
Coastal Fish Fauna in the Cystoseira s.l. Algal Belts: Experiences from the Northern Adriatic Sea

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
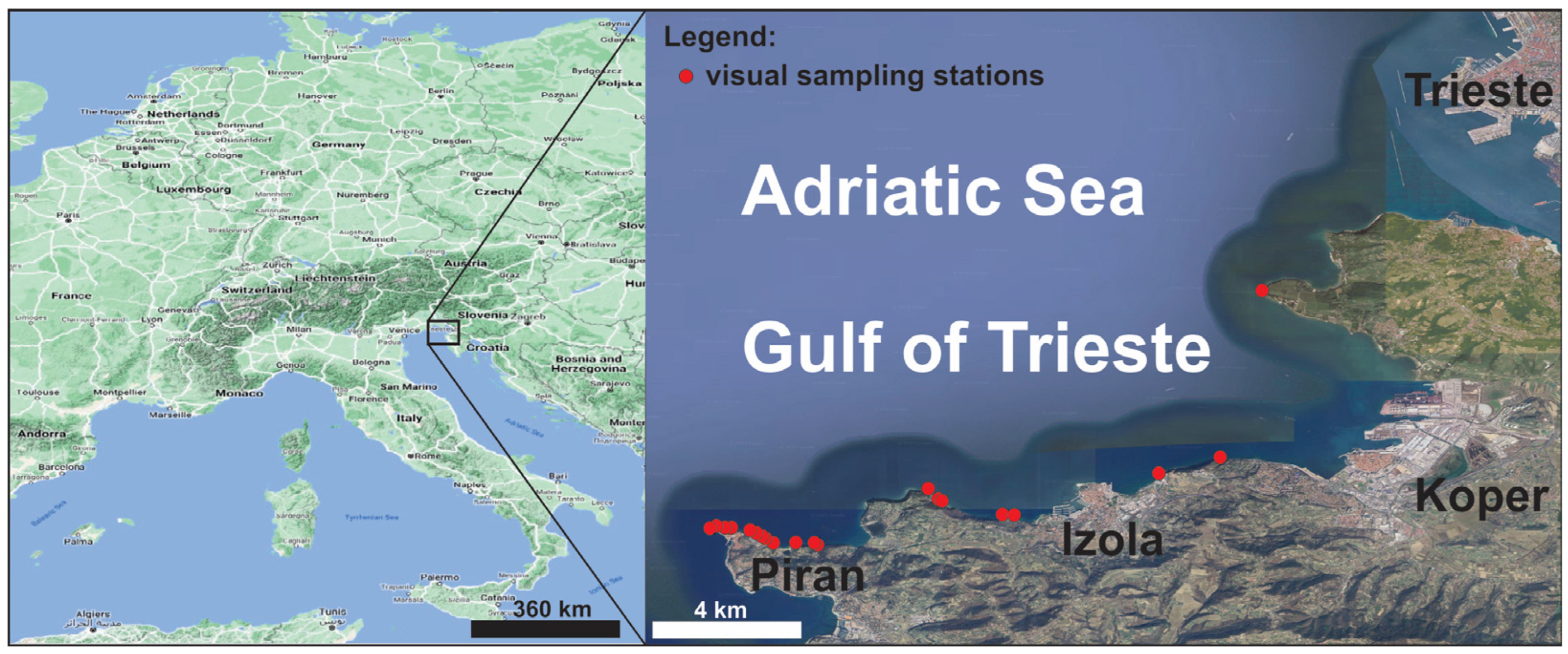
2.1. Study Area
2.2. Fieldwork
2.2.1. Fish Assemblage Composition and Abundance
2.2.2. Habitat Types
2.3. Data Analysis
3. Results
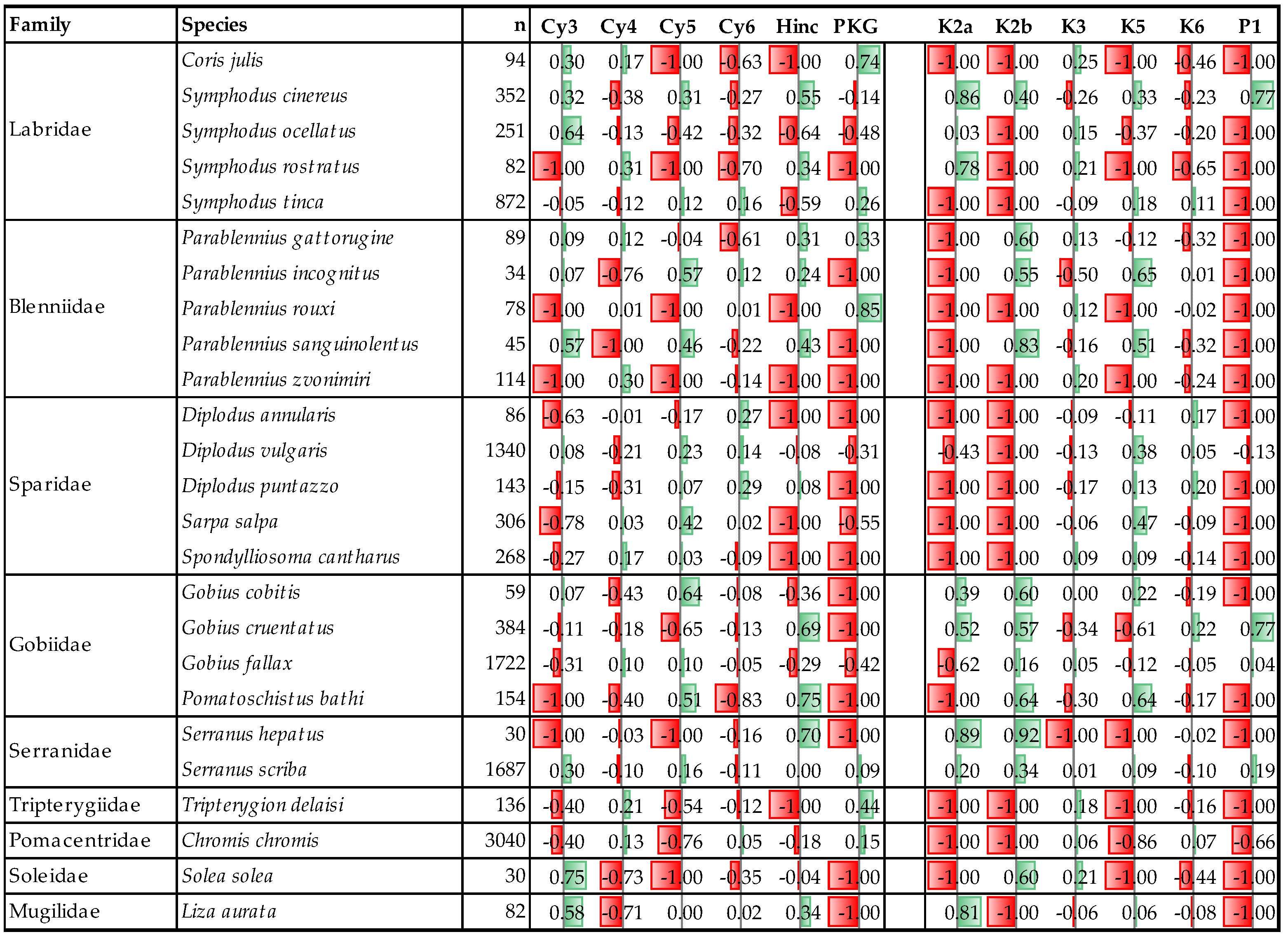
3.1. Coastal Fish Assemblage Composition and Abundance
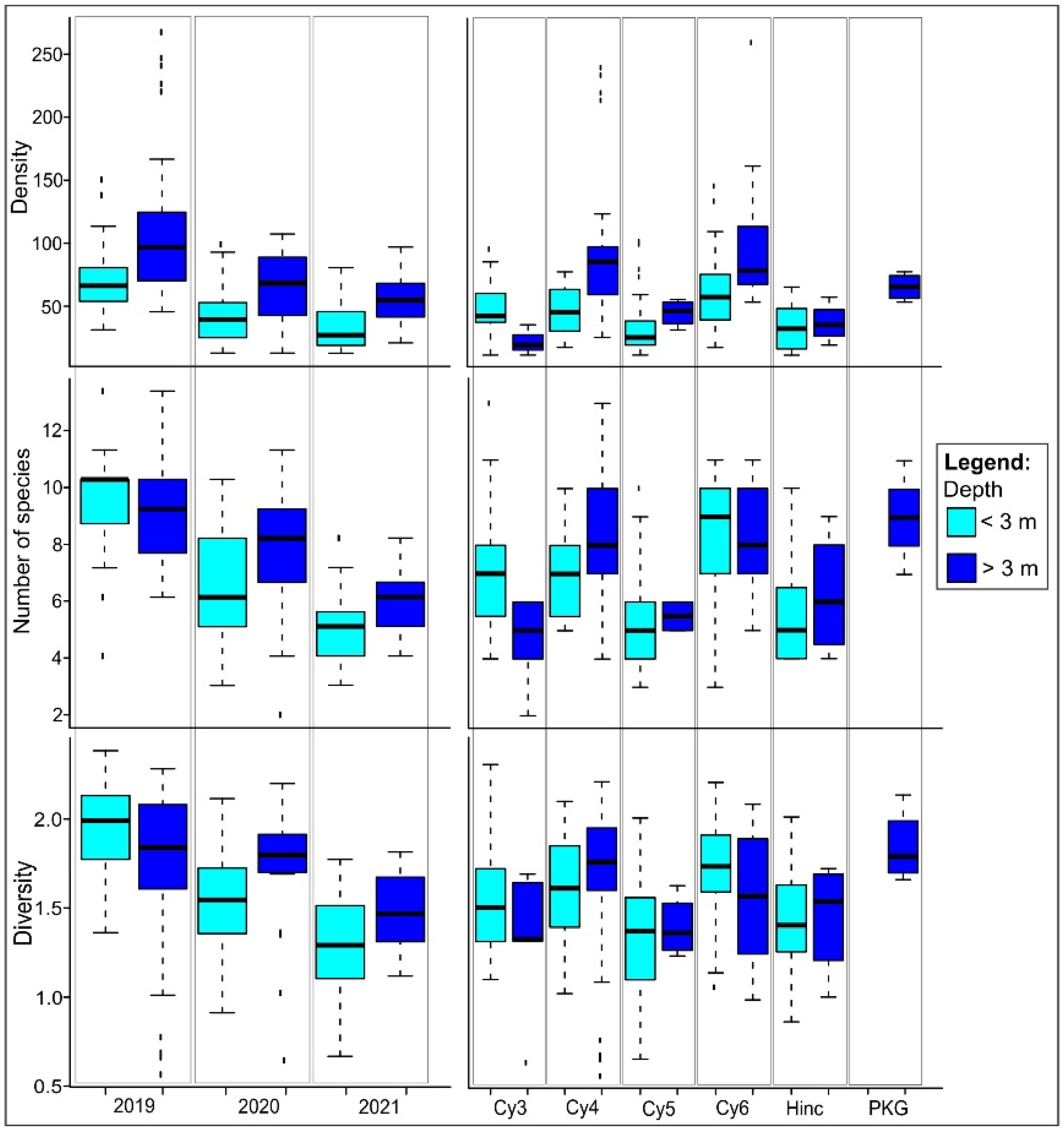
3.2. Coastal Fish Habitat Preference and Trends
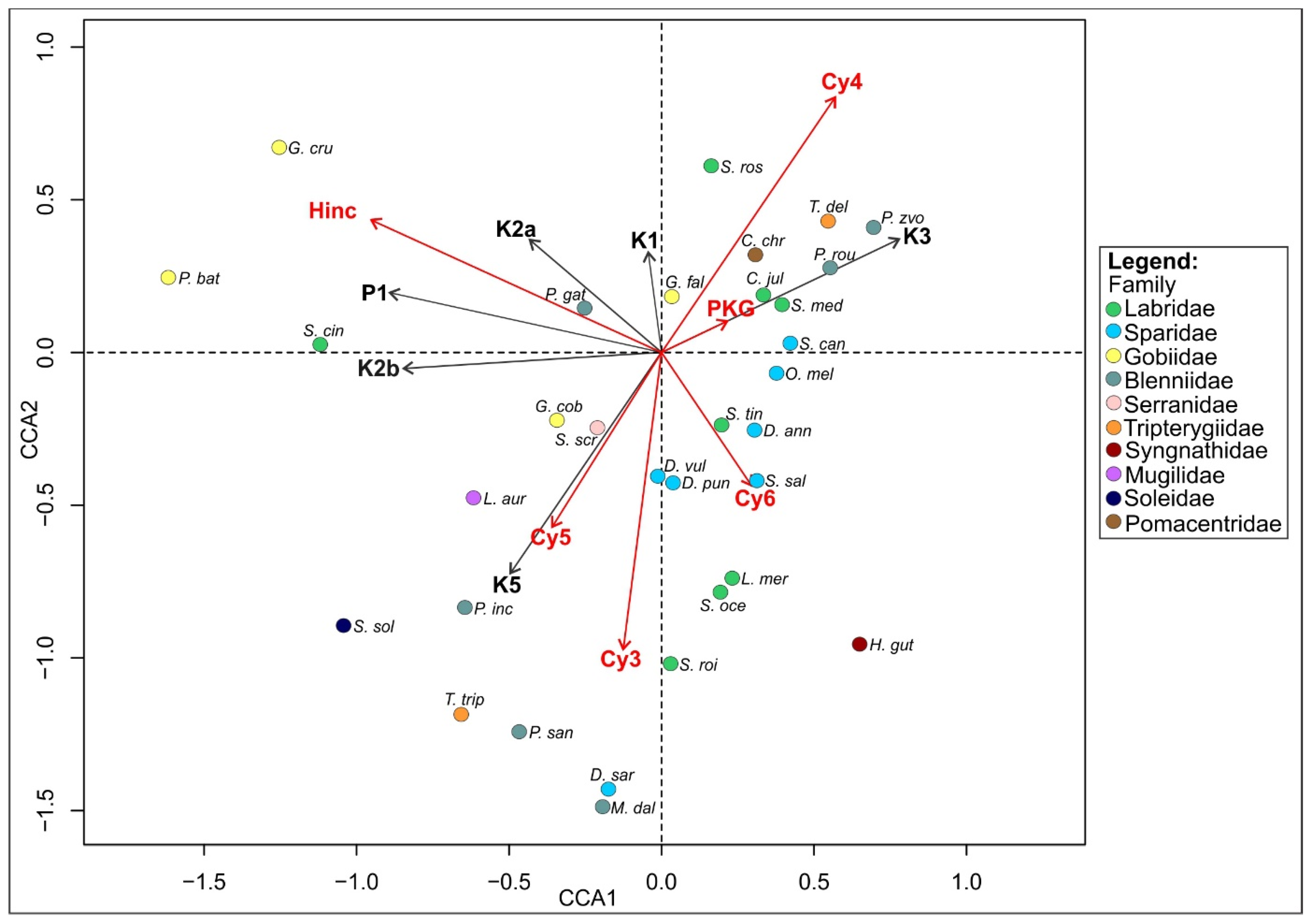
3.3. Coastal Fish Community Differentiating Factors
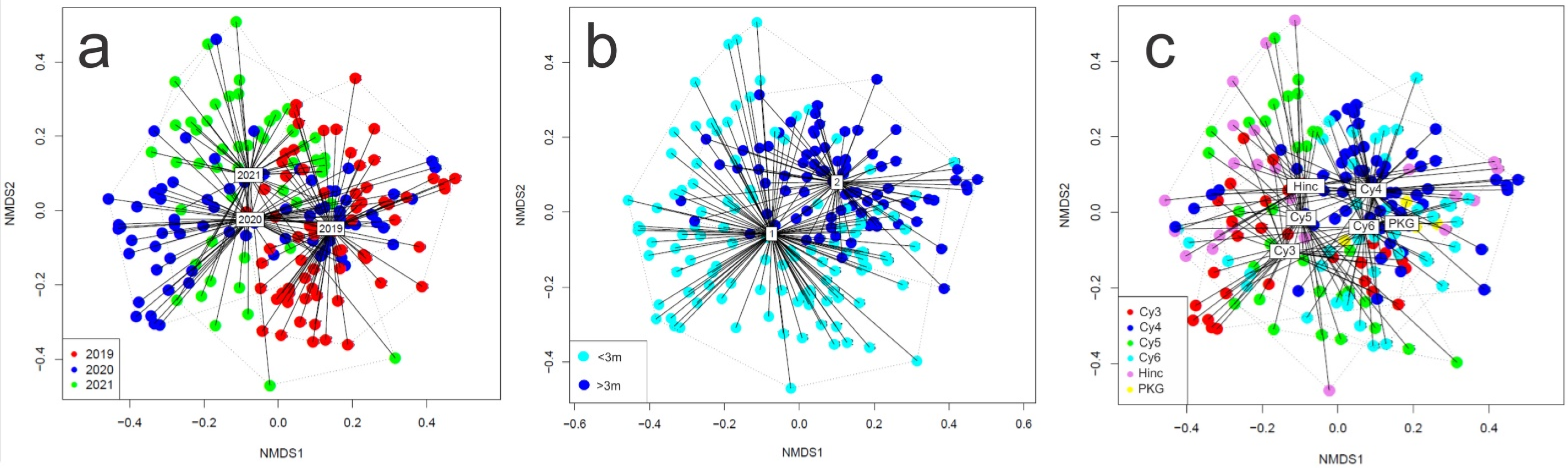
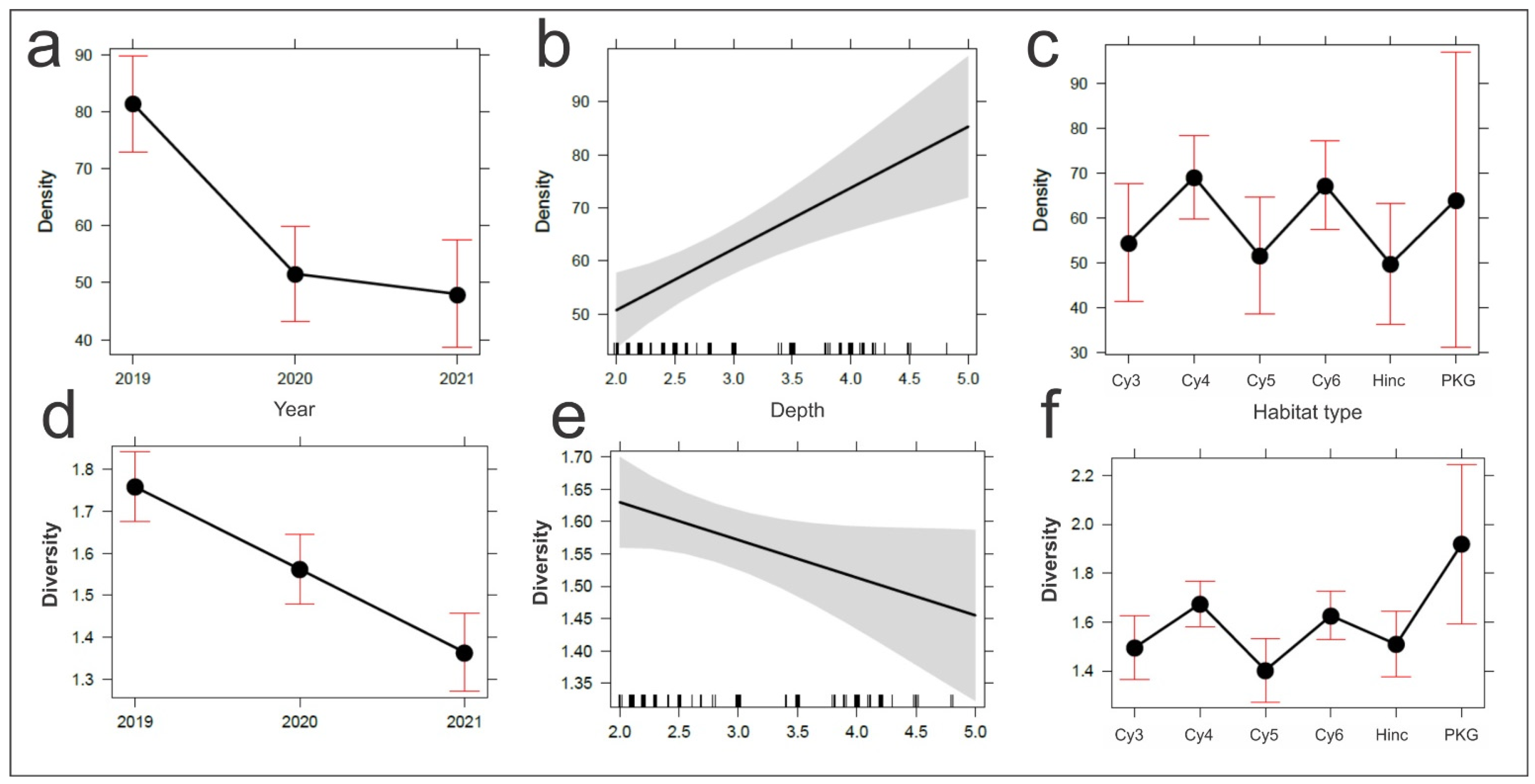
3.4. The Time-Depth-Habitat Type Effect on Coastal Fish Density and Diversity
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Molinari-Novoa, E.A.; Guiry, M.D. Reinstatement of the Genera Gongolaria Boehmer and Ericaria Stackhouse (Sargassaceae, Phaeophyceae). Eur. J. Phycol. 2020, 54, 172. [Google Scholar]
- Vergés, A.; Alcoverro, T.; Ballesteros, E. Role of Fish Herbivory in Structuring the Vertical Distribution of Canopy Algae Cystoseira Spp. in the Mediterranean Sea. Mar. Ecol. Prog. Ser. 2009, 375, 1–11. [Google Scholar] [CrossRef]
- Urra, J.; Rueda, J.L.; Ramírez, Á.M.; Marina, P.; Tirado, C.; Salas, C.; Gofas, S. Seasonal Variation of Molluscan Assemblages in Different Strata of Photophilous Algae in the Alboran Sea (Western Mediterranean). J. Sea Res. 2013, 83, 83–93. [Google Scholar] [CrossRef]
- Pitacco, V.; Orlando-Bonaca, M.; Mavrič, B.; Popović, A.; Lipej, L. Mollusc Fauna Associated with the Cystoseira Algal Associations in the Gulf of Trieste (Northern Adriatic Sea). Medit. Mar. Sci. 2014, 15, 225–238. [Google Scholar] [CrossRef]
- Gozler, A.M.; Kopuz, U.; Agirbas, E. Seasonal Changes of Invertebrate Fauna Associated with Cystoseira Barbata Facies of Southeastern Black Sea Coast. Afr. J. Biotechnol. 2010, 8, 8852–8859. [Google Scholar]
- Orlando-Bonaca, M.; Trkov, D.; Klun, K.; Pitacco, V. Diversity of Molluscan Assemblage in Relation to Biotic and Abiotic Variables in Brown Algal Forests. Plants 2022, 11, 2131. [Google Scholar] [CrossRef]
- Ballesteros, E.; Garrabou, J.; Hereu, B.; Zabala, M.; Cebrian, E.; Sala, E. Deep-Water Stands of Cystoseira Zosteroides C. Agardh (Fucales, Ochrophyta) in the Northwestern Mediterranean: Insights into Assemblage Structure and Population Dynamics. Estuar. Coast. Shelf S 2009, 82, 477–484. [Google Scholar] [CrossRef]
- Mačić, V.; Svirčev, Z. Macroepiphytes on Cystoseira Species (Phaeophyceae) on the Coast of Montenegro. Fresenius Environ. Bull. 2014, 23, 29–34. [Google Scholar] [CrossRef]
- Peleg, O.; Guy-Haim, T.; Yeruham, E.; Silverman, J.; Rilov, G. Tropicalization May Invert Trophic State and Carbon Budget of Shallow Temperate Rocky Reefs. J. Ecol. 2020, 108, 844–854. [Google Scholar] [CrossRef]
- De La Fuente, G.; Fontana, M.; Asnaghi, V.; Chiantore, M.; Mirata, S.; Salis, A.; Damonte, G.; Scarfì, S. The Remarkable Antioxidant and Anti-Inflammatory Potential of the Extracts of the Brown Alga Cystoseira amentacea Var. Stricta. Mar. Drugs 2020, 19, 2. [Google Scholar] [CrossRef] [PubMed]
- Thiriet, P.; Cheminée, A.; Mangialajo, L.; Francour, P.P. How 3D Complexity of Macrophyte-Formed Habitats Affect the Processes Structuring Fish Assemblages Within Coastal Temperate Seascapes? In Underwater Seascapes: From Geographical to Ecological Perspectives; Springer International Publishing: Cham, Germany, 2014; pp. 185–199. ISBN 978-3-319-03439-3. [Google Scholar]
- Hinz, H.; Reñones, O.; Gouraguine, A.; Johnson, A.F.; Moranta, J. Fish Nursery Value of Algae Habitats in Temperate Coastal Reefs. PeerJ 2019, 7, e6797. [Google Scholar] [CrossRef] [PubMed]
- Cheminée, A.; Sala, E.; Pastor, J.; Bodilis, P.; Thiriet, P.; Mangialajo, L.; Cottalorda, J.-M.; Francour, P. Nursery Value of Cystoseira Forests for Mediterranean Rocky Reef Fishes. J. Exp. Mar. Biol. Ecol. 2013, 442, 70–79. [Google Scholar] [CrossRef]
- Cheminée, A.; Pastor, J.; Bianchimani, O.; Thiriet, P.; Sala, E.; Cottalorda, J.-M.; Dominici, J.-M.; Lejeune, P.; Francour, P. Juvenile Fish Assemblages in Temperate Rocky Reefs Are Shaped by the Presence of Macro-Algae Canopy and Its Three-Dimensional Structure. Sci. Rep. 2017, 7, 14638. [Google Scholar] [CrossRef] [PubMed]
- Thibaut, T.; Pinedo, S.; Torras, X.; Ballesteros, E. Long-Term Decline of the Populations of Fucales (Cystoseira Spp. and Sargassum Spp.) in the Albères Coast (France, North-Western Mediterranean). Mar. Pollut. Bull. 2005, 50, 1472–1489. [Google Scholar] [CrossRef] [PubMed]
- Mangialajo, L.; Ruggieri, N.; Asnaghi, V.; Chiantore, M.; Povero, P.; Cattaneo-Vietti, R. Ecological Status in the Ligurian Sea: The Effect of Coastline Urbanisation and the Importance of Proper Reference Sites. Mar. Pollut. Bull. 2007, 55, 30–41. [Google Scholar] [CrossRef]
- Guidetti, P.; Fraschetti, S.; Terlizzi, A.; Boero, F. Effects of Desertification Caused by Lithophaga Lithophaga (Mollusca) Fishery on Littoral Fish Assemblages along Rocky Coasts of Southeastern Italy. Conserv. Biol. 2004, 18, 1417–1423. [Google Scholar] [CrossRef]
- Claudet, J.; Fraschetti, S. Human-Driven Impacts on Marine Habitats: A Regional Meta-Analysis in the Mediterranean Sea. Biol. Conserv. 2010, 143, 2195–2206. [Google Scholar] [CrossRef]
- Blanfuné, A.; Boudouresque, C.-F.; Verlaque, M.; Thibaut, T. The Fate of Cystoseira Crinita, a Forest-Forming Fucale (Phaeophyceae, Stramenopiles), in France (North Western Mediterranean Sea). Estuar. Coast. Shelf S. 2016, 181, 196–208. [Google Scholar] [CrossRef]
- Alberto, F.; Raimondi, P.T.; Reed, D.C.; Coelho, N.C.; Leblois, R.; Whitmer, A.; Serrão, E.A. Habitat Continuity and Geographic Distance Predict Population Genetic Differentiation in Giant Kelp. Ecology 2010, 91, 49–56. [Google Scholar] [CrossRef]
- Savonitto, G.; De La Fuente, G.; Tordoni, E.; Ciriaco, S.; Srijemsi, M.; Bacaro, G.; Chiantore, M.; Falace, A. Addressing Reproductive Stochasticity and Grazing Impacts in the Restoration of a Canopy-forming Brown Alga by Implementing Mitigation Solutions. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 1611–1623. [Google Scholar] [CrossRef]
- Verdura, J.; Santamaría, J.; Ballesteros, E.; Smale, D.A.; Cefalì, M.E.; Golo, R.; Caralt, S.; Vergés, A.; Cebrian, E. Local-scale Climatic Refugia Offer Sanctuary for a Habitat-forming Species during a Marine Heatwave. J. Ecol. 2021, 109, 1758–1773. [Google Scholar] [CrossRef]
- Falace, A.; Alongi, G.; Cormaci, M.; Furnari, G.; Curiel, D.; Cecere, E.; Petrocelli, A. Changes in the Benthic Algae along the Adriatic Sea in the Last Three Decades. Chem. Ecol. 2010, 26, 77–90. [Google Scholar] [CrossRef]
- Orlando-Bonaca, M.; Rotter, A. Any Signs of Replacement of Canopy-Forming Algae by Turf-Forming Algae in the Northern Adriatic Sea? Ecol. Indic. 2018, 87, 272–284. [Google Scholar] [CrossRef]
- Orlando-Bonaca, M.; Pitacco, V.; Lipej, L. Loss of Canopy-Forming Algal Richness and Coverage in the Northern Adriatic Sea. Ecol. Indic. 2021, 125, 107501. [Google Scholar] [CrossRef]
- Cormaci, M.; Furnari, G. Changes of the Benthic Algal Flora of the Tremiti Islands (Southern Adriatic) Italy. Hydrobiologia 2003, 398, 75–79. [Google Scholar]
- Vukovič, A. Associations of Marine Benthic Algae in the Gulf of Piran (In Slovenian). Biol. Vestn. 1980, 28, 103–124. [Google Scholar]
- Giakoumi, S.; Kokkoris, G.D. Effects of Habitat and Substrate Complexity on Shallow Sublittoral Fish Assemblages in the Cyclades Archipelago, North-Eastern Mediterranean Sea. Medit. Mar. Sci. 2013, 14, 58–68. [Google Scholar] [CrossRef]
- Lipej, L.; Bonaca, M.O.; Šiško, M. Coastal Fish Diversity in Three Marine Protected Areas and One Unprotected Area in the Gulf of Trieste (Northern Adriatic). Mar. Ecol. 2003, 24, 259–273. [Google Scholar] [CrossRef]
- Orlando Bonaca, M.; Lipej, L. Factors Affecting Habitat Occupancy of Fish Assemblage in the Gulf of Trieste (Northern Adriatic Sea). Mar. Ecol. 2005, 26, 42–53. [Google Scholar] [CrossRef]
- Battelli, C.; Gregorič, N. First Report of an Aegagropilous Form of Rytiphlaea Tinctoria from the Lagoon of Strunjan (Gulf of Trieste, Northern Adriatic). Annales 2020, 30, 61–68. [Google Scholar]
- ARSO—Slovenian Environment Agency. Available online: https://www.arso.gov.si/en/ (accessed on 10 February 2023).
- Boicourt, W.C.; Kuzmić, M.; Hopkins, T.S. The Inland Sea: Circulation of Chesapeake Bay and the Northern Adriatic. In Ecosystems at the Land-Sea Margin: Drainage Basin to Coastal Sea; Malone, T.C., Malej, A., Harding, L.W., Smodlaka, N., Turner, R.E., Eds.; Coastal and Estuarine Studies; American Geophysical Union: Washington, DC, USA, 1999; Volume 55, pp. 81–129. [Google Scholar]
- Mozetič, P.; Umani, S.F.; Cataletto, B.; Malej, A. Seasonal and Inter-Annual Plankton Variability in the Gulf of Trieste (Northern Adriatic). ICES J. Mar. Sci. 1998, 55, 711–722. [Google Scholar] [CrossRef]
- Stravisi, F. The Vertical Structure Annual Cycle of the Mass Field Parameters in the Gulf of Trieste. Boll. Ocean. Teor. Appl 1983, 1, 239–250. [Google Scholar]
- Ogorelec, B.; Faganeli, J.; Mišič, M.; Čermelj, B. Reconstruction of Paleoenvironment in the Bay of Koper (Gulf of Trieste, Northern Adriatic). Annales 1997, 11, e200. [Google Scholar]
- Harmelin-Vivien, M.; Harmelin, J.; Chauvet, C.; Duval, C.; Galzin, R.; Lejeune, P.; Barnabé, G.; Blanc, F.; Chevalier, R.; Duclerc, J. Evaluation Visuelle Des Peuplements et Populations de Poissons Méthodes et Problèmes. Rev. D’ecologie Terre Vie 1985, 40, 467–539. [Google Scholar] [CrossRef]
- Harmelin-Vivien, M.L.; Francour, P. Trawling or Visual Censuses? Methodological Bias in the Assessment of Fish Populations in Seagrass Beds. Mar. Ecol. 1992, 13, 41–51. [Google Scholar] [CrossRef]
- Patzner, R.; Santos, R. Ecology of Rocky Littoral Fishes of the Azores. Cour. Forschungsinst. Senckenb. 1993, 159, 423–427. [Google Scholar]
- Kulbicki, M.; Cornuet, N.; Vigliola, L.; Wantiez, L.; Moutham, G.; Chabanet, P. Counting Coral Reef Fishes: Interaction between Fish Life-History Traits and Transect Design. J. Exp. Mar. Biol. Ecol. 2010, 387, 15–23. [Google Scholar] [CrossRef]
- Mazzoldi, C.; De Girolamo, M. Littoral Fish Community of the Island Lampedusa (Italy): A Visual Census Approach. Ital. J. Zool. 1998, 65, 275–280. [Google Scholar] [CrossRef]
- Francour, P. The effect of protection level on a coastal fish community at Scandola, Corsica. Rev. D’ecologie Terre Vie 1991, 46, 65–81. [Google Scholar] [CrossRef]
- Harmelin-Vivien, M.; Harmelin, J.; Leboulleux, V. Microhabitat Requirements for Settlement of Juvenile Sparid Fishes on Mediterranean Rocky Shores. In Proceedings of the Space Partition within Aquatic Ecosystems: Proceedings of the Second International Congress of Limnology and Oceanography, Evian, France, 25–28 May 1993; pp. 309–320. [Google Scholar]
- Harmelin, J.G. Structure et Variabilité de I’ichtyofaune d’une Zone Rocheuse Protégée En Méditerranée (Pare National de Port-Cros, France) Structure and Variability of the Ichthyo-fauna in a Mediterranean Protected Rocky Area (National Park of Port-Cros, France). Mar. Ecol. 1987, 8, 263–284. [Google Scholar] [CrossRef]
- Francour, P. Pluriannual Analysis of the Reserve Effect on Ichthyofauna in the Scandola Natural Reserve (Corsica, Northwestern Mediterranean). Oceanol. Acta 1994, 17, 309–3017. [Google Scholar]
- Hilldén, N.-O. Behavioural Ecology of the Labrid Fishes (Teleostei: Labridae) at Tjärnö on the Swedish West Coast; Stockholm University: Stockholm, Sweden, 1984. [Google Scholar]
- Macpherson, E. Substrate Utilisation in a Mediterranean Littoral Fish Community. Mar. Ecol. Prog. Ser. 1994, 114, 211–218. [Google Scholar] [CrossRef]
- Marconato, A.; Mazzoldi, C.; De Girolamo, M.; Stefanni, S.; Maio, G. L’uso Del ‘“Visual Census”’Nello Studio Della Fauna Ittica Costiera. Biol. Mar. Mediterr. 1996, 3, 512–513. [Google Scholar]
- Guidetti, P.; Bussotti, S. Nearshore Fish Assemblages Associated with Shallow Rocky Habitats along the Southern Croatian Coast (Eastern Adriatic Sea). Vie Milieu Life Environ. 2000, 50, 171–176. [Google Scholar]
- Giménez-Casalduero, F.; Gomariz-Castillo, F.J.; Calvín, J.C. Hierarchical Classification of Marine Rocky Landscape as Management Tool at Southeast Mediterranean Coast. Ocean Coast. Manag. 2011, 54, 497–506. [Google Scholar] [CrossRef]
- Thiriet, P. Comparison of Fish Assemblage Structure and Underlying Ecological Processes, between Cystoseira Forests and Less Structurally Complex Habitats of North-Western Mediterranean Rocky Subtidal; Université Nice Sophia Antipolis: Nice, France, 2014. [Google Scholar]
- UNEP. UNEP Draft Classification of Marine Habitat Types for the Mediterranean Region. Mediterranean Action Plan. In Proceedings of the Meeting of Experts on Marine Habitat Types in the Mediterranean Region, Hyères, France, 18 November 1998. SPA/RAC, 149/3: Annex I and II 1998. [Google Scholar]
- Walag, A.M.P.; Canencia, M.O.P. Physico-Chemical Parameters and Macrobenthic Invertebrates of the Intertidal Zone of Gusa, Cagayan de Oro City, Philippines. Adv. Env. Sci. 2016, 8, 71–82. [Google Scholar]
- Huang, J.; Huang, L.; Wu, Z.; Mo, Y.; Zou, Q.; Wu, N.; Chen, Z. Correlation of Fish Assemblages with Habitat and Environmental Variables in a Headwater Stream Section of Lijiang River, China. Sustainability 2019, 11, 1135. [Google Scholar] [CrossRef]
- Ivlev, V. Experimental Ecology of the Feeding of Fishes; Yale University Press: New Haven, CO, USA, 1961. [Google Scholar]
- Wood, K.A.; Stillman, R.A. Do Birds of a Feather Flock Together? Comparing Habitat Preferences of Piscivorous Waterbirds in a Lowland River Catchment. Hydrobiologia 2014, 738, 87–95. [Google Scholar] [CrossRef]
- Kong, D.; Luo, W.; Liu, Q.; Li, Z.; Huan, G.; Zhang, J.; Yang, X. Habitat Use, Preference, and Utilization Distribution of Two Crane Species (Genus: Grus) in Huize National Nature Reserve, Yunnan–Guizhou Plateau, China. PeerJ 2018, 6, e5105. [Google Scholar] [CrossRef]
- Lechowicz, M.J. The Sampling Characteristics of Electivity Indices. Oecologia 1982, 52, 22–30. [Google Scholar] [CrossRef]
- Copp, G.H.; Jurajda, P. Size-Structured Diel Use of River Banks by Fish. Aquat. Sci. 1999, 61, 75–91. [Google Scholar] [CrossRef]
- R Core Team. R Development Core Team R: A Language and Environment for Statistical Computing 2023; R Core Team: Vienna, Austria, 2023. [Google Scholar]
- Legendre, P.; Gallagher, E.D. Ecologically Meaningful Transformations for Ordination of Species Data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. The Vegan Package. Community Ecology Package. Available online: http://R-Forge.r-Project.Org/Projects/Vegan/ (accessed on 1 February 2023).
- Shannon, C.E.; Weaver, W. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity. Curr. Biol. 2021, 31, R1174–R1177. [Google Scholar] [CrossRef]
- Ruitton, S.; Francour, P.; Boudouresque, C.F. Relationships between Algae, Benthic Herbivorous Invertebrates and Fishes in Rocky Sublittoral Communities of a Temperate Sea (Mediterranean). Estuar. Coast. Shelf S 2000, 50, 217–230. [Google Scholar] [CrossRef]
- Sala, E.; Ballesteros, E.; Dendrinos, P.; Di Franco, A.; Ferretti, F.; Foley, D.; Fraschetti, S.; Friedlander, A.; Garrabou, J.; Güçlüsoy, H. The Structure of Mediterranean Rocky Reef Ecosystems across Environmental and Human Gradients, and Conservation Implications. PLoS ONE 2012, 7, e32742. [Google Scholar] [CrossRef]
- Turk, R.; Orlando-Bonaca, M.; Dobrajc, Z.; Lipej, L. Cystoseira Communities in the Slovenian Coast and Their Importance for Fish Fauna. In Proceedings of the third Mediterranean Symposium on Marine Vegetation, Marseilles, France, 27–29 March 2007; UNEP-MAP-RAC-SPA. Regional Activity Centre for Specially Protected Areas: Tunis, Tunisia, 2007; pp. 203–208. [Google Scholar]
- Lipej, B.; Oven, T. Research and Monitoring of Škocjan Bay—Examination of Water Quality Status in the Lagoon of Natural Reserve of Škocjan Bay (In Slovenian); Annual Report; DOPPS BirdLife Slovenia: Ljubljana, Slovenia, 2009; p. 24. [Google Scholar]
- Cuadros, A.; Moranta, J.; Cardona Pascual, L.; Thiret, P.; Francour, P.; Vidal, E.; Sintes, J.; Cheminee, A. Juvenile Fish in Cystoseira Forests: Influence of Habitat Complexity and Depth on Fish Behaviour and Assemblage Composition. Mediterr. Mar. Sci. 2019, 20, 380–392. [Google Scholar] [CrossRef]
- Onofri, I. Prilog Poznavanju Ekologije Porodice Labridae Pelješkog Kanala i Okolnog Podruþja. Master’s Thesis, Univerza u Splitu, Split, Croatia, 1970. [Google Scholar]
- Orlando-Bonaca, M.; Lipej, L.; Orfanidis, S. Benthic Macrophytes as a Tool for Delineating, Monitoring and Assessing Ecological Status: The Case of Slovenian Coastal Waters. Mar. Pollut. Bull. 2008, 56, 666–676. [Google Scholar] [CrossRef]
- Lipej, L.; Orlando-Bonaca, M.; Ozebek, B.; Dulčić, J. Nest Characteristics of Three Labrid Species in the Gulf of Trieste (Northern Adriatic Sea). Acta Adriat. 2009, 50, 139–150. [Google Scholar]
- Gibson, R.N.; Yoshiyama, R.M. Intertidal Fish Communities. In Intertidal Fishes: Life in Two Worlds; Academic Press: San Diego, CA, USA, 1999; pp. 264–296. [Google Scholar]
- Peirano, A.; Niccolai, I.; Mauro, R.; Bianchi, C.N. Seasonal Grazing and Food Preference of Herbivores in a Posidonia Oceanica Meadow. Sci. Mar. 2001, 65, 367–374. [Google Scholar] [CrossRef]
- Dulčić, J.; Lipej, L.; Glamuzina, B.; Bartulović, V. Diet of Spondyliosoma Cantharus and Diplodus Puntazzo (Sparidae) in the Eastern Central Adriatic. Cybium 2006, 30, 115–122. [Google Scholar]
- Gianni, F.; Bartolini, F.; Pey, A.; Laurent, M.; Martins, G.M.; Airoldi, L.; Mangialajo, L. Threats to Large Brown Algal Forests in Temperate Seas: The Overlooked Role of Native Herbivorous Fish. Sci. Rep. 2017, 7, 6012. [Google Scholar] [CrossRef]
- Lokovšek, A.; Orlando-Bonaca, M.; Trkov, D.; Lipej, L. An Insight into the Feeding Ecology of Serranus scriba, a Shallow Water Mesopredator in the Northern Adriatic Sea, with a Non-Destructive Method. Fishes 2022, 7, 210. [Google Scholar] [CrossRef]
- García-Rubies, A.; Macpherson, E. Substrate Use and Temporal Pattern of Recruitment in Juvenile Fishes of the Mediterranean Littoral. Mar. Biol. 1995, 124, 35–42. [Google Scholar] [CrossRef]
- Blanfuné, A.; Boudouresque, C.F.; Verlaque, M.; Thibaut, T. The Ups and Downs of a Canopy-Forming Seaweed over a Span of More than One Century. Sci. Rep. 2019, 9, 5250. [Google Scholar] [CrossRef]
- Orlando-Bonaca, M.; Pitacco, V.; Slavinec, P.; Šiško, M.; Makovec, T.; Falace, A. First Restoration Experiment for Gongolaria Barbata in Slovenian Coastal Waters. What Can Go Wrong? Plants 2021, 10, 239. [Google Scholar] [CrossRef]
- Buñuel, X.; Alcoverro, T.; Pagès, J.F.; Romero, J.; Ruiz, J.M.; Arthur, R. The Dominant Seagrass Herbivore Sarpa Salpa Shifts Its Shoaling and Feeding Strategies as They Grow. Sci. Rep. 2020, 10, 10622. [Google Scholar] [CrossRef]
- Bevilacqua, S.; Savonitto, G.; Lipizer, M.; Mancuso, P.; Ciriaco, S.; Srijemsi, M.; Falace, A. Climatic Anomalies May Create a Long-lasting Ecological Phase Shift by Altering the Reproduction of a Foundation Species. Ecology 2019, 100, 239. [Google Scholar] [CrossRef]
- Iveša, L. Effects of Increased Seawater Temperature and Benthic Mucilage Formation on Shallow Cystoseira Forests of the West Istrian Coast (Northern Adriatic Sea). Seventh Eur. Phycol. Congr. Eur. J. Phycol. 2019, 54, 887–893. [Google Scholar]
- Bulleri, F.; Eriksson, B.K.; Queirós, A.; Airoldi, L.; Arenas, F.; Arvanitidis, C.; Bouma, T.J.; Crowe, T.P.; Davoult, D.; Guizien, K.; et al. Harnessing Positive Species Interactions as a Tool against Climate-Driven Loss of Coastal Biodiversity. PLoS Biol. 2018, 16, e2006852. [Google Scholar] [CrossRef] [PubMed]
- Deza, A.A.; Anderson, T.W. Habitat Fragmentation, Patch Size, and the Recruitment and Abundance of Kelp Forest Fishes. Mar. Ecol. Prog. Ser. 2010, 416, 229–240. [Google Scholar] [CrossRef]
- Perkol-Finkel, S.; Airoldi, L. Loss and Recovery Potential of Marine Habitats: An Experimental Study of Factors Maintaining Resilience in Subtidal Algal Forests at the Adriatic Sea. PLoS ONE 2010, 5, e10791. [Google Scholar] [CrossRef]
- Iveša, L.; Djakovac, T.; Devescovi, M. Long-Term Fluctuations in Cystoseira Populations along the West Istrian Coast (Croatia) Related to Eutrophication Patterns in the Northern Adriatic Sea. Mar. Pollut. Bull. 2016, 106, 162–173. [Google Scholar] [CrossRef]
- Falace, A.; Zanelli, E.; Bressan, G. Algal Transplantation as a Potential Tool for Artificial Reef Management and Environmental Mitigation. Bull. Mar. Sci. 2006, 78, 161–166. [Google Scholar]
- Medrano, A.; Hereu, B.; Cleminson, M.; Pagès-Escolà, M.; Rovira, G.L.; Sola, J.; Linares, C. From Marine Deserts to Algal Beds: Treptacantha Elegans Revegetation to Reverse Stable Degraded Ecosystems inside and Outside a No-Take Marine Reserve. Restor. Ecol. 2020, 28, 632–644. [Google Scholar] [CrossRef]
- Falace, A.; Kaleb, S.; De La Fuente, G.; Asnaghi, V.; Chiantore, M. Ex Situ Cultivation Protocol for Cystoseira amentacea Var. Stricta (Fucales, Phaeophyceae) from a Restoration Perspective. PLoS ONE 2018, 13, e0193011. [Google Scholar] [CrossRef] [PubMed]
- Verdura Brugarola, J.; Sales Villalonga, M.; Ballesteros i Sagarra, E.; Cefalì, M.E.; Cebrian Pujol, E. Restoration of a Canopy-Forming Alga Based on Recruitment Enhancement: Methods and Long-Term Success Assessment. Front. Plant Sci. 2018, 9, 1832. [Google Scholar] [CrossRef]
- Orlando-Bonaca, M.; Savonitto, G.; Asnaghi, V.; Trkov, D.; Pitacco, V.; Šiško, M.; Makovec, T.; Slavinec, P.; Lokovšek, A.; Ciriaco, S. Where and How—New Insight for Brown Algal Forest Restoration in the Adriatic. Front. Mar. Sci. 2022, 9, 988584. [Google Scholar] [CrossRef]
- Gann, G.D.; McDonald, T.; Walder, B.; Aronson, J.; Nelson, C.R.; Jonson, J.; Hallett, J.G.; Eisenberg, C.; Guariguata, M.R.; Liu, J. International Principles and Standards for the Practice of Ecological Restoration. Restor. Ecol. 2019, 27, S1–S46. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat Type-Algal Cover | Code |
|---|---|
| Rocks with dense Cystoseira s.l. | Cy3 |
| Rocks or boulders with dense Padina pavonica | Cy4 |
| Terrace with dense Cystoseira s.l. | Cy5 |
| Boulder with dense Cystoseira s.l. | Cy6 |
| Rocks, terraces, or boulders with Halopithys incurva | Hinc |
| Rocks/boulders with precoralligenous communities | PKG |
| Habitat Type-Substrate Structure | |
| Rough sand <0.5 cm | P1 |
| Bare stones 0.5–10 cm | K1 |
| Bare stones 10–20 cm | K2a |
| Bare stones 20–30 cm | K2b |
| Bare or turf vegetated rocks 30–50 cm | K3 |
| Bare or turf vegetated terrace >50 cm | K5 |
| Bare or turf vegetated boulders >50 cm | K6 |
| Family | Species | 2019 | 2020 | 2021 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FO% | N% | d | Max | FO% | N% | d | Max | FO% | N% | d | Max | ||
| Labridae | Coris julis | 16.40 | 0.66 | 0.60 | 8.00 | 15.50 | 1.32 | 0.68 | 10.00 | 3.90 | 0.28 | 0.62 | 4.00 |
| Labrus merula | 3.00 | 0.07 | 0.06 | 2.00 | 1.40 | 0.06 | 0.03 | 2.00 | 3.90 | 0.19 | 0.08 | 2.00 | |
| Symphodus cinereus | 82.10 | 4.37 | 3.94 | 12.00 | 60.60 | 6.49 | 3.32 | 18.00 | 59.60 | 5.43 | 2.23 | 14.00 | |
| Symphodus doderleini | 1.49 | 0.03 | 0.03 | 2.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Symphodus mediterraneus | 3.00 | 0.10 | 0.09 | 4.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.90 | 0.37 | 0.15 | 8.00 | |
| Symphodus ocellatus | 32.80 | 1.66 | 1.49 | 18.00 | 26.80 | 5.78 | 2.96 | 44.00 | 5.80 | 1.03 | 0.42 | 12.00 | |
| Symphodus roissali | 7.50 | 0.20 | 0.18 | 4.00 | 9.90 | 0.50 | 0.25 | 6.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Symphodus rostratus | 13.40 | 1.13 | 1.01 | 18.00 | 8.50 | 0.33 | 0.17 | 4.00 | 1.90 | 0.09 | 0.04 | 2.00 | |
| Symphodus tinca | 89.60 | 10.69 | 9.64 | 34.00 | 52.10 | 5.39 | 2.76 | 14.00 | 19.20 | 1.78 | 0.73 | 6.00 | |
| Blenniidae | Microlipophrys dalmatinus | 23.90 | 0.89 | 0.81 | 8.00 | 2.80 | 0.11 | 0.06 | 2.00 | 13.50 | 0.65 | 0.27 | 2.00 |
| Parablennius gattorugine | 23.90 | 0.66 | 0.60 | 6.00 | 19.70 | 0.88 | 0.45 | 4.00 | 19.20 | 0.94 | 0.38 | 2.00 | |
| Parablennius incognitus | 13.40 | 0.43 | 0.39 | 6.00 | 2.80 | 0.11 | 0.06 | 2.00 | 3.90 | 0.19 | 0.08 | 2.00 | |
| Parablennius rouxi | 19.40 | 0.63 | 0.57 | 8.00 | 16.90 | 1.10 | 0.56 | 8.00 | 0.00 | 0.00 | 0.08 | 2.00 | |
| Parablennius sanguinolentus | 9.00 | 0.33 | 0.30 | 6.00 | 7.00 | 0.94 | 0.48 | 8.00 | 5.80 | 0.37 | 0.15 | 4.00 | |
| Parablennius tentacularis | 0.00 | 0.00 | 0.00 | 0.00 | 7.00 | 0.44 | 0.23 | 4.00 | 1.90 | 0.09 | 0.04 | 2.00 | |
| Parablennius zvonimiri | 11.50 | 0.03 | 0.03 | 2.00 | 2.80 | 0.44 | 0.23 | 8.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Sparidae | Diplodus annularis | 26.90 | 0.93 | 0.84 | 6.00 | 9.90 | 0.61 | 0.31 | 6.00 | 3.90 | 0.37 | 0.15 | 6.00 |
| Diplodus vulgaris | 98.50 | 13.37 | 12.06 | 42.00 | 71.80 | 9.47 | 4.85 | 26.00 | 50.00 | 8.79 | 3.62 | 32.00 | |
| Diplodus puntazzo | 32.80 | 1.06 | 0.96 | 8.00 | 23.90 | 1.38 | 0.70 | 8.00 | 11.50 | 1.40 | 0.58 | 10.00 | |
| Diplodus sargus | 0.00 | 0.00 | 0.00 | 0.00 | 1.40 | 0.11 | 0.06 | 4.00 | 3.90 | 0.19 | 0.08 | 2.00 | |
| Oblada melanura | 4.50 | 0.13 | 0.12 | 4.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Sarpa salpa | 20.90 | 3.67 | 3.31 | 54.00 | 8.50 | 1.10 | 0.56 | 14.00 | 1.90 | 2.06 | 0.85 | 44.00 | |
| Spondylliosoma cantharus | 10.50 | 0.46 | 0.42 | 8.00 | 1.40 | 0.06 | 0.03 | 2.00 | 1.90 | 0.09 | 0.04 | 2.00 | |
| Gobiidae | Gobius cobitis | 16.40 | 0.40 | 0.36 | 4.00 | 9.90 | 0.39 | 0.20 | 2.00 | 13.50 | 1.12 | 0.46 | 10.00 |
| Gobius cruentatus | 23.90 | 0.53 | 0.48 | 2.00 | 56.30 | 8.31 | 4.25 | 20.00 | 21.20 | 2.34 | 0.96 | 10.00 | |
| Gobius fallax | 92.50 | 12.45 | 11.22 | 42.00 | 62.00 | 7.93 | 4.06 | 24.00 | 88.50 | 31.90 | 13.12 | 52.00 | |
| Pomatoschistus bathi | 14.90 | 1.62 | 1.46 | 14.00 | 21.10 | 2.15 | 1.10 | 10.00 | 26.90 | 3.55 | 1.46 | 12.00 | |
| Serranidae | Serranus hepatus | 0.00 | 0.00 | 0.00 | 0.00 | 16.90 | 0.83 | 0.42 | 6.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Serranus scriba | 95.50 | 9.76 | 8.81 | 24.00 | 97.20 | 20.91 | 10.70 | 46.00 | 94.20 | 19.64 | 8.08 | 22.00 | |
| Tripterygiidae | Tripterygion delaisi | 37.30 | 1.66 | 1.49 | 10.00 | 9.90 | 0.50 | 0.25 | 4.00 | 15.40 | 0.84 | 0.35 | 4.00 |
| Tripterygion tripteronotus | 11.90 | 0.46 | 0.42 | 8.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Syngnathidae | Hippocampus guttulatus | 1.50 | 0.03 | 0.03 | 2.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Pomacentridae | Chromis chromis | 77.60 | 31.58 | 28.48 | 200.00 | 50.70 | 21.63 | 11.07 | 44.00 | 50.00 | 16.18 | 6.65 | 36.00 |
| Soleidae | Solea solea | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.90 | 0.00 | 0.04 | 2.00 |
| Mugilidae | Liza aurata | 0.00 | 0.00 | 0.00 | 0.00 | 4.20 | 0.77 | 0.39 | 16.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Cy3 | Cy4 | Cy5 | Cy6 | Hinc | |
|---|---|---|---|---|---|
| Cy4 | C. chr (>Cy4) | ||||
| S.scr (>Cy3) | |||||
| G. fal (>Cy4) | |||||
| 0.45 | |||||
| Cy5 | G. fal (>Cy5) | C. chr (>Cy4) | |||
| S. scr (>Cy3) | G. fal (>Cy5) | ||||
| D. vul (>Cy5) | S. scr (>Cy5) | ||||
| 0.40 | 0.44 | ||||
| Cy6 | C. chr (>Cy6) | C. chr (>Cy4) | G. fal (>Cy5) | ||
| S. scr (>Cy3) | G. fal (>Cy4) | C. chr (>Cy6) | |||
| G. fal (>Cy6) | D. vul (>Cy6) | D. vul (>Cy5) | |||
| 0.41 | 0.45 | 0.41 | |||
| Hinc | S. scr (>Cy3) | C. chr (>Cy4) | G. fal (>Cy5) | C. chr (>Cy6) | |
| S.cin (>Hinc) | G. fal (>Cy4) | D. vul (>Cy5) | G. fal (>Cy6) | ||
| C. chr (>Hinc) | S. cin (>Hinc) | S. cin (>Hinc) | S.cin (>Hinc) | ||
| 0.35 | 0.41 | 0.38 | 0.38 | ||
| PKG | C. chr (>PKG) | C. chr (>PKG) | C. chr (>Cy5) | C. chr (>PKG) | C. chr (>PKG) |
| S. scr (>Cy3) | G. fal (>Cy4) | G. fal (>Cy5) | D. vul (>Cy6) | S. cin (>Hinc) | |
| S. oce (>Cy3) | S. scr (>PKG) | S. tin (>PKG) | G. fal (>Cy6) | G. cru (>Hinc) | |
| 0.44 | 0.45 | 0.46 | 0.42 | 0.39 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lipej, L.; Ivajnšič, D.; Pitacco, V.; Trkov, D.; Mavrič, B.; Orlando-Bonaca, M. Coastal Fish Fauna in the Cystoseira s.l. Algal Belts: Experiences from the Northern Adriatic Sea. J. Mar. Sci. Eng. 2023, 11, 888. https://doi.org/10.3390/jmse11050888
Lipej L, Ivajnšič D, Pitacco V, Trkov D, Mavrič B, Orlando-Bonaca M. Coastal Fish Fauna in the Cystoseira s.l. Algal Belts: Experiences from the Northern Adriatic Sea. Journal of Marine Science and Engineering. 2023; 11(5):888. https://doi.org/10.3390/jmse11050888
Chicago/Turabian StyleLipej, Lovrenc, Danijel Ivajnšič, Valentina Pitacco, Domen Trkov, Borut Mavrič, and Martina Orlando-Bonaca. 2023. "Coastal Fish Fauna in the Cystoseira s.l. Algal Belts: Experiences from the Northern Adriatic Sea" Journal of Marine Science and Engineering 11, no. 5: 888. https://doi.org/10.3390/jmse11050888