

Deciphering Preferences for Shelter Volume and Distribution by Coral Reef Fish, Using Systematic and Functional Grouping

Abstract
:1. Introduction
2. Materials and Methods
2.1. Location and Experimental Design of the Shelters
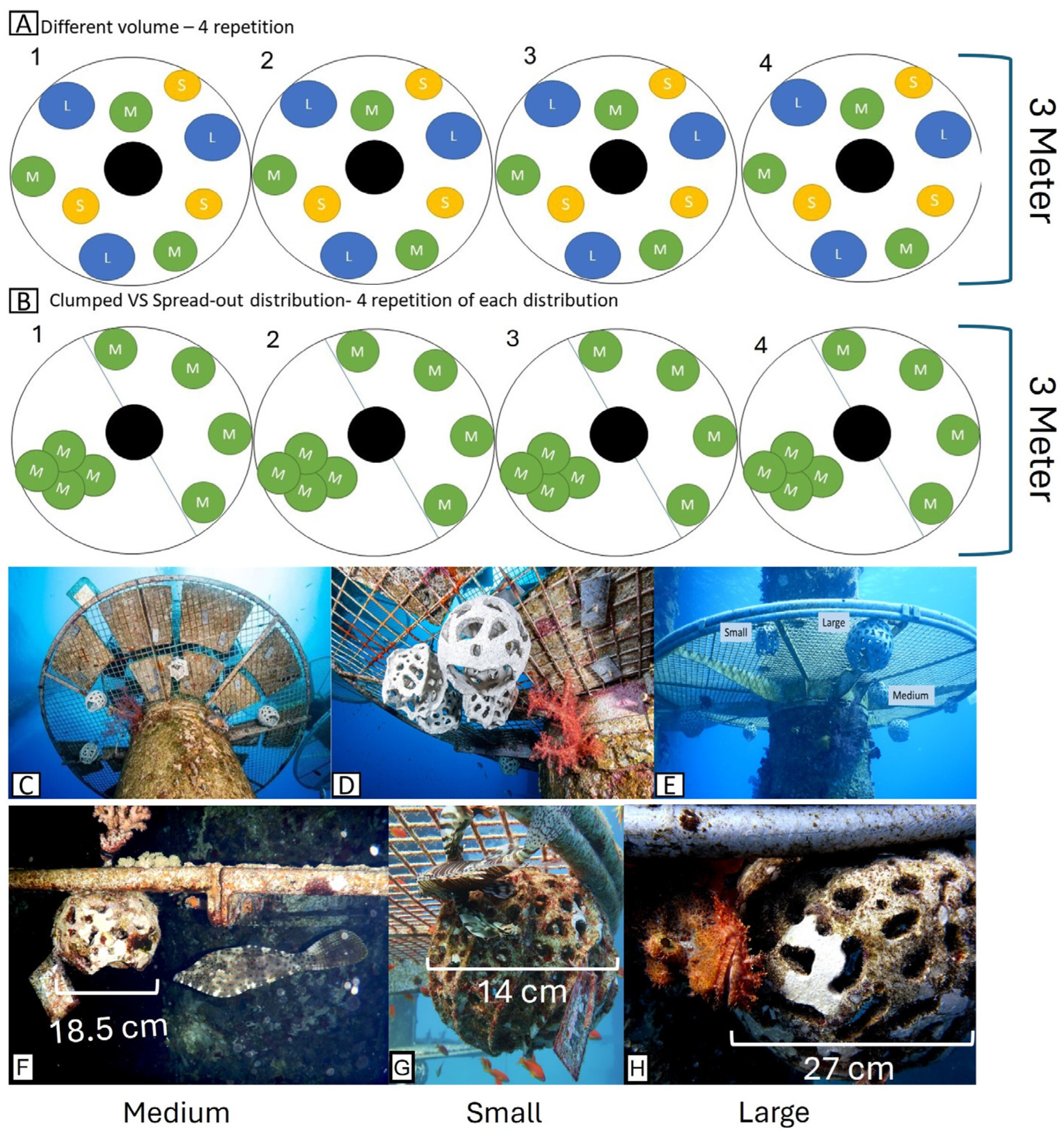
2.1.1. Experimental Shelters Design
2.1.2. Experimental Configuration for Dispersed vs. Clumped RAS
2.1.3. Experimental Configuration for the Different Sized RAS
2.2. Monitoring Procedure and Sampled Areas
Survey Technique
2.3. Data Analyses to Categorize the Different Species
2.3.1. The Classification of Species into Dietary Classes Was Conducted as Follows
2.3.2. The Species Were Categorized into Families as Follows
2.3.3. The Four-Step Analysis Includes
- For each family among the five most abundant families in the surveys, preferences for specific shelters were examined by comparing fish abundance at RAS of different sizes (using the Wilcoxon rank sum test) and fish abundance at different distributions of RAS (using the Sign test).
- The first step was repeated for each of the five dietary groups, examining specific shelters groups’ preferences. This involved comparing fish abundances across RASs of different sizes (using the Wilcoxon rank sum test) and across different distributions of RASs (using the Sign test).
- We examined 19 species. The predicted preferences were assigned in a table based on species family and dietary association for each species. The overlaying preferences of both groups (family, diet) were highlighted in the table. For example, the species Pseudanthias squamipinnis, belonging to the family Serranidae and classified as a planktivore, was predicted to have the same preferences as other species from the family Serranidae and those classified as planktivores.
- Due to the small number of individuals entering the shelters, it was not feasible to conduct a parametric statistical analysis to determine the shelter preferences of each species. Therefore, the sum of total fish numbers from all surveys conducted during the 10-month experiment for each shelter size or distribution type was used to test the predictions for each species recorded in the previous steps. The total number of fish for each species was recorded in the table and compared to family and dietary group predictions to determine whether the results were consistent with predicted preferences.
3. Results
3.1. General
3.2. Profiling Species-Specific Preferences
3.2.1. Preferences for Shelters across Different Families
3.2.2. Preferences for Shelters across Diet Groups
3.2.3. Using the Two Previous Steps to Profile the Preference of Each Species
3.2.4. Validation of Predicted Preferences against Total Number of Fish for Each Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
| Column Number | Balcony Depths (Meters) | Experiment |
| 8 | 9.5 | Dispersed vs. clumped |
| 9 | 10 11.4 | Dispersed vs. clumped Dispersed vs. clumped |
| A | 8.7 10.1 11.3 | Different sized RAS Different sized RAS Control |
| B | 8 10.6 12.5 | Different sized RAS Different sized RAS Dispersed vs. clumped |
Appendix B
| Acanthuridae 1. Acanthurus nigrofuscus/Ctenochaetus striatus 2. Zebrasoma desjardinii 3. Zebrasoma xanthurum Antennariidae 4. Antennatus coccineus Apogonidae 5. Apogon erythrosoma 6. Cheilodipterus novemstriatus 7. Ostorhinchus cyanosoma Balistidae 8. Sufflamen albicaudatus Blenniidae 9. Ecsenius dentex 10. Ecsenius frontalis 11. Ecsenius gravieri 12. Ecsenius midas 13. Mimoblennius cirrosus 14. Plagiotremus tapeinosoma 15. Blenniidae Sp. Caesionidae (Fusiliers) 16. Caesio lunaris/Caesio suevica Chaetodontidae (Butterflyfishes) 17. Chaetodon auriga 18. Chaetodon austriacus 19. Chaetodon fasciatus 20. Heniochus intermedius 21. Chaetodon paucifasciatus 22. Chaetodon trifascialis Congridae 23. Conger cinereus Gobiidae 24. Eviota guttata 25. Gobiidae familly (Unknown) 26. Pleurosicya micheli Holocentridae 27. Myripristis murdjan 28. Neoniphon sammara 29. Sargocentron diadema 30. Sargocentron rubrum Kyphosidae 31. Kyphosus cinerascens/Kyphosus vaigiensis Labridae (Wrasses) 32. Bodianus anthioides 33. Bodianus diana 34. Cheilinus abudjubbe 35. Cheilinus lunulatus 36. Coris aygula 37. Labroides dimidiatus 38. Larabicus quadrilineatus 39. Oxycheilinus mentalis 40. Pseudocheilinus hexataenia 41. Thalassoma lunare 42. Thalassoma rueppellii Lethrinidae 43. Monotaxis grandoculis Monacanthidae 44. Aluterus scriptus 45. Cantherhines pardalis Mullidae 46. Parupeneus cyclostomus 47. Parupeneus forsskali/Parupeneus macronema Muraenidae 48. Gymnothorax flavimarginatus 49. Gymnothorax griseus 50. Gymnothorax pharaoensis 51. Muraenidae sp. Ostraciidae 52. Ostracion cubicus Pempheridae 53. Pempheris vanicolensis Pomacanthidae (Angelfishes) 54. Apolemichthys xanthotis 55. Pomacanthus imperator Pomacentridae (Damselfishes) 56. Abudefduf vaigiensis 57. Amblyglyphidodon flavilatus 58. Amblyglyphidodon indicus 59. Chromis viridis 60. Chromis weberi 61. Dascyllus trimaculatus 62. Marginate dascyllus 63. Neopomacentrus miryae 64. Pomacentrus trichrourus Pseudochromidae (Dottybacks) 65. Pseudochromis fridmani 66. Pseudochromis olivaceus 67. Pseudochromis springeri Scaridae (Parrotfishes) 68. Calotomus viridescens 69. Chlorurus gibbus 70. Chlorurus sordidus 71. Scarus ferrugineus 72. Scarus niger Scorpaenidae 73. Pterois miles 74. Scorpaenodes corallinus 75. Scorpaenopsis oxycephala Serranidae 76. Aethaloperca rogaa 77. Cephalopholis argus 78. Cephalopholis hemistiktos 79. Cephalopholis miniata 80. Epinephelus fasciatus 81. Pseudanthias squamipinnis 82. Pseudanthias taeniatus Siganidae 83. Siganus argenteus 84. Siganus rivulatus 85. Siganus stellatus Sparidae 86. Diplodus noct Syngnathidae 87. Corythoichthys flavofasciatus/Corythoichthys schultzi Synodontidae 88. Synodus variegatus Tetraodontidae 89. Arothron diadematus 90. Arothron hispidus 91. Canthigaster margaritata Tripterygiidae 92. Norfolkia brachylepis |
Appendix C
| Pomacentridae (N = 9) | Serranidae (N = 7) | Labridae (N = 11) | Acanthuridae (N = 3) | Scaridae (N = 5) |
|
|
|
|
|
Appendix D
| Planktivore (N = 8) | Corallivore (N = 2) | Herbivore (N = 9) | Benthivore (N = 14) | Piscivore (N = 8) |
|
|
|
|
|
References
- Gil-Agudelo, D.L.; Cintra-Buenrostro, C.E.; Brenner, J.; González-Díaz, P.; Kiene, W.; Lustic, C.; Pérez-España, H. Coral Reefs in the Gulf of Mexico Large Marine Ecosystem: Conservation Status, Challenges, and Opportunities. Front. Mar. Sci. 2020, 6, 807. [Google Scholar] [CrossRef]
- Brathwaite, A.; Clua, E.; Roach, R.; Pascal, N. Coral Reef Restoration for Coastal Protection: Crafting Technical and Financial Solutions. J. Environ. Manag. 2022, 310, 114718. [Google Scholar] [CrossRef] [PubMed]
- Barbier, E.B. Marine Ecosystem Services. Curr. Biol. 2017, 27, R507–R510. [Google Scholar] [CrossRef] [PubMed]
- Coker, D.J.; Wilson, S.K.; Pratchett, M.S. Importance of Live Coral Habitat for Reef Fishes. Rev. Fish Biol. Fish. 2014, 24, 89–126. [Google Scholar] [CrossRef]
- Nash, K.L.; Graham, N.A.J.; Wilson, S.K.; Bellwood, D.R. Cross-Scale Habitat Structure Drives Fish Body Size Distributions on Coral Reefs. Ecosystems 2013, 16, 478–490. [Google Scholar] [CrossRef]
- Levy, G.; Shaish, L.; Haim, A.; Rinkevich, B. Mid-Water Rope Nursery-Testing Design and Performance of a Novel Reef Restoration Instrument. Ecol. Eng. 2010, 36, 560–569. [Google Scholar] [CrossRef]
- Pratchett, M.S.; Hoey, A.S.; Wilson, S.K. Reef Degradation and the Loss of Critical Ecosystem Goods and Services Provided by Coral Reef Fishes. Curr. Opin. Environ. Sustain. 2014, 7, 37–43. [Google Scholar] [CrossRef]
- McCormick, M.I.; Chivers, D.P.; Allan, B.J.M.; Ferrari, M.C.O. Habitat Degradation Disrupts Neophobia in Juvenile Coral Reef Fish. Glob. Chang. Biol. 2017, 23, 719–727. [Google Scholar] [CrossRef]
- Rinkevich, B. Management of Coral Reefs: We Have Gone Wrong When Neglecting Active Reef Restoration. Mar. Pollut. Bull. 2008, 56, 1821–1824. [Google Scholar] [CrossRef]
- Chabanet, P.; Ralambondrainy, H.; Amanieu, M.; Faure, G.; Galzin, R. Relationships between Coral Reef Substrata and Fish. Coral Reefs 1997, 16, 93–102. [Google Scholar] [CrossRef]
- Carral, L.; Lamas, M.I.; Barros, J.J.C.; López, I.; Carballo, R. Proposed Conceptual Framework to Design Artificial Reefs Based on Particular Ecosystem Ecology Traits. Biology 2022, 11, 680. [Google Scholar] [CrossRef] [PubMed]
- Rinkevich, B. Conservation of Coral Reefs through Active Restoration Measures: Recent Approaches and Last Decade Progress. Environ. Sci. Technol. 2005, 39, 4333–4342. [Google Scholar] [CrossRef] [PubMed]
- Rinkevich, B. Climate Change and Active Reef Restoration-Ways of Constructing the “Reefs of Tomorrow”. J. Mar. Sci. Eng. 2015, 3, 111–127. [Google Scholar] [CrossRef]
- Dehnert, I.; Saponari, L.; Galli, P.; Montano, S. Comparing Different Farming Habitats for Mid-Water Rope Nurseries to Advance Coral Restoration Efforts in the Maldives. PeerJ 2022, 10, e12874. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, T.; Ding, G.; Yu, D.; Yang, W.; Sun, Q.; Wang, X.; Lin, H. Moderate Relative Size of Covered and Non-Covered Structures of Artificial Reef Enhances the Sheltering Effect on Reef Fish. Front. Mar. Sci. 2023, 10, 1130626. [Google Scholar] [CrossRef]
- Higgins, E.; Metaxas, A.; Scheibling, R.E. A Systematic Review of Artificial Reefs as Platforms for Coral Reef Research and Conservation. PLoS ONE 2022, 17, e0261964. [Google Scholar] [CrossRef] [PubMed]
- Belhassen, Y.; Rousseau, M.; Tynyakov, J.; Shashar, N. Evaluating the Attractiveness and Effectiveness of Artificial Coral Reefs as a Recreational Ecosystem Service. J. Environ. Manag. 2017, 203, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Polak, O.; Shashar, N. Can a Small Artificial Reef Reduce Diving Pressure from a Natural Coral Reef? Lessons Learned from Eilat, Red Sea. Ocean Coast Manag. 2012, 55, 94–100. [Google Scholar] [CrossRef]
- Shani, A.; Polak, O.; Shashar, N. Artificial Reefs and Mass Marine Ecotourism. Tour. Geogr. 2012, 14, 361–382. [Google Scholar] [CrossRef]
- Doligez, B.; Boulinier, T. Habitat Selection and Habitat Suitability Preferences. In Encyclopedia of Ecology; Elsevier: Amsterdam, The Netherlands, 2008; pp. 1810–1830. [Google Scholar]
- Fretwell, S.D.; Lucas, H.L. On Territorial Behavior and Other Factors Influencing Habitat Distribution in Birds. Acta Biotheor 1968, 19, 16–36. [Google Scholar] [CrossRef]
- Stewart, A.; Komers, P.E. Testing the Ideal Free Distribution Hypothesis: Moose Response to Changes in Habitat Amount. ISRN Ecol. 2012, 2012, 945209. [Google Scholar] [CrossRef]
- Rilov, G.; Benayahub, Y. Vertical Artificial Structures as an Alternative Habitat for Coral Reef Fishes in Disturbed Environments. Mar. Environ. Res. 1998, 45, 431451. [Google Scholar] [CrossRef]
- Holbrook, S.J.; Brooks, A.J.; Schmitt, R.J. Predictability of Fish Assemblages on Coral Patch Reefs. Mar. Freshw. Res. 2002, 53, 181–188. [Google Scholar] [CrossRef]
- Khalaf, M.A.; Kochzius, M. Changes in Trophic Community Structure of Shore Fishes at an Industrial Site in the Gulf of Aqaba Red Sea. Mar. Ecol. Prog. Ser. 2002, 239, 287–299. [Google Scholar] [CrossRef]
- Polak, O.; Polak, T.; Shashar, N. Effects of Habitat Relocation on Fish Community Structure- a Case Study. Environ. Biol. Fishes 2014, 97, 1335–1345. [Google Scholar] [CrossRef]
- Hixon, M.A.; Beets, J.P. Predation, Prey Refuges, and the Structure of Coral-Reef Fish Assemblages. Ecol. Monogr. 1993, 63, 77–101. [Google Scholar] [CrossRef]
- Julian Caley, M.; St John, J.; John, J.S. Refuge Availability Structures Assemblages of Tropical Reef Fishes. J. Anim. Ecol. 1996, 65, 414–428. [Google Scholar] [CrossRef]
- Eggleston, D.B.; Lipcius, R.N.; Grover, J.J. Predator and Shelter-Size Effects on Coral Reef Fish and Spiny Lobster Prey. Mar. Ecol. Prog. Ser. 1997, 149, 43–59. [Google Scholar] [CrossRef]
- Friedlander, A.M.; Parrish, J.D. Habitat Characteristics Affecting Fish Assemblages on a Hawaiian Coral Reef. J. Exp. Mar. Biol. Ecol. 1998, 224, 1–30. [Google Scholar] [CrossRef]
- Emslie, M.J.; Cheal, A.J.; Sweatman, H.; Delean, S. Recovery from Disturbance of Coral and Reef Fish Communities on the Great Barrier Reef, Australia. Mar. Ecol. Prog. Ser. 2008, 371, 177–190. [Google Scholar] [CrossRef]
- Bejarano, S.; Mumby, P.J.; Sotheran, I. Predicting Structural Complexity of Reefs and Fish Abundance Using Acoustic Remote Sensing (RoxAnn). Mar. Biol. 2011, 158, 489–504. [Google Scholar] [CrossRef]
- Hixon, M.A.; Beets, J.P. Shelter Characteristics and Caribbean Fish Assemblages Experiments with Artificial Reefs. Bull. Mar. Sci. 1989, 44, 666–680. [Google Scholar]
- Vega Fernández, T.; D’Anna, G.; Badalamenti, F.; Pérez-Ruzafa, A. Habitat Connectivity as a Factor Affecting Fish Assemblages in Temperate Reefs. Aquat. Biol. 2007, 1, 239–248. [Google Scholar] [CrossRef]
- Belmaker, J.; Ziv, Y.; Shashar, N. The Influence of Connectivity on Richness and Temporal Variation of Reef Fishes. Landsc. Ecol. 2011, 26, 587–597. [Google Scholar] [CrossRef]
- Lemoine, H.R.; Paxton, A.B.; Anisfeld, S.C.; Claire, R.; Peterson, C.H. Selecting the Optimal Artificial Reefs to Achieve Fish Habitat Enhancement Goals. In Biological Conservation; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Paxton, A.B.; Shertzer, K.W.; Bacheler, N.M.; Kellison, G.T.; Riley, K.L.; Taylor, J.C. Meta-Analysis Reveals Artificial Reefs Can Be Effective Tools for Fish Community Enhancement but Are Not One-Size-Fits-All. Front. Mar. Sci. 2020, 7, 282. [Google Scholar] [CrossRef]
- Belmaker, J.; Ziv, Y.; Shashar, N. Habitat Patchiness and Predation Modify the Distribution of a Coral-Dwelling Damselfish. Mar. Biol. 2009, 156, 447–454. [Google Scholar] [CrossRef]
- McCord, C.L.; Nash, C.M.; Cooper, W.J.; Westneat, M.W. Phylogeny of the Damselfishes (Pomacentridae) and Patterns of Asymmetrical Diversification in Body Size and Feeding Ecology. PLoS ONE 2021, 16, e0258889. [Google Scholar] [CrossRef]
- Bull, A.S.; Love, M.S. Worldwide Oil and Gas Platform Decommissioning: A Review of Practices and Reefing Options. Ocean Coast Manag. 2019, 168, 274–306. [Google Scholar] [CrossRef]
- Macreadie, P.I.; Fowler, A.M.; Booth, D.J. Rigs-to-Reefs: Will the Deep Sea Benefit from Artificial Habitat? Front. Ecol. Environ. 2011, 9, 455–461. [Google Scholar] [CrossRef]
- Golani, D.; Diamant, A. Fish Colonization of an Artificial Reef in the Gulf of Elat, Northern Red Sea. Environ. Biol. Fishes 1999, 54, 275–282. [Google Scholar] [CrossRef]
- Belmaker, J.; Ben-Moshe, N.; Ziv, Y.; Shashar, N. Determinants of the Steep Species-Area Relationship of Coral Reef Fishes. Coral Reefs 2007, 26, 103–112. [Google Scholar] [CrossRef]
- Caldwell, Z.R.; Zgliczynski, B.J.; Williams, G.J.; Sandin, S.A. Reef Fish Survey Techniques: Assessing the Potential for Standardizing Methodologies. PLoS ONE 2016, 11, e0153066. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Froese, R.; Pauly, D. FishBase. Available online: https://fishbase.mnhn.fr/search.php (accessed on 18 August 2020).
- Lieske, E.; Myers, R.F. Coral Reef Guide Red Sea; Collins: New York, NY, USA, 2004. [Google Scholar]
- Debelius, H. Red Sea Reef Guide, 5th ed.; Hollywood Import & Export Inc.: Gainesville, FL, USA, 2011. [Google Scholar]
- Bailey-Brock, J.H. Fouling Community Development on an Artificial Reef in Hawaiian Waters. Bull. Mar. Sci. 1989, 44, 580–591. [Google Scholar]
- Nadler, L.E.; McNeill, D.C.; Alwany, M.A.; Bailey, D.M. Effect of Habitat Characteristics on the Distribution and Abundance of Damselfish within a Red Sea Reef. Environ. Biol. Fishes 2014, 97, 1265–1277. [Google Scholar] [CrossRef]
- Khalaf, M.A.; Al-Horani, F.A.; Al-Rousan, S.A.; Manasrah, R.S. Community Structure of the Family Pomacentridae along the Jordanian Coast, Gulf of Aqaba, Red Sea. Zool. Middle East 2006, 37, 47–62. [Google Scholar] [CrossRef]
- Roberts, C.M.; Ormond, R.F.G. Habitat Complexity and Coral Reef Fish Diversity and Abundance on Red Sea Fringing Reefs. Mar. Ecol. Prog. Ser. 1987, 41, 1–8. [Google Scholar] [CrossRef]
- Hackradt, C.W.; Félix-Hackradt, F.C.; García-Charton, J.A. Influence of Habitat Structure on Fish Assemblage of an Artificial Reef in Southern Brazil. Mar. Environ. Res. 2011, 72, 235–247. [Google Scholar] [CrossRef]
- Berumen, M.L.; Hoey, A.S.; Bass, W.H.; Bouwmeester, J.; Catania, D.; Cochran, J.E.M.; Khalil, M.T.; Miyake, S.; Mughal, M.R.; Spaet, J.L.Y.; et al. The Status of Coral Reef Ecology Research in the Red Sea. Coral Reefs 2013, 32, 737–748. [Google Scholar] [CrossRef]
- Pitcher, T.J. (Ed.) The Behaviour of Teleost Fishes, 1st ed.; Springer: New York, NY, USA, 1986. [Google Scholar]
- Li, L.; Ma, Z. Species Sorting and Neutral Theory Analyses Reveal Archaeal and Bacterial Communities Are Assembled Differently in Hot Springs. Front. Bioeng. Biotechnol. 2020, 8, 464. [Google Scholar] [CrossRef]
- Oliveira, M.T.; Ramos, J.; Santos, M.N. An Approach to the Economic Value of Diving Sites: Artificial versus Natural Reefs off Sal Island, Cape Verde. J. Appl. Ichthyol. 2015, 31, 86–95. [Google Scholar] [CrossRef]
- Seaman, W. (Ed.) Artificial Reef Evaluation: With Application to Natural Marine Habitats, 1st ed.; CRC Press: Boca Raton, FL, USA, 2000; ISBN 9780429124495. [Google Scholar]
- Polak, O.; Shashar, N. Economic Value of Biological Attributes of Artificial Coral Reefs. ICES J. Mar. Sci. 2013, 70, 904–912. [Google Scholar] [CrossRef]
- Firth, L.B.; Farnworth, M.; Fraser, K.P.P.; McQuatters-Gollop, A. Make a Difference: Choose Artificial Reefs over Natural Reefs to Compensate for the Environmental Impacts of Dive Tourism. Sci. Total Environ. 2023, 901, 165488. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Families | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Pomacentridae | Serranidae | Labridae | Acanthuridae | Scaridae | ||||||
| Day | Night | Day | Night | Day | Night | Day | Night | Day | Night | |
| Shelter size preferences | L > S M > S | NA | NA | NA | M > S M > L | NA | NA | NA | M > S | NA |
| Dispersal type preferences | NA | Clumped > Dispersed | Clumped > Dispersed | Clumped > Dispersed | NA | Dispersed > Clumped | Dispersed > Clumped | NA | NA | NA |
| Diet | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Planktivore | Benthivore | Herbivore | Piscivore | Corallivore | ||||||
| Day | Night | Day | Night | Day | Night | Day | Night | Day | Night | |
| Shelter size preferences | L > S | NA | M > S | NA | M > S | NA | M > L | NA | NA | NA |
| Dispersal type preferences | NA | NA | NA | Dispersed > Clumped | Dispersed > Clumped | NA | NA | NA | NA | NA |
| Pseudocheilinus hexataenia | |||
|---|---|---|---|
| Labridae | Benthivore | ||
| Day | Night | Day | Night |
| M > S M > L | Dispersed > Clumped | M > S | Dispersed > Clumped |
| Species Name | Family | Dietary | Survey Results |
|---|---|---|---|
| Abudefduf vaigiensis | Pomacentridae | Planktivore | |
| Day | Large > Small | Large > Small | Large (37) > Medium (27) > Small (1) |
| Medium > Small | Dispersed (16) > Clumped (9) | ||
| Night | Dispersed > Clumped | No significant preferences | Medium (5) > Large (3) > Small (0) |
| Clumped (1) > Dispersed (0) | |||
| Pseudanthias | Serranidae | Planktivore | |
| squamipinnis | |||
| Clumped > Dispersed | Large > Small | Large (199) > Medium (132) > Small (98) | |
| Clumped (38) > Dispersed (13) | |||
| Clumped > Dispersed | No significant preferences | Medium (4) > Large (2) > Small (1) | |
| Clumped (10) > Dispersed (1) | |||
| Acanthurus nigrofuscus/Ctenochaetus striatus | Acanthuridae | Herbivore | |
| Day | Dispersed > Clumped | Dispersed > Clumped | Large (9) > Medium (7) > Small (6) |
| Medium > Small | Dispersed (36) > Clumped (14) | ||
| Night | No presence | No presence | No presence |
| Bodianus anthioides | Labridae | Benthivore | |
| Day | Medium > Small | Medium > Small | Large (2) > Medium (0) = Small (0) |
| Medium > Large | Dispersed (3) = Clumped (3) | ||
| Night | Dispersed > Clumped | Dispersed > Clumped | Large (0) = Medium (0) = Small (0) |
| Dispersed (19) > Clumped (7) | |||
| Oxycheilinus mentalis | Labridae | Piscivore | |
| Day | Medium > Small | Medium > Large | Medium (10) > Small (5) > Large (1) |
| Medium > Large | Dispersed (13) < Clumped (6) | ||
| Night | Dispersed > Clumped | No significant presence | Large (0) = Medium (0) = Small (0) |
| Dispersed (3) > Clumped (2) | |||
| Pomacentrus trichrourus | Pomacentridae | Planktivore | |
| Day | Medium > Small | Large > Small | Medium (1) > Large (0) = Small (0) |
| Large > Small | Clumped (6) > Dispersed (0) | ||
| Night | Dispersed > Clumped | No significant preferences | Large (2) > Medium (1) > Small (0) |
| Clumped (1) = Dispersed (1) | |||
| Thalassoma lunare | Labridae | Benthivore | |
| Day | Medium > Small | Medium > Small | Medium (19) > Large (17) > Small (8) |
| Medium > Large | Dispersed (8) < Clumped (5) | ||
| Night | Dispersed > Clumped | Dispersed > Clumped | No presence |
| Neopomacentrus miryae | Pomacentridae | Planktivore | |
| Day | Medium > Small | Large > Small | Large (47) > Medium (29) = Small (22) |
| Large > Small | Dispersed (8) > Clumped (1) | ||
| Night | Dispersed > Clumped | No significant preferences | Medium (67) > Large (64) = Small (59) |
| Dispersed (65) > Clumped (38) | |||
| Thalassoma rueppellii | Labridae | Benthivore | |
| Day | Medium > Small | Medium > Small | Medium (56) > Large (37) > Small (28) |
| Medium > Large | Dispersed (36) < Clumped (24) | ||
| Night | Dispersed > Clumped | Dispersed > Clumped | No presence |
| Scarus ferrugineus | Scaridae | Herbivore | |
| Day | Medium > Small | Dispersed > Clumped | Medium (8) = Large (8) > Small (1) |
| Medium > Small | Dispersed (6) < Clumped (1) | ||
| Night | No presence | No presence | No presence |
| Zebrasoma xanthurum | Acanthuridae | Herbivore | |
| Day | Dispersed > Clumped | Dispersed > Clumped | Small (1) > Large (0) = Medium (0) |
| Medium > Small | Clumped (1) > Dispersed (0) | ||
| Night | No presence | No presence | No presence |
| Cephalopholis miniata | Serranidae | Piscivore | |
| Day | Clumped > Dispersed | Medium > Large | No presence |
| Night | Clumped > Dispersed | No significant presence | No presence |
| Pseudocheilinus | Labridae | Benthivore | |
| Hexataenia | |||
| Day | Medium > Small | Day | Large (1) > Medium (0) = Small (0) |
| Medium > Large | Medium > Small | Dispersed (0) = Clumped (0) | |
| Night | Dispersed > Clumped | Dispersed > Clumped | No presence |
| Amblyglyphidodon indicus | Pomacentridae | Planktivore | |
| Day | Medium > Small | Large > Small | Large (0) = Medium (0) = Small (0) |
| Large > Small | Dispersed (13) > Clumped (7) | ||
| Night | Dispersed > Clumped | No significant preferences | Large (1) > Medium (0) = Small (0) |
| Clumped (2) > Dispersed (1) | |||
| Bodianus diana | Labridae | Benthivore | |
| Day | Medium > Small | Medium > Small | Large (1) > Medium (0) = Small (0) |
| Medium > Large | Dispersed (2) > Clumped (1) | ||
| Night | Dispersed > Clumped | Dispersed > Clumped | No presence |
| Aethaloperca rogaa | Serranidae | Piscivore | |
| Day | Clumped > Dispersed | Medium > Large | Small (1) > Large (0) = Medium (0) |
| Clumped (1) > Dispersed (0) | |||
| Night | Clumped > Dispersed | No significant presence | No presence |
| Scarus niger | Scaridae | Herbivore | |
| Day | Medium > Small | Dispersed > Clumped | Medium (1) = Large (1) > Small (0) |
| Medium > Small | Dispersed (0) = Clumped (0) | ||
| Night | No presence | No presence | No presence |
| Calotomus viridescens | Scaridae | Herbivore | |
| Day | Medium > Small | Dispersed > Clumped | Medium (7) > Small (3) > Large (0) |
| Medium > Small | Dispersed (3) > Clumped (2) | ||
| Night | No presence | No presence | No presence |
| Larabicus quadrilineatus | Labridae | Corallivore | |
| Day | Medium > Small | Not enough data | No presence |
| Medium > Large | |||
| Night | Dispersed > Clumped | Not enough data | No presence |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shabi, T.; Ziv, Y.; Yosef, R.; Shashar, N. Deciphering Preferences for Shelter Volume and Distribution by Coral Reef Fish, Using Systematic and Functional Grouping. J. Mar. Sci. Eng. 2024, 12, 186. https://doi.org/10.3390/jmse12010186
Shabi T, Ziv Y, Yosef R, Shashar N. Deciphering Preferences for Shelter Volume and Distribution by Coral Reef Fish, Using Systematic and Functional Grouping. Journal of Marine Science and Engineering. 2024; 12(1):186. https://doi.org/10.3390/jmse12010186
Chicago/Turabian StyleShabi, Tamar, Yaron Ziv, Reuven Yosef, and Nadav Shashar. 2024. "Deciphering Preferences for Shelter Volume and Distribution by Coral Reef Fish, Using Systematic and Functional Grouping" Journal of Marine Science and Engineering 12, no. 1: 186. https://doi.org/10.3390/jmse12010186