
Analysis of the Swordfish Xiphias gladius Linnaeus, 1758 Catches by the Pelagic Longline Fleets in the Eastern Pacific Ocean
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Fisheries Database
2.3. Environmental Database
2.4. Quantitative Analysis
2.5. CPUE Standardization
2.6. BFAST Algorithm
3. Results
3.1. Spatial–Temporal Variation in Fishing Effort
3.2. Space-Time Variation of Capture per Unit Effort (CPUE)
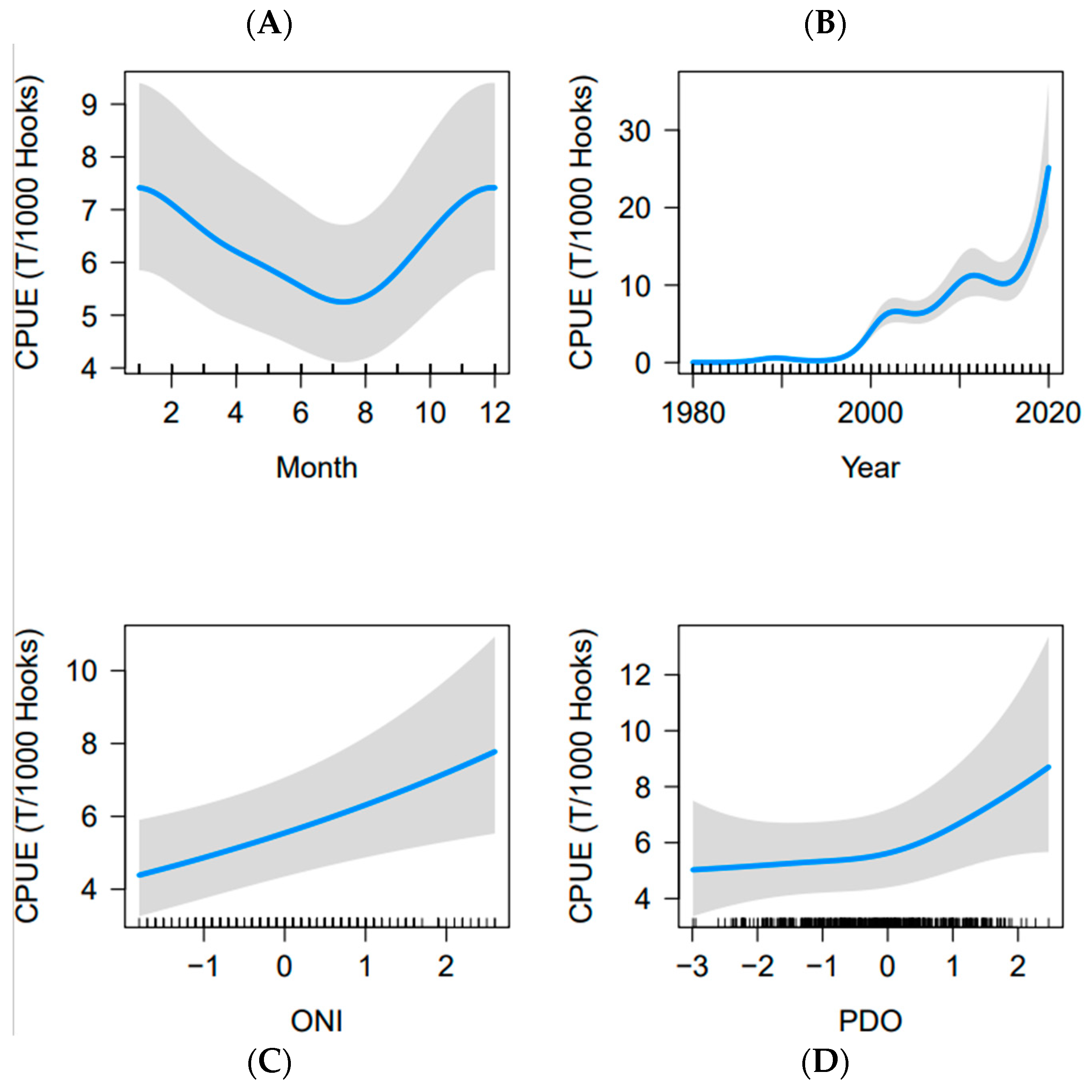
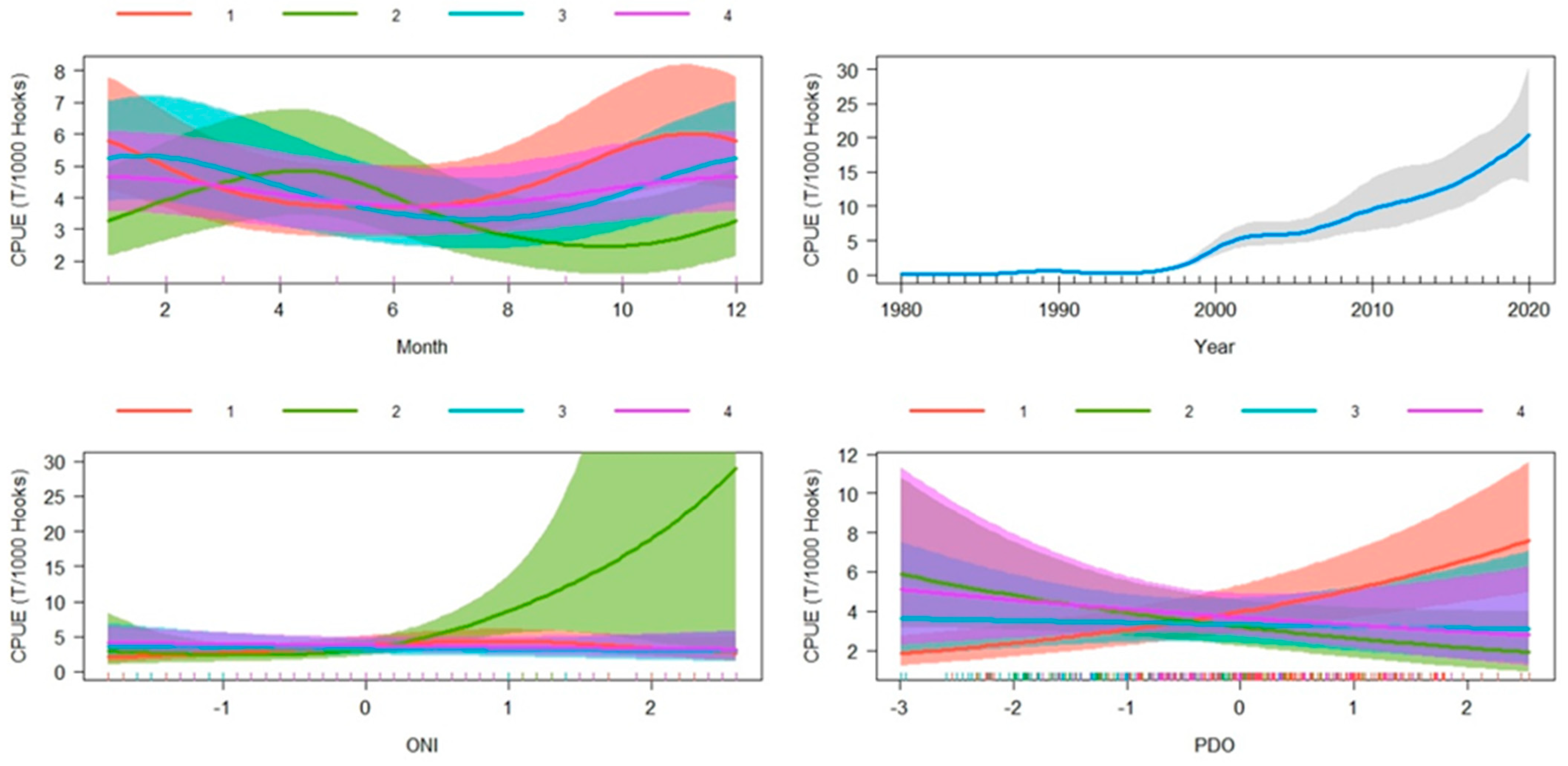
3.3. GAMM
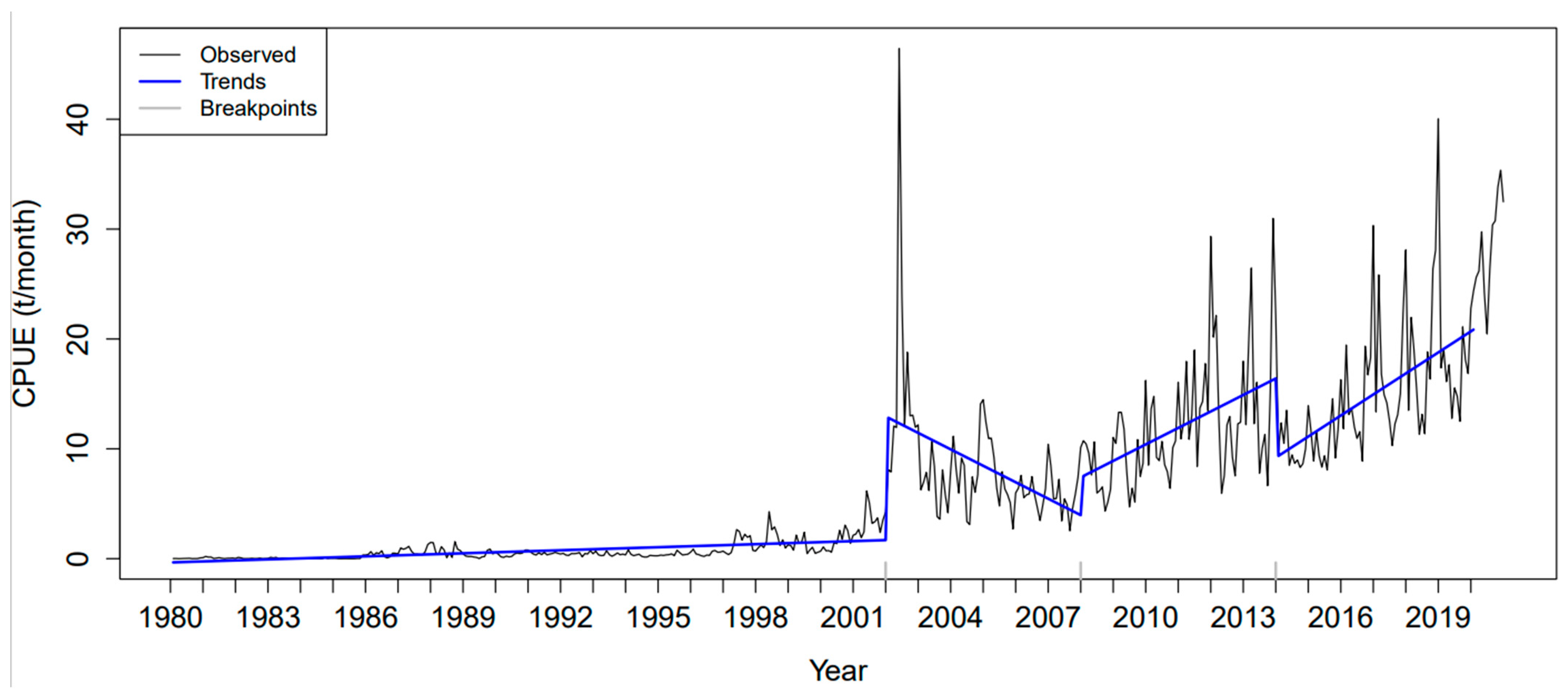
3.4. BFAST Algorithm
4. Discussion
4.1. Spatial–Temporal Variation in Fishing Effort
4.2. Spatial-Temporal Variation of the Catch per Unit of Effort CPUE
4.3. CPUE Standardization (GAMM)
4.4. CPUE Trends Obtained with the BFAST Algorithm
4.5. Possible Fishery Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

References
- Lecointre, G.; Le Guyader, G. Classification Phylogénétique du Vivant; Editions Belin: Paris, France, 2006; pp. 1–544. [Google Scholar]
- Nakamura, I. Billfishes of the World. An Annotated and Illustrated Catalogue of Marlins, Sailfishes, Spearfishes and Swordfishes Known to Date; FAO: Rome, Italy, 1985; pp. 1–65. [Google Scholar]
- Bedford, D.; Hagerman, F. The billfish fishery resources of the California Current. CALCOFI Rep. 1983, 24, 70–78. [Google Scholar]
- Takahashi, M.; Okamura, H.; Yokawa, K.; Okazaki, M. Swimming behavior and migration of a swordfish recorded by an archival tag. Mar. Fresh Res. 2003, 54, 527–534. [Google Scholar] [CrossRef]
- Lobell, M.J. The Fisheries of Chile, Present Status, and Future Possibilities (United States Fisheries Mission to Chile); U.S. Department Interior, Fish Wildlife Service: Washington, DC, USA, 1947; pp. 14–20.
- Fitch, J.E. Swordfish; California Department of Fish and Game: Sacramento, CA, USA, 1960; pp. 1–79.
- Palko, B.J.; Bearsdley, G.L.; Richards, W.J. Synopsis of the Biology of the Swordfish, Xiphias gladius Linnaeus; FAO: Rome, Italy, 1981; pp. 1–21. [Google Scholar]
- Mejuto-García, J. Aspectos Biológicos y Pesqueros del Pez Espada (Xiphias gladius Linnaeus, 1758) del Océano Atlántico, con Especial Referencia a las Áreas de Actividad de la Flota Española. Ph.D. Thesis, Universidad de Santiago de Compostela, Galicia, Spain, 2007. [Google Scholar]
- Abecassis, M.; Dewar, H.; Hawn, D.; Polovina, J. Modeling swordfish daytime vertical habitat in the North Pacific Ocean from pop-up archival tags. Mar. Ecol. Prog. Ser. 2012, 452, 219–236. [Google Scholar] [CrossRef]
- Moreira, F. Food of the swordfish, Xiphias gladius Linnaeus, 1758, off the Portuguese Coast. J. Fish. Biol. 1990, 36, 623–624. [Google Scholar] [CrossRef]
- Hernández-García, V. The diet of the swordfish Xiphias gladius Linnaeus, 1758, in the central east Atlantic, with emphasis on the role of cephalopods. Fish. Bull. 1995, 93, 403–411. [Google Scholar]
- Markaida, U.; Sosa-Nishizaki, O. Food and feeding habits of Swordfish, Xiphias gladius L., off Western Baja California. In: Biology and fisheries of swordfish Xiphias gladius. NOAA Tech. Rep. NMFS 1998, 142, 1–276. [Google Scholar]
- Trujillo-Olvera, A.; Ortega-Garcia, S.; Tripp-Valdez, A.; Escobar-Sanchez, O.; Acosta-Pachón, T.A. Feeding habits of the swordfish (Xiphias gladius Linnaeus, 1758) in the subtropical northeast Pacific. Hydrobiologia 2018, 822, 173–188. [Google Scholar] [CrossRef]
- Abascal, F.J.; Mejuto, J.; Quintans, M.; Ramos-Cartelle, A. Horizontal and vertical movements of swordfish in the Southeast Pacific. ICES J. Mar. Sci. 2010, 67, 466–474. [Google Scholar] [CrossRef]
- Sosa-Nishizaki, O. Revisión histórica del manejo de los picudos en el Pacífico mexicano. Cienc. Mar. 1998, 24, 95–111. [Google Scholar] [CrossRef]
- Fleischer, L.A.; Traulsen, A.K.; Ulloa-Ramírez, P.A. Mexican progress report on the marlin and swordfish fishery. In Proceedings of the Working Document Submitted to the ISC Billfish Working Group Workshop, Honolulu, HI, USA, 3–10 February 2009. [Google Scholar]
- Folsom, B.W.; Weidner, D.M.; Wildman, M.R. An Analysis of Swordfish Fisheries, Market Trends, and Trade Patterns Past-Present-Future; U.S. Department of Commerce: Silver Spring, MD, USA, 1997. Available online: https://repository.library.noaa.gov/view/noaa/33517 (accessed on 15 March 2024).
- Squire, J.L.; Muhlia-Melo, A.F. A review of striped marlin (Tetrapturus audax), swordfish (Xiphias gladius), and sailfish (Istiophorus platypterus) fisheries and resource management by Mexico and the United States in the northeast Pacific Ocean. NOAA NMFS SWFSC Adm. Rep. 1993, lj-93-96, 1–44. [Google Scholar]
- Marín-Enríquez, E.; Abitia-Cárdenas, L.A.; Moreno-Sánchez, X.G.; Ramírez-Pérez, J.S. Historical analysis of blue marlin (Makaira nigricans Lacepède, 1802) catches by the pelagic longline fleet in the eastern Pacific Ocean. Mar. Freshw. Res. 2020, 71, 532–541. [Google Scholar] [CrossRef]
- Wyrtki, K. An estimate of equatorial upwelling in the Pacific. Am. Meteorol. Soc. 1981, 11, 1205–1214. [Google Scholar] [CrossRef]
- Zhang, Y.; Wallace, J.M.; Battisti, D.S. ENSO-like interdecadal variability. J. Climate 1997, 10, 1004–1020. [Google Scholar] [CrossRef]
- Mantua, N.J.; Hare, S.R. The Pacific decadal oscillation. J. Oceanogr. 2002, 58, 35–44. [Google Scholar] [CrossRef]
- Newman, M.; Alexander, M.A.; Ault, T.R.; Cobb, K.M.; Deser, C.; Di Lorenzo, E.; Mantua, N.J.; Miller, A.J.; Minobe, S.; Nakamura, H.; et al. The Pacific Decadal Oscillation, Revisited. J. Climate 2016, 29, 4399–4427. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R.’; Springer: New York, NY, USA, 2009; pp. 1–458. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; pp. 1–488. [Google Scholar]
- Verbesselt, J.; Hyndman, R.; Zeileis, A.; Culvenor, D. Phenological Change Detection While Accounting for Abrupt and Gradual Trends in Satellite Image Time Series. Remote Sens. Environ. 2010, 114, 2970–2980. [Google Scholar] [CrossRef]
- Feng, Z.; Yu, W.; Chen, X. Concurrent habitat fluctuations of two economically important marine species in the Southeast Pacific Ocean off Chile in relation to ENSO perturbations. Fish. Oceanogr. 2022, 31, 123–134. [Google Scholar] [CrossRef]
- Morales-Bojorquez, E.; Pacheco-Bedoya, J.L. Jumbo squid Dosidicus gigas: A new fishery in Ecuador. Rev. Fish. Sci. Aquac. 2016, 24, 98–110. [Google Scholar] [CrossRef]
- Gilly, W.F.; Markaida, U.; Baxter, C.H.; Block, B.A.; Boustany, A.; Zeidberg, L.; Reisenbichler, K.; Robinson, B.; Bazzino, G.; Salinas, C. Vertical and horizontal migrations by jumbo squid, Dosidicus gigas, revealed by electronic tagging. Mar. Ecol. Prog. Ser. 2006, 324, 1–17. [Google Scholar] [CrossRef]
- Markaida, U.; Quiñonez-Velázquez, C.; Sosa-Nishizaki, O. Age, growth, and maturation of jumbo squid Dosidicus gigas (Cephalopoda: Ommastrephidae) from the Gulf of California, Mexico. Fish. Res. 2004, 66, 31–47. [Google Scholar] [CrossRef]
- Estrella-Arellano, C.; Swartzman, G. The Peruvian artisanal fishery: Changes in patterns and distribution over time. Fish. Res. 2010, 101, 133–145. [Google Scholar] [CrossRef]
- Bayliff, W.H. The Fisheries for Tunas in the Eastern Pacific Ocean. In Advances in Tuna Aquaculture. From Hatchery to Market; Benetti, D.D., Partridge, G.J., Buentello, A., Eds.; Academic Press: Oxford, UK, 2016; pp. 21–41. [Google Scholar]
- Neilson, J.D.; Loefer, J.; Prince, E.D.; Royer, F.; Calmettes, B.; Gaspar, P.; Lopez, R.; Andrushchenko, I. Seasonal Distributions and Migrations of Northwest Atlantic Swordfish: Inferences from Integration of Pop-Up Satellite Archival Tagging Studies. PLoS ONE 2014, 9, e112736. [Google Scholar] [CrossRef]
- Quispe-Machaca, M.; Guzmán-Rivas, F.A.; Ibañez, C.M.; Urzúa, A. Trophodynamics of the jumbo squid Dosidicus gigas during winter in the Southeast Pacific Ocean off the coast of Chile: Diet analyses and fatty acid profile. Fish. Res. 2022, 245, 106154. [Google Scholar] [CrossRef]
- Lan, K.-W.; Lee, M.-A.; Wang, S.-P.; Chen, Z.-Y. Environmental variations on swordfish (Xiphias gladius) catch rates in the Indian Ocean. Fish. Res. 2015, 166, 67–79. [Google Scholar] [CrossRef]
- Parker, D.; Winker, H.; West, W.; Kerwath, S.E. Standardization of the catch per unit effort for swordfish (Xiphias gladius) for the South African longline fishery. Sci. Pap. ICCAT 2017, 74, 1295–1305. [Google Scholar]
- Shumann, E.H.; Perris, L.A.; Hunter, I.T. Upwelling along the South Coast of the Cape Province, South Africa. S. Afr. J. Sci. 1982, 78, 238–242. [Google Scholar]
- Santana-Hernández, H.; Macías-Zamora, R.; Vidaurri-Sotelo, A. Relación entre la abundancia de peces de pico y la temperatura del agua en el Pacífico mexicano. Cienc. Pesq. 1996, 13, 62–65. [Google Scholar]
- Hinton, M.G.; Bayliff, W.H.; Suter, J.M. Assessment of swordfish in the eastern Pacific Ocean. In Proceedings of the ‘5th Meeting. Document SAR-5-05-SWO’, La Jolla, CA, USA, 11–13 May 2004; Inter-American Tropical Tuna Commission, Working Group on Stock Assessment: La Jolla, CA, USA. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | AIC | ΔAIC | p Value |
|---|---|---|---|
| s(Month) | 1298 | 392 | 0.156 |
| +s(Year) | 922 | 17 | 2 × 10−16 |
| +s(ONI) | 930 | 25 | 4.17 × 10−5 |
| +s(PDO) | 905 | 0 | 0.037 |
| Estimate | Standard Error | t Value | Pr (>|t|) | |
|---|---|---|---|---|
| Period 1 | 0.00777 | 0.00056 | 13.868 | 2.00 × 10−16 |
| Period 2 | −0.12360 | 0.03020 | −4.093 | 1.12 × 10−4 |
| Period 3 | 0.12615 | 0.02747 | 4.591 | 1.89 × 10−5 |
| Period 4 | 0.21796 | 0.02358 | 9.242 | 2.38 × 10−14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Félix-Salazar, L.A.; Marín-Enríquez, E.; Aragón-Noriega, E.A.; Ramirez-Perez, J.S. Analysis of the Swordfish Xiphias gladius Linnaeus, 1758 Catches by the Pelagic Longline Fleets in the Eastern Pacific Ocean. J. Mar. Sci. Eng. 2024, 12, 496. https://doi.org/10.3390/jmse12030496
Félix-Salazar LA, Marín-Enríquez E, Aragón-Noriega EA, Ramirez-Perez JS. Analysis of the Swordfish Xiphias gladius Linnaeus, 1758 Catches by the Pelagic Longline Fleets in the Eastern Pacific Ocean. Journal of Marine Science and Engineering. 2024; 12(3):496. https://doi.org/10.3390/jmse12030496
Chicago/Turabian StyleFélix-Salazar, Luis Adán, Emigdio Marín-Enríquez, Eugenio Alberto Aragón-Noriega, and Jorge Saul Ramirez-Perez. 2024. "Analysis of the Swordfish Xiphias gladius Linnaeus, 1758 Catches by the Pelagic Longline Fleets in the Eastern Pacific Ocean" Journal of Marine Science and Engineering 12, no. 3: 496. https://doi.org/10.3390/jmse12030496