez-Porro, C.; Ventosa, A. Hallophilic and Haloakalophilic, Aerobic Endospore-Forming Bacteria in Soil; Springer: New York, NY, USA, 2011; pp. 309–339. [Google Scholar]
- Schmidt, T.R.; Scott, E.J.; Dyer, D.W. Whole-genome phylogenies of the family Bacillaceae and expansion of the sigma factor gene family in the Bacillus cereus species-group. BMC Genom. 2011, 12, 430. [Google Scholar] [CrossRef] [PubMed]
- Hoyles, L.; Honda, H.; Logan, N.; Halket, G.; La Ragione, R.; McCartney, A. Recognition of greater diversity of Bacillus species and related bacteria in human faeces. Res. Microbiol. 2012, 163, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, D.; Andres, V.; Featherston, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Miroslav, P.; Martina, H.; Kamil, K.; Jean-Pierre, S. Assessment of Acetylcholinesterase Activity Using Indoxylacetate and Comparison with the Standard Ellman’s Method. Int. J. Mol. Sci. 2011, 12, 2631–2640. [Google Scholar]
- Danijela, K.; Danica, D.; Gordana, S.; Ivan, P.; Aleksandra, Y.; Jovana, I. Xanthine Oxidase: Isolation, Assays of Activity, and Inhibition. J. Chem. 2015. [Google Scholar] [CrossRef]
- Pistia-Brueggeman, G.; Hollingsworth, R.I. A preparation and screening strategy for glycosidase inhibitors. Tetrahedron 2001, 57, 8773. [Google Scholar] [CrossRef]
- Valentao, P.; Frenandes, E.; Carvalho, F.; Andrade, B.P.; Seabra, R.M.; Bastos, M.L. Studies on the antioxidant activity of Lippia citfiodora infusion: Scavenging effect on superoxide radical, hydroxyl radical and hypochlorous acid. Biol. Pharm. Bull. 2002, 25, 1324–1327. [Google Scholar] [CrossRef] [PubMed]
- López-Cruz, R.I.; Zenteno-Savín, T.; Galván-Magaña, F. Superoxide production, oxidative damage and enzymatic antioxidant defenses in shark skeletal muscle. Comp. Biochem. Physiol. 2010, 156, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Harborne, J.B. A Guide to Modern Techniques of Plant Analysis, Chapman and Hall, London. Phytochem. Methods 1980, 278, 1980. [Google Scholar]
- Feng, S.; Wang, R.; Lin, K.; Zhang, Y.; Du, L.; Fan, X.; Cao, W. Identification of Strain Bs-208 and Its Inhibition Against Plant Pathogenic Fungi. Chin. J. Biol. Control 2003, 19, 171–174. [Google Scholar]
- Bladt, S. Plant Drug Analysis: A Thin Layer Chromatography Atlas; Springer: Berlin/Heidelberg, Germany, 1996. [Google Scholar]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the 30 Years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [PubMed]
- Fusetani, N. Antifouling marine natural products. Nat. Prod. Rep. 2011, 28, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.T.; Hamann, M.T.; Enticknap, J.J.; Rao, K.V. Kahalalide-Producing Bacteria; PCT/US2004/036201 May 2005. Available online: http://www.wipo.int/patentscope/search/en/WO2005042720 (accessed on 16 August 2011).
- Mansson, M.; Nielsen, A.; Kjærulff, L.; Gotfredsen, C.H.; Wietz, M.; Ingmer, H.; Gram, L.; Larsen, T.O. Inhibition of virulence gene expression in Staphylococcus aureus by novel depsipeptides from a Marine photobacterium. Mar. Drugs 2011, 9, 2537–2552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamann, M.T. Technology Evaluation: Kahalalide F, PharmaMar. Curr. Opin. Mol. Ther. 2004, 6, 657–665. [Google Scholar] [PubMed]
- Fernandez, D.; Assigbese, K.; Dubois, M.P.; Geiger, J.P. Molecular characterization of races and vegetative compatibility groups in Fusarium oxysporum f. sp. Vasinfectum. Appl. Environ. Microbiol. 1994, 60, 4039–4046. [Google Scholar] [PubMed]
- Nel, B. Isolation and characterization of nonpathogenic Fusarium oxysporum isolates from the rhizosphere of healthy banana plants. Plant Pathol 2006, 55, 207–216. [Google Scholar] [CrossRef]
- Van Den Berg, N.; Berger, D.K.; Hein, I.; Birch, P.R.J.; Wingfield, M.J.; Viljoen, A. Tolerance in banana to Fusarium wilt is associated with early up-regulation of cell wall-strengthening genes in the roots. Mol. Plant Pathol. 2007, 8, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Sivamani, E. Biological control of Fusarium oxysporum f. sp. cubense in banana by inoculation with Pseudomonas fluorescens. Plant Soil 1988, 107, 3–9. [Google Scholar]
- Peng, H.X.; Sivasithamparam, K.; Turner, D.W. Chlamydospore germination and Fusarium wilt of banana plantlets in suppressive and conducive soils are affected by physical and chemical factors. Soil Biol. Biochem. 1999, 31, 1363–1374. [Google Scholar] [CrossRef]
- Thangavelu, R.; Palaniswami, A.; Velazhahan, R. Mass production of Trichoderma harzianum for managing Fusarium wilt of banana. Agric. Ecosyst. Environ. 2004, 103, 259–263. [Google Scholar] [CrossRef]
- Meenakshi, S.; Saurabh, K. Colletotrichum gloeosporioides: An Anthracnose Causing Pathogen of Fruits and Vegetables. Biosci. Biotechnol. Res. Asia 2015, 12, 1233–1246. [Google Scholar]
- Quiroz-Sarmiento, V.R.; Ferrera-Cerrato, R.; Alarcón, A.; Lara-Hernández, M.E. Antagonismo in vitro de cepas de Aspergillus y Trichoderma hacia hongos filamentosos que afectan el cultivo de ajo. Rev. Mex. Micol. 2008, 26, 27–34. [Google Scholar]
- Hasan, M.M.; Rahman, S.M.E.; Kim, G.H.; Abdallah, E.; Oh, D.H. Antagonistic Potentiality of Trichoderma harzianum Towards Seed-Borne Fungal Pathogens of Winter Wheat cv. Protiva In Vitro and In Vivo. J. Microbiol. Biotechnol. 2012, 22, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Benítez, T.; Rincón, A.M.; Limón, M.C.; Codón, A.C. Biocontrol Mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains Code | Family | Species |
|---|---|---|
| 13 | Aerococcaceae | Aerococcus urinaeequi IFO 12173 |
| 4 | Bacillaceae | Bacillus thuringiensis strain 61436 |
| 8 | Bacillaceae | Bacillus amyloliquefaciens strain PD9 |
| 12 | Bacillaceae | Bacillus subtilis |
| 16 | Bacillaceae | Bacillus aerophilus |
| 18 | Bacillaceae | Bacillus amyloliquefaciens |
| 19 | Bacillaceae | Bacillus pumilus |
| 20 | Bacillaceae | Bacillus velezensis strain Lzh-a42 |
| 21 | Bacillaceae | Bacillus sp. |
| 27 | Bacillaceae | Bacillus altitudinis |
| 31 | Bacillaceae | Bacillus anthracis ATCC 14578 |
| 33 | Bacillaceae | Bacillus aerophilus |
| 36 | Bacillaceae | Bacillus thuringiensis strain 61436 |
| 47 | Bacillaceae | Bacillus sp. strain M4 |
| 50 | Bacillaceae | Bacillus subtilis |
| 52 | Bacillaceae | Bacillus aerophilus |
| 55 | Bacillaceae | Bacillus anthracis ATCC 14578 |
| 37 | Brevibacteriaceae | Brevibacterium casei NCDO 2048 |
| 49 | Enterobacteriaceae | Serratia marcescens strain UMH2 |
| 11 | Micrococcaceae | Micrococcus sp. HW4 |
| 10 | Pseudoalteromonadaceae | Pseudoalteromonas shioyasakiensis SE3(T) |
| 46 | Shewanellaceae | Shewanella haliotis |
| 53 | Shewanellaceae | Shewanella haliotis |
| 26 | Shewanellaceae | Shewanella haliotis |
| 2 | Staphylococcaceae | Staphylococcus kloosii ATCC 43959(T) |
| 9 | Staphylococcaceae | Staphylococcus saprophyticus subsp. bovis GTC 843(T) |
| 34 | Staphylococcaceae | Staphylococcus cohnii subsp. urealyticus ATCC(49330) |
| 35 | Staphylococcaceae | Staphylococcus haemolyticus ATCC 29970 |
| 39 | Staphylococcaceae | Staphylococcus cohnii subsp. cohnii ATCC 29974 |
| 51 | Staphylococcaceae | Staphylococcus cohnii subsp. urealyticus ATCC 49330 |
| 1 | Vibrionaceae | Vibrio neocaledonicus |
| 3 | Vibrionaceae | Vibrio neocaledonicus |
| 5 | Vibrionaceae | Vibrio neocaledonicus |
| 6 | Vibrionaceae | Vibrio furnissii |
| 7 | Vibrionaceae | Vibrio alginolyticus |
| 14 | Vibrionaceae | Vibrio neocaledonicus |
| 15 | Vibrionaceae | Vibrio diabolicus |
| 17 | Vibrionaceae | Vibrio furnissii |
| 22 | Vibrionaceae | Vibrio neocaledonicus NC470 |
| 23 | Vibrionaceae | Vibrio antiquaries |
| 24 | Vibrionaceae | Vibrio alginolyticus strain 27 |
| 25 | Vibrionaceae | Vibrio neocaledonicus NC470 |
| 28 | Vibrionaceae | Vibrio furnissii |
| 29 | Vibrionaceae | Vibrio neocaledonicus |
| 30 | Vibrionaceae | Vibrio antiquaries |
| 32 | Vibrionaceae | Vibrio alginolyticus strain 27 |
| 38 | Vibrionaceae | Vibrio neocaledonicus |
| 40 | Vibrionaceae | Vibrio antiquaries |
| 41 | Vibrionaceae | Vibrio neocaledonicus NC470 |
| 42 | Vibrionaceae | Vibrio furnissii |
| 43 | Vibrionaceae | Vibrio neocaledonicus |
| 44 | Vibrionaceae | Vibrio sp. CF4-11 |
| 45 | Vibrionaceae | Vibrio neocaledonicus |
| 48 | Vibrionaceae | Vibrio neocaledonicus |
| 54 | Vibrionaceae | Vibrio neocaledonicus |
| Code | Species | %AChEI 50 µg/mL | %AChEI 25 µg/mL | %AChEI 12.5 µg/mL | %AChEI 6.25 µg/mL |
|---|---|---|---|---|---|
| 1 * | Vibrio neocaledonicus | 98.9 *** | 69.9 ** | 21.78 | 11.0 |
| 3 | Vibrio neocaledonicus | 93.0 *** | 91.1 *** | 50.2 * | 4.2 |
| 38 | Vibrio neocaledonicus | 90.8*** | 88.8 *** | 28.2 | 19.3 |
| 10 | Pseudoalteromonas shioyasakiensis SE3(T) | 86.7 *** | 84.1 *** | 21.6 | 5.2 |
| 28 | Vibrio furnissii | 89.3 *** | 79.71*** | 0 | 0 |
| 27 | Bacillus altitudinis | 83.3 *** | 78.59 *** | 79.88 *** | 30.0 |
| 49 | Serratia marcescens UMH2 | 76.4 ** | 52.1 * | 45.3 | 12.3 |
| 20 | Bacillus velezensis Lzh-a42 | 71.0 ** | 62.5 ** | 58.2 * | 4.0 |
| 46 | Shewanella haliotis | 66.7 ** | 24.49 | 10.20 | 0 |
| 2 * | Staphylococcus kloosii ATCC43959(T) | 65.0 ** | 29.51 | 31.11 | 2.4 |
| 15 | Vibrio diabolicus | 61.0 * | 63.38 ** | 45.07 | 40.1 |
| 8 | Bacillus amyloliquefaciens PD29 | 59.6 * | 54.48 * | 45.07 | 15.5 |
| 26 | Shewanella haliotis | 58.2 * | 49.95 | 45.91 | 12.3 |
| 53 | Shewanella haliotis | 58.8 * | 50.47 * | 32.04 | 19.3 |
| 19 | Bacillus pumilus | 57.0 * | 56.0 * | 48.32 | 30.3 |
| 21 | Bacillus sp. | 56.28* | 45.86 | 35.12 | 22.3 |
| 18 | Bacillus amyloliquefaciens | 55.9 * | 52.45 * | 43.71 | 9.7 |
| 23 | Vibrio antiquaries | 55.1 * | 39.49 | 19.46 | 0 |
| 13 | Aerococcus urinaeequi IFO12173 | 54.6 * | 42.15 | 34.06 | 9.1 |
| 5 | Vibrio neocaledonicus | 52.0 * | 34.54 | 12.25 | 0 |
| 30 | Vibrio antiquaries | 50.0* | 40.09 | 32.48 | 2.5 |
| 6 | Vibrio furnissii | 50.2 * | 35.74 | 33.53 | 28.9 |
| 24 | Vibrio alginolyticus | 50.9 * | 37.76 | 26.23 | 12.7 |
| C+ | Huperzine-A | 98.9% |
| Strain Code | Species | %XOI 125 µg/mL | %XOI 62.5 µg/mL | %XOI 31.25 µg/mL |
|---|---|---|---|---|
| 25 | Vibrio neocaledonicus NC470 | 31.17 | 7.56 | 2.31 |
| 3 | Vibrio neocaledonicus | 17.49 | 15.04 | 2.18 |
| 1 | Vibrio neocaledonicus | 25.97 | 25.42 | 24.31 |
| 14 | Vibrio neocaledonicus | 26.64 | 19.91 | 19.64 |
| 38 | Vibrio neocaledonicus | 27.89 | 23.63 | 5.17 |
| 10 | Pseudoalteromonas shioyasakiensis SE3(T) | 5.2 | 0 | 0 |
| 2 | Staphylococcus kloosii ATCC43959(T) | 16.34 | 10.1 | 7.81 |
| 24 | Vibrio alginolyticus | 20.96 | 8.67 | 3.63 |
| 9 | Staphylococcus saprophyticus subsp. bovis | 16.17 | 10.37 | 0 |
| 13 | Aerococcus urinaeequi IFO12173 | 23.91 | 17.55 | 0 |
| 8 | Bacillus amyloliquefaciens PD29 | 0 | 0 | 0 |
| 50 | Bacillus subtilis | 15.42 | 4.29 | 0 |
| C+ | Allopurinol | 96.37% |
| Code | Species | Alkaloids | Terpenes | Polyphenols |
|---|---|---|---|---|
| 1 | Vibrio neocaledonicus | − | + | + |
| 3 | Vibrio neocaledonicus | − | + | + |
| 38 | Vibrio neocaledonicus | − | + | + |
| 10 | Pseudoalteromonas shioyasakiensis SE3(T) | − | − | + |
| 28 | Vibrio furnissii | + | + | + |
| 27 | Bacillus altitudinis | − | − | + |
| 49 | Serratia marcescens UMH2 | − | − | + |
| 20 | Bacillus velezensis Lzh-a42 | + | + | + |
| 46 | Shewanella haliotis | − | + | + |
| 2 | Staphylococcus kloosii ATCC43959(T) | − | + | + |
| 15 | Vibrio diabolicus | − | + | + |
| 8 | Bacillus amyloliquefaciens PD29 | + | + | + |
| 26 | Shewanella haliotis | − | + | + |
| 53 | Shewanella haliotis | − | − | + |
| 19 | Bacillus pumilus | − | + | + |
| 21 | Bacillus sp. | + | − | + |
| 18 | Bacillus amyloliquefaciens | − | + | + |
| 23 | Vibrio antiquaries | − | + | + |
| 13 | Aerococcus urinaeequi IFO12173 | − | − | + |
| 5 | Vibrio neocaledonicus | − | + | + |
| 30 | Vibrio antiquaries | − | + | + |
| 6 | Vibrio furnissii | + | − | + |
| 24 | Vibrio alginolyticus | + | + | + |
| Strains Code | Inhibitory Activity | Name of Strains | |
|---|---|---|---|
| Foc4 | Cg | ||
| 5 | + | + | Vibrio neocaledonicus |
| 8 | + | + | Bacillus amyloliquefaciens strain PD9 |
| 12 | + + | + + | Bacillus subtilis |
| 16 | + | + | Bacillus aerophilus |
| 18 | + + + + | + + + + | Bacillus amyloliquefaciens |
| 19 | + + | − | Bacillus pumilus |
| 20 | + + | + + | Bacillus velezensis strain Lzh-a42 |
| 21 | + + | + + | Bacillus sp. * |
| 33 | + | − | Bacillus aerophilus |
| 47 | + + | − | Bacillus sp. strain M4 |
| 50 | + | + + + + | Bacillus subtilis |
| 52 | + + | − | Bacillus aerophilus |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, L.; Guo, S.; Ma, F.; Chang, C.; Gómez-Betancur, I. In Vitro Inhibition of Acetylcholinesterase, Alphaglucosidase, and Xanthine Oxidase by Bacteria Extracts from Coral Reef in Hainan, South China Sea. J. Mar. Sci. Eng. 2018, 6, 33. https://doi.org/10.3390/jmse6020033
Tan L, Guo S, Ma F, Chang C, Gómez-Betancur I. In Vitro Inhibition of Acetylcholinesterase, Alphaglucosidase, and Xanthine Oxidase by Bacteria Extracts from Coral Reef in Hainan, South China Sea. Journal of Marine Science and Engineering. 2018; 6(2):33. https://doi.org/10.3390/jmse6020033
Chicago/Turabian StyleTan, Lin, Suxia Guo, Funing Ma, Chen Chang, and Isabel Gómez-Betancur. 2018. "In Vitro Inhibition of Acetylcholinesterase, Alphaglucosidase, and Xanthine Oxidase by Bacteria Extracts from Coral Reef in Hainan, South China Sea" Journal of Marine Science and Engineering 6, no. 2: 33. https://doi.org/10.3390/jmse6020033